
Transcriptome Analysis Uncovers the Gene Expression Profile of Hemileia vastatrix (Race XXXIII) during the Interactions with Resistant and Susceptible Coffee

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Cytological Evaluation of the Infection Process and Plant Responses
2.2. Sequencing of H. vastatrix and Coffee-Fungus Interaction Libraries
2.3. Read Processing, Mapping, Transcriptome Assembly, Expression Quantification, and Identification of Differentially Expressed Genes (DEGs)
2.4. Co-Expression Analysis
2.5. Functional Annotation
2.6. Differentially Expressed Gene Validation by Real-Time qPCR
3. Results
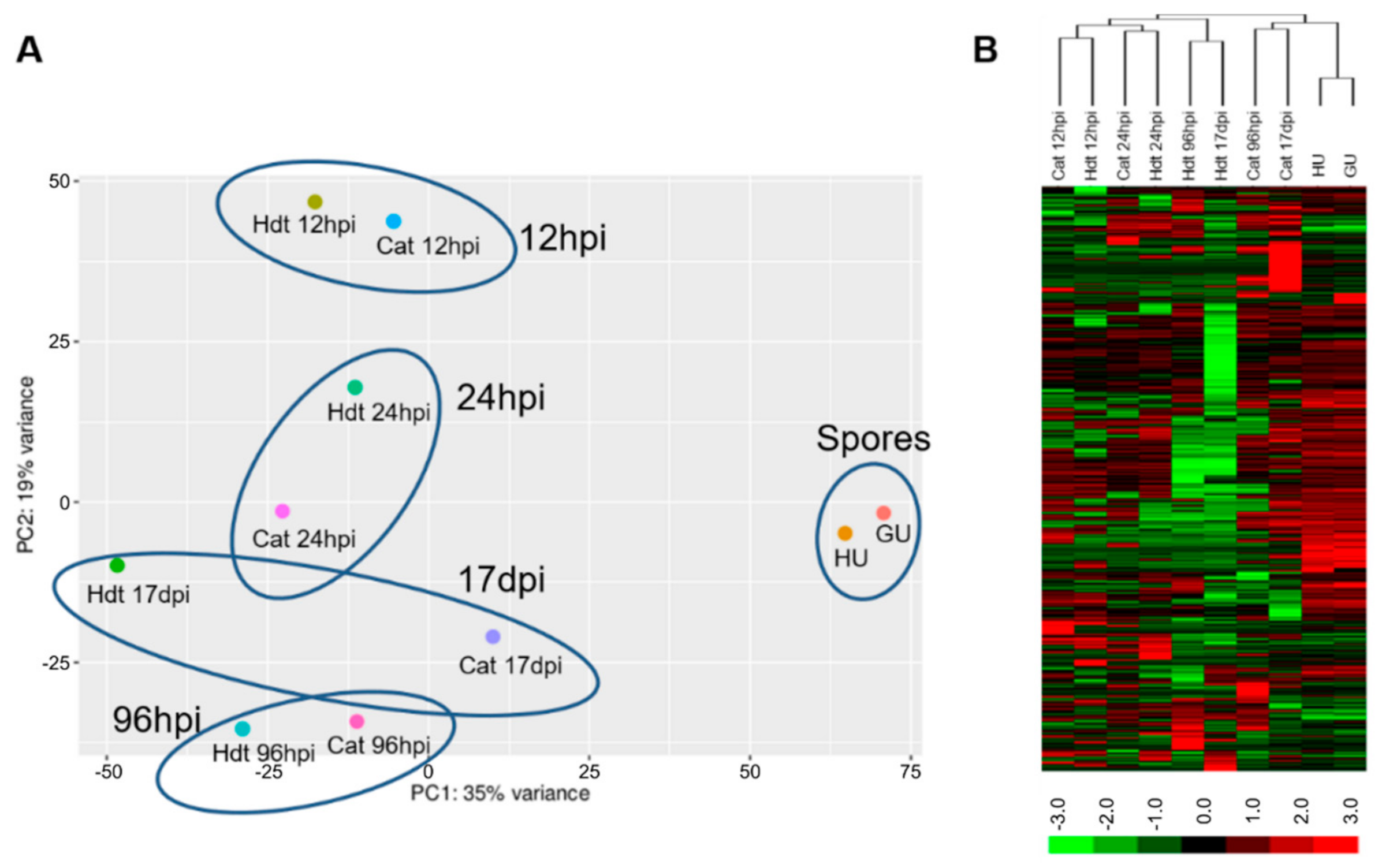
3.1. Transcriptome of H. vastatrix
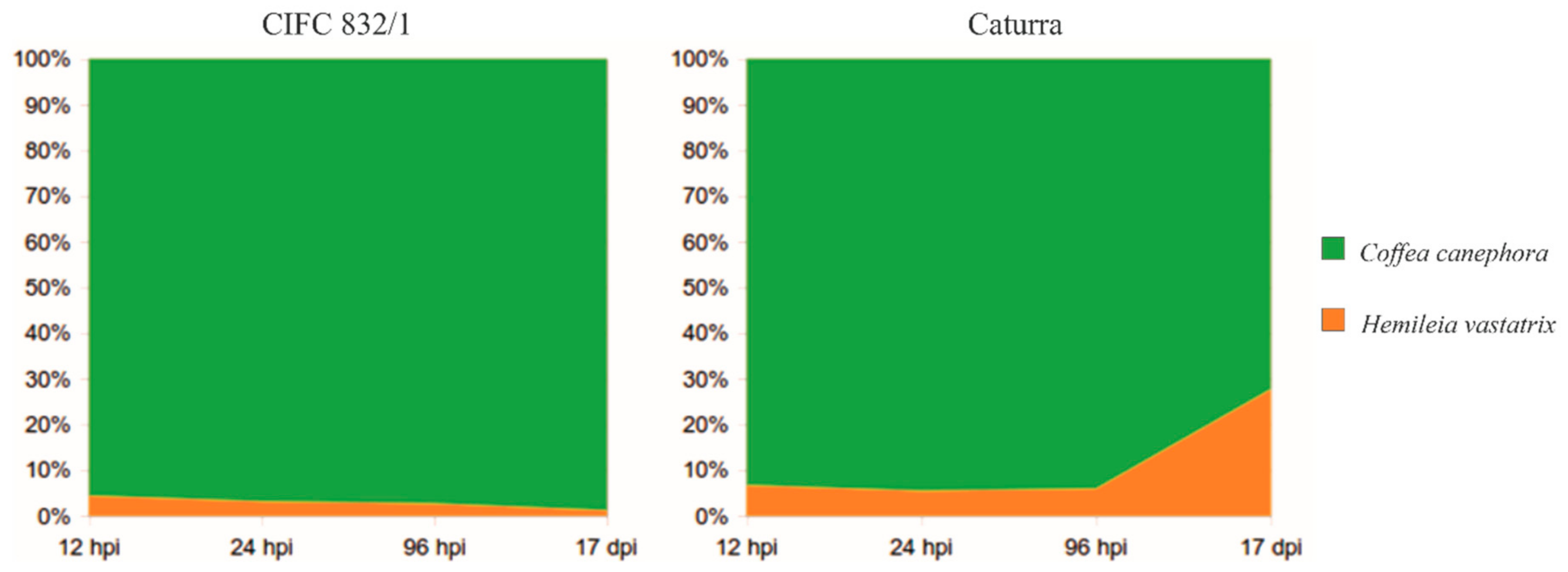
3.2. Changes in the Fungal Transcriptome When Interacting with the Plant
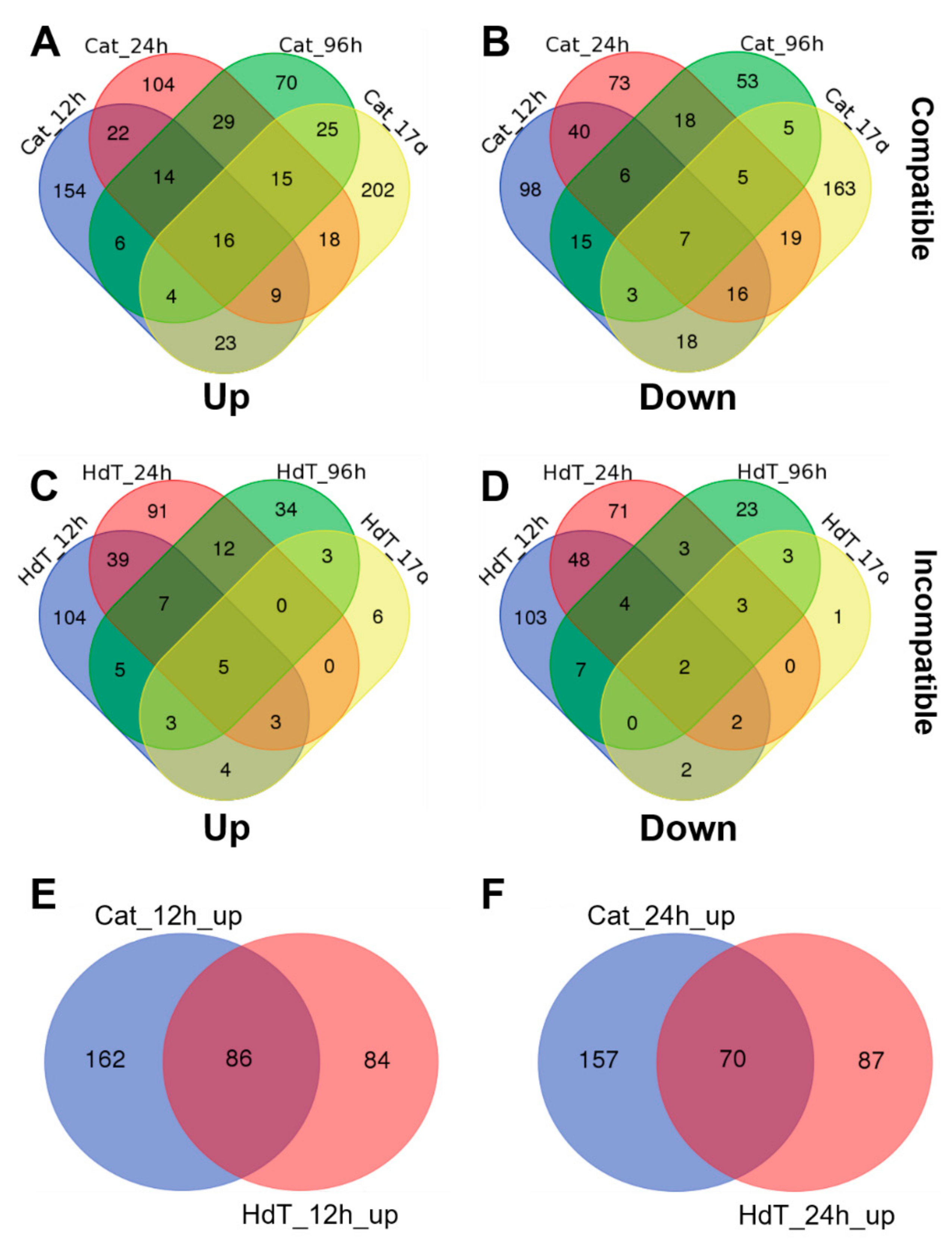
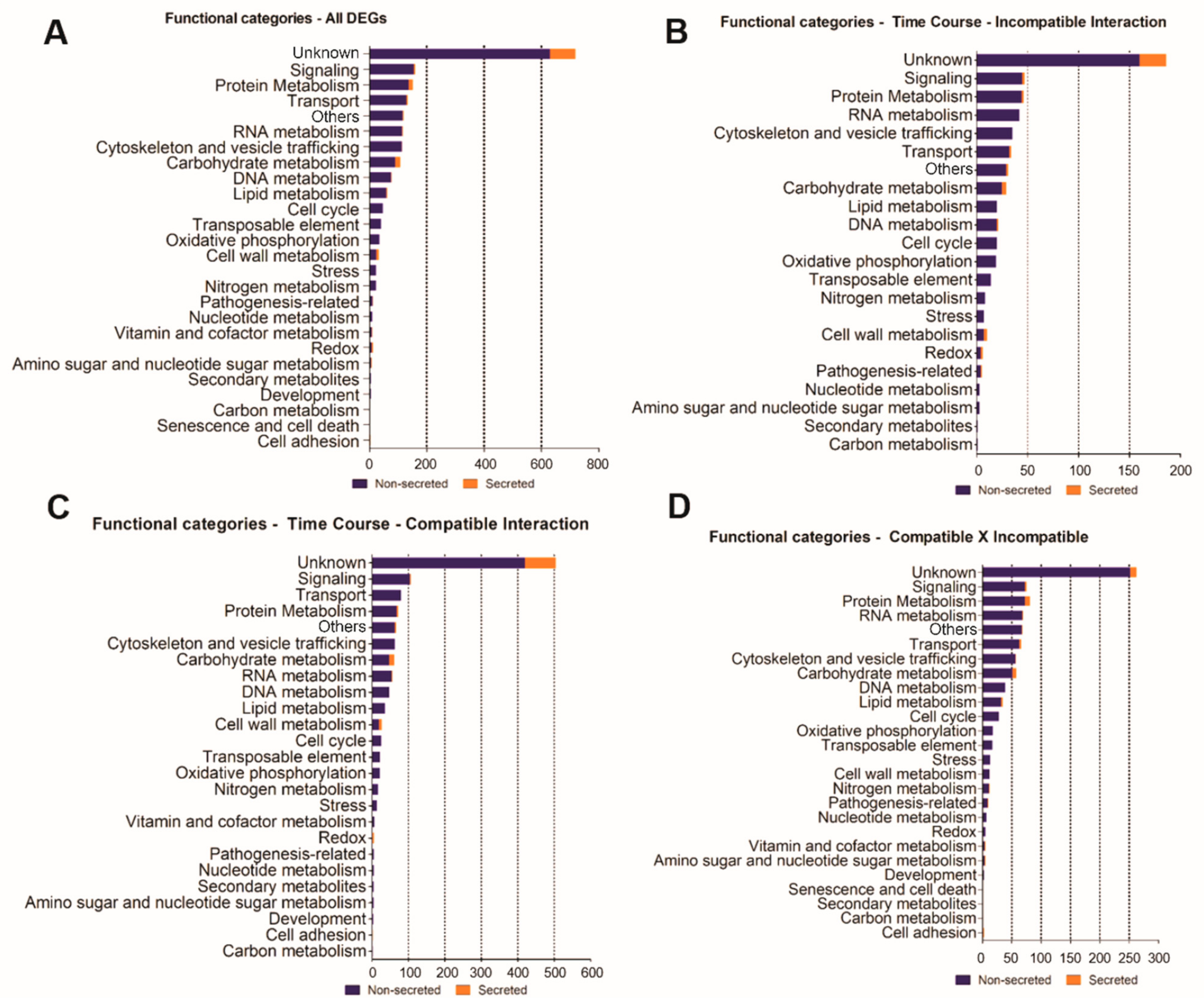
3.3. Differentially Expressed Genes (DEGs)
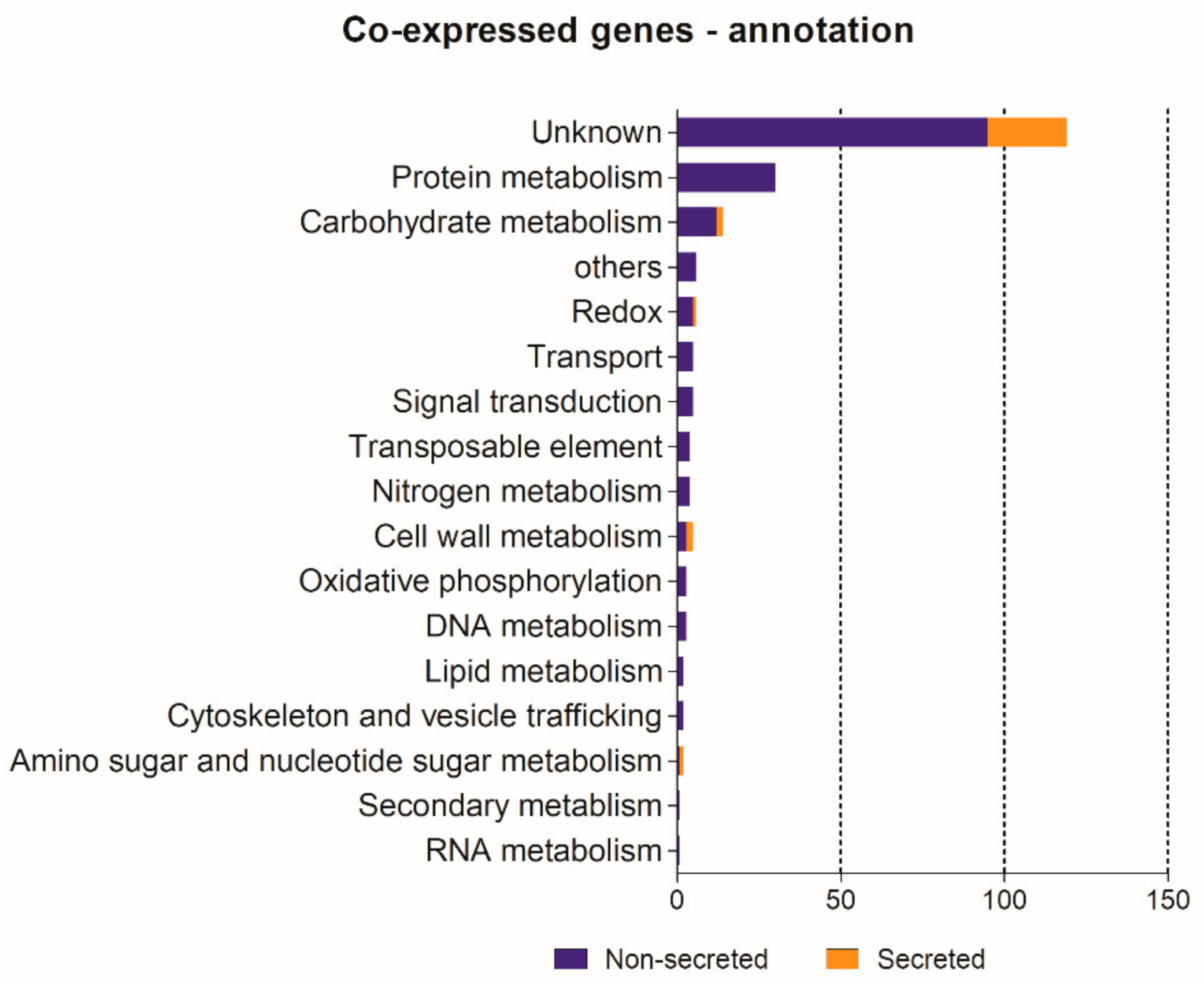
3.4. Identification of Putative Effector Homologues from Other Races and Co-Expression Network Analysis
4. Expression Analysis by qPCR
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zambolim, L.; Caixeta, E.T. An overview of physiological specialization of coffee leaf rust-new designation of pathotypes. Int. J. Curr. Res. 2021, 13, 15564–15575. [Google Scholar] [CrossRef]
- Zambolim, L. Current status and management of coffee leaf rust in Brazil. Trop. Plant Pathol. 2016, 41, 1–8. [Google Scholar] [CrossRef]
- Avelino, J.; Cristancho, M.; Georgiou, S.; Imbach, P.; Aguilar, L.; Bornemann, G.; Läderach, P.; Anzueto, F.; Hruska, A.J.; Morales, C. The coffee rust crises in Colombia and Central America (2008–2013): Impacts, plausible causes and proposed solutions. Food Secur. 2015, 7, 303–321. [Google Scholar] [CrossRef]
- Pereira, D.R.; Nadaleti, D.H.S.; Rodrigues, E.C.; da Silva, A.D.; Malta, M.R.; de Carvalho, S.P.; Carvalho, G.R. Genetic and chemical control of coffee rust (Hemileia vastatrix Berk et Br.): Impacts on coffee (Coffea arabica L.) quality. J. Sci. Food Agric. 2021, 101, 2836–2845. [Google Scholar] [CrossRef] [PubMed]
- Rozo, Y.; Escobar, C.; Gaitán, Á.; Cristancho, M. Aggressiveness and Genetic Diversity of Hemileia vastatrix During an Epidemic in Colombia. J. Phytopathol. 2012, 160, 732–740. [Google Scholar] [CrossRef]
- Keith, L.M.; Sugiyama, L.S.; Brill, E.; Adams, B.L.; Fukada, M.; Hoffman, K.M.; Ocenar, J.; Kawabata, A.; Kong, A.T.; McKemy, J.M.; et al. First Report of Coffee Leaf Rust Caused by Hemileia vastatrix on Coffee (Coffea arabica) in Hawaii. Plant Dis. 2022. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Camejo, L.A.; Eamvijarn, A.; Díaz-Valderrama, J.R.; Karlsen-Ayala, E.; Koch, R.; Johnson, E.; Pruvot-Woehl, S.; Mejía, L.C.; Montagnon, C.; Maldonado-Fuentes, C.; et al. Global analysis of Hemileia vastatrix populations shows clonal reproduction for the coffee leaf rust pathogen throughout most of its range. Phytopathology 2021. [Google Scholar] [CrossRef]
- Várzea, V.M.P.; Marques, D. Durable Resistance to Coffee Leaf Rust. In Durable Resistance to Coffee Leaf Rust; Zambolim, L., Zambolim, E.M., Eds.; Universidade Federal de Viçosa: Viçosa, Brasil, 2005; pp. 53–74. [Google Scholar]
- Silva, D.N.; Várzea, V.; Paulo, O.S.; Batista, D. Population genomic footprints of host adaptation, introgression and recombination in coffee leaf rust. Mol. Plant Pathol. 2018, 19, 1742–1753. [Google Scholar] [CrossRef]
- Talhinhas, P.; Batista, D.; Diniz, I.; Vieira, A.; Silva, D.N.; Loureiro, A.; Tavares, S.; Pereira, A.P.; Azinheira, H.G.; Guerra-Guimarães, L.; et al. The coffee leaf rust pathogen Hemileia vastatrix: One and a half centuries around the tropics. Mol. Plant Pathol. 2017, 18, 1039–1051. [Google Scholar] [CrossRef]
- Cabral, P.G.C.; Zambolim, E.M.; Zambolim, L.; Lelis, T.P.; Capucho, A.S.; Caixeta, E.T. Identification of a new race of Hemileia vastatrix in Brazil. Australas. Plant Dis. Notes 2009, 4, 129–130. [Google Scholar] [CrossRef]
- Capucho, A.S.; Zambolim, E.M.; Freitas, R.L.; Haddad, F.; Caixeta, E.T.; Zambolim, L. Identification of race XXXIII of Hemileia vastatrix on Coffea arabica Catimor derivatives in Brazil. Australas. Plant Dis. Notes 2012, 7, 189–191. [Google Scholar] [CrossRef]
- Setotaw, T.A.; Caixeta, E.T.; Zambolim, E.M.; Sousa, T.V.; Pereira, A.A.; Baião, A.C.; Cruz, C.D.; Zambolim, L.; Sakiyama, N.S. Genome Introgression of Híbrido de Timor and Its Potential to Develop High Cup Quality C. arabica Cultivars. J. Agric. Sci. 2020, 12, 64. [Google Scholar] [CrossRef]
- Schulze-Lerfert, P.; Panstruga, R. Establishment of biotrophy by parasitic fungi and reprogramming of host cells for disease resistance. Annu. Rev. Phytopathol. 2003, 41, 641–667. [Google Scholar] [CrossRef]
- Voegele, R.T.; Mendgen, K. Rust haustoria: Nutrient uptake and beyond. New Phytol. 2003, 159, 93–100. [Google Scholar] [CrossRef]
- Voegele, R.T. Uromyces fabae: Development, metabolism, and interactions with its host Vicia faba. FEMS Microbiol. Lett. 2006, 259, 165–173. [Google Scholar] [CrossRef][Green Version]
- Xu, Q.; Tang, C.; Wang, L.; Zhao, C.; Kang, Z.; Wang, X. Haustoria–arsenals during the interaction between wheat and Puccinia striiformis f. sp. tritici. Mol. Plant Pathol. 2020, 21, 83–94. [Google Scholar] [CrossRef]
- Polonio, Á.; Pérez-García, A.; Martínez-Cruz, J.; Fernández-Ortuño, D.; de Vicente, A. The Haustorium of Phytopathogenic Fungi: A Short Overview of a Specialized Cell of Obligate Biotrophic Plant Parasites. In Progress in Botany; Cánovas, F.M., Lüttge, U., Risueño, M.C., Leuschner, C., Eds.; Springer: Cham, Switzerland, 2020; pp. 337–355. [Google Scholar]
- Mendgen, K.; Hahn, M. Plant infection and the establishment of fungal biotrophy. Trends Plant Sci. 2002, 7, 352–356. [Google Scholar] [CrossRef]
- Dodds, P.N.; Rafiqi, M.; Gan, P.H.P.; Hardham, A.R.; Jones, D.A.; Ellis, J.G. Effectors of biotrophic fungi and oomycetes: Pathogenicity factors and triggers of host resistance. New Phytol. 2009, 183, 993–1000. [Google Scholar] [CrossRef]
- Figueroa, M.; Ortiz, D.; Henningsen, E.C. Tactics of host manipulation by intracellular effectors from plant pathogenic fungi. Curr. Opin. Plant Biol. 2021, 62, 102054. [Google Scholar] [CrossRef]
- Flor, H.H. Current status of the gene-fob-gene concept. Annu. Rev. Phytopathol. 1971, 9, 275–296. [Google Scholar] [CrossRef]
- Rodrigues, C.J.; Bettencourt, A.J.; Rijo, L. Races of the Pathogen and Resistance to Coffee Rust. Annu. Rev. Phytopathol. 1975, 13, 49–70. [Google Scholar] [CrossRef]
- Hae Choi, Y.; Verpoorte, R. Metabolomics: What You See is What You Extract. Phytochem. Anal. 2014, 25, 289–290. [Google Scholar] [CrossRef]
- De Almeida, D.P.; Castro, I.S.L.; de Oliveira Mendes, T.A.; Alves, D.R.; Barka, G.D.; Barreiros, P.R.R.M.; Zambolim, L.; Sakiyama, N.S.; Caixeta, E.T. Receptor-Like Kinase (RLK) as a candidate gene conferring resistance to Hemileia vastatrix in coffee. Sci. Agric. 2020, 78, 1–9. [Google Scholar] [CrossRef]
- Barka, G.D.; Caixeta, E.T.; Ferreira, S.S.; Zambolim, L. In silico guided structural and functional analysis of genes with potential involvement in resistance to coffee leaf rust: A functional marker based approach. PLoS ONE 2020, 15, e0222747. [Google Scholar] [CrossRef] [PubMed]
- Florez, J.C.; Mofatto, L.S.; do Livramento Freitas-Lopes, R.; Ferreira, S.S.; Zambolim, E.M.; Carazzolle, M.F.; Zambolim, L.; Caixeta, E.T. High throughput transcriptome analysis of coffee reveals prehaustorial resistance in response to Hemileia vastatrix infection. Plant Mol. Biol. 2017, 95, 607–623. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, D.; Tisserant, E.; Talhinhas, P.; Azinheira, H.; Vieira, A.; Petitot, A.-S.; Loureiro, A.; Poulain, J.; DA Silva, C.; Silva, M.D.C.; et al. 454-pyrosequencing of Coffea arabica leaves infected by the rust fungus Hemileia vastatrix reveals in planta-expressed pathogen-secreted proteins and plant functions in a late compatible plant-rust interaction. Mol. Plant Pathol. 2012, 13, 17–37. [Google Scholar] [CrossRef] [PubMed]
- Talhinhas, P.; Azinheira, H.G.; Vieira, B.; Loureiro, A.; Tavares, S.; Batista, D.; Morin, E.; Petitot, A.-S.; Paulo, O.S.; Poulain, J.; et al. Overview of the functional virulent genome of the coffee leaf rust pathogen Hemileia vastatrix with an emphasis on early stages of infection. Front. Plant Sci. 2014, 5, 88. [Google Scholar] [CrossRef] [PubMed]
- Cristancho, M.A.; Botero-Rozo, D.O.; Giraldo, W.; Tabima, J.; Riaño-Pachón, D.M.; Escobar, C.; Rozo, Y.; Rivera, L.F.; Durán, A.; Restrepo, S.; et al. Annotation of a hybrid partial genome of the coffee rust (Hemileia vastatrix) contributes to the gene repertoire catalog of the Pucciniales. Front. Plant Sci. 2014, 5, 1–11. [Google Scholar] [CrossRef]
- Porto, B.N.; Caixeta, E.T.; Mathioni, S.M.; Vidigal, P.M.P.; Zambolim, L.; Zambolim, E.M.; Donofrio, N.; Polson, S.W.; Maia, T.A.; Chen, C.; et al. Genome sequencing and transcript analysis of Hemileia vastatrix reveal expression dynamics of candidate effectors dependent on host compatibility. PLoS ONE 2019, 14, e0215598. [Google Scholar] [CrossRef]
- Freitas, R.d.L.; Maciel-Zambolim, E.; Silva, M.d.C.; Caixeta, E.T.; Lelis, D.T.; Zambolim, L.; Sakiyama, N.S. Cytological Evaluation of the Infection Process of Hemileia vastatrix (race XXXIII) in Resistant and Susceptible Coffee. In Proceedings of the International Conference on Coffee Science (ASIC), Armenia, Colombia, 8–13 September 2014; p. 226. [Google Scholar]
- Silva, M.C.; Nicole, M.; Rijo, L.; Geiger, J.P.; Rodrigues, C.J., Jr. Cytochemical Aspects of the Plant–Rust Fungus Interface during the Compatible Interaction Coffea arabica (cv. Caturra)–Hemileia vastatrix (race III). Int. J. Plant Sci. 1999, 160, 79–91. [Google Scholar] [CrossRef]
- Silva, M.C.; Nicole, M.; Guerra-Guimarães, L.; Rodrigues, C.J. Hypersensitive cell death and post-haustorial defence responses arrest the orange rust (Hemileia vastatrix) growth in resistant coffee leaves. Physiol. Mol. Plant Pathol. 2002, 60, 169–183. [Google Scholar] [CrossRef]
- Silva, M.C.; Guerra-Guimarães, L.; Loureiro, A.; Nicole, M.R. Involvement of peroxidases in the coffee resistance to orange rust (Hemileia vastatrix). Physiol. Mol. Plant Pathol. 2008, 72, 29–38. [Google Scholar] [CrossRef]
- Diniz, I.; Talhinhas, P.; Azinheira, H.G.; Várzea, V.; Medeira, C.; Maia, I.; Petitot, A.S.; Nicole, M.; Fernandez, D.; do Céu Silva, M. Cellular and molecular analyses of coffee resistance to Hemileia vastatrix and nonhost resistance to Uromyces vignae in the resistance-donor genotype HDT832/2. Eur. J. Plant Pathol. 2012, 133, 141–157. [Google Scholar] [CrossRef]
- Bennett, M.; Gallagher, M.; Fagg, J.; Bestwick, C.; Paul, T.; Beale, M.; Mansfield, J. The hypersensitive reaction, membrane damage and accumulation of autofluorescent phenolics in lettuce cells challenged by Bremia lactucae. Plant J. 1996, 9, 851–865. [Google Scholar] [CrossRef]
- Heath, M.C. Involvement of reactive oxygen species in the response of resistant (hypersensitive) or susceptible cowpeas to the cowpea rust fungus. New Phytol. 1998, 138, 251–263. [Google Scholar] [CrossRef]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef]
- Maia, T.; Badel, J.L.; Marin-Ramirez, G.; Rocha, C.d.M.; Fernandes, M.B.; da Silva, J.C.F.; de Azevedo-Junior, G.M.; Brommonschenkel, S.H. The Hemileia vastatrix effector HvEC-016 suppresses bacterial blight symptoms in coffee genotypes with the SH 1 rust resistance gene. New Phytol. 2017, 213, 1315–1329. [Google Scholar] [CrossRef] [PubMed]
- Saito, R.; Smoot, M.E.; Ono, K.; Ruscheinski, J.; Wang, P.-L.; Lotia, S.; Pico, A.R.; Bader, G.D.; Ideker, T. A travel guide to Cytoscape plugins. Nat. Methods 2012, 9, 1069–1076. [Google Scholar] [CrossRef] [PubMed]
- Vieira, A.; Talhinhas, P.; Loureiro, A.; Duplessis, S.; Fernandez, D.; Silva, M.d.C.; Paulo, O.S.; Azinheira, H.G. Validation of RT-qPCR reference genes for in planta expression studies in Hemileia vastatrix, the causal agent of coffee leaf rust. Fungal Biol. 2011, 115, 891–901. [Google Scholar] [CrossRef] [PubMed]
- Hellemans, J.; Mortier, G.; De Paepe, A.; Speleman, F.; Vandesompele, J. qBase relative quantification framework and software for management and automated analysis of real-time quantitative PCR data. Genome Biol. 2007, 8, R19. [Google Scholar] [CrossRef]
- Silva, M.d.C.; Várzea, V.; Guerra-Guimarães, L.; Azinheira, H.G.; Fernandez, D.; Petitot, A.-S.; Bertrand, B.; Lashermes, P.; Nicole, M. Coffee resistance to the main diseases: Leaf rust and coffee berry disease. Brazilian J. Plant Physiol. 2006, 18, 119–147. [Google Scholar] [CrossRef]
- Denoeud, F.; Carretero-Paulet, L.; Dereeper, A.; Droc, G.; Guyot, R.; Pietrella, M.; Zheng, C.; Alberti, A.; Anthony, F.; Aprea, G.; et al. The coffee genome provides insight into the convergent evolution of caffeine biosynthesis. Science. 2014, 345, 1181–1184. [Google Scholar] [CrossRef]
- Stergiopoulos, I.; de Wit, P.J.G.M. Fungal Effector Proteins. Annu. Rev. Phytopathol. 2009, 47, 233–263. [Google Scholar] [CrossRef]
- Ramiro, D.A.; Escoute, J.; Petitot, A.-S.; Nicole, M.; Maluf, M.P.; Fernandez, D. Biphasic haustorial differentiation of coffee rust (Hemileia vastatrix race II) associated with defence responses in resistant and susceptible coffee cultivars. Plant Pathol. 2009, 58, 944–955. [Google Scholar] [CrossRef]
- Ravensdale, M.; Nemri, A.; Thrall, P.H.; Ellis, J.G.; Dodds, P.N. Co-evolutionary interactions between host resistance and pathogen effector genes in flax rust disease. Mol. Plant Pathol. 2011, 12, 93–102. [Google Scholar] [CrossRef]
- Bourras, S.; Kunz, L.; Xue, M.; Praz, C.R.; Müller, M.C.; Kälin, C.; Schläfli, M.; Ackermann, P.; Flückiger, S.; Parlange, F.; et al. The AvrPm3-Pm3 effector-NLR interactions control both race-specific resistance and host-specificity of cereal mildews on wheat. Nat. Commun. 2019, 10, 1–16. [Google Scholar] [CrossRef]
- Shalaby, S.; Horwitz, B.A. Plant phenolic compounds and oxidative stress: Integrated signals in fungal–plant interactions. Curr. Genet. 2015, 61, 347–357. [Google Scholar] [CrossRef]
- Jiang, S.; Han, S.; He, D.; Cao, G.; Fang, K.; Xiao, X.; Yi, J.; Wan, X. The accumulation of phenolic compounds and increased activities of related enzymes contribute to early defense against walnut blight. Physiol. Mol. Plant Pathol. 2019, 108, 101433. [Google Scholar] [CrossRef]
- Dobon, A.; Bunting, D.C.E.; Cabrera-Quio, L.E.; Uauy, C.; Saunders, D.G.O. The host-pathogen interaction between wheat and yellow rust induces temporally coordinated waves of gene expression. BMC Genomics 2016, 17, 380. [Google Scholar] [CrossRef]
- Jing, L.; Guo, D.; Hu, W.; Niu, X. The prediction of a pathogenesis-related secretome of Puccinia helianthi through high-throughput transcriptome analysis. BMC Bioinform. 2017, 18, 166. [Google Scholar] [CrossRef]
- Rodriguez-Moreno, L.; Ebert, M.K.; Bolton, M.D.; Thomma, B.P.H.J. Tools of the crook- infection strategies of fungal plant pathogens. Plant J. 2018, 93, 664–674. [Google Scholar] [CrossRef]
- Vieira, A.; Talhinhas, P.; Loureiro, A.; Thürich, J.; Duplessis, S.; Fernandez, D.; do Céu Silva, M.; Paulo, O.S.; Azinheira, H.G. Expression profiling of genes involved in the biotrophic colonisation of Coffea arabica leaves by Hemileia vastatrix. Eur. J. Plant Pathol. 2012, 133, 261–277. [Google Scholar] [CrossRef]
- Yin, C.; Downey, S.I.; Klages-Mundt, N.L.; Ramachandran, S.; Chen, X.; Szabo, L.J.; Pumphrey, M.; Hulbert, S.H. Identification of promising host-induced silencing targets among genes preferentially transcribed in haustoria of Puccinia. BMC Genomics 2015, 16, 579. [Google Scholar] [CrossRef]
- Quackenbush, J. Microarrays--Guilt by Association. Science 2003, 302, 240–241. [Google Scholar] [CrossRef]
- De Carvalho, M.C.d.C.G.; Costa Nascimento, L.; Darben, L.M.; Polizel-Podanosqui, A.M.; Lopes-Caitar, V.S.; Qi, M.; Rocha, C.S.; Carazzolle, M.F.; Kuwahara, M.K.; Pereira, G.A.G.; et al. Prediction of the in planta Phakopsora pachyrhizi secretome and potential effector families. Mol. Plant Pathol. 2017, 18, 363–377. [Google Scholar] [CrossRef]
- Wang, W.; Jiao, F. Effectors of Phytophthora pathogens are powerful weapons for manipulating host immunity. Planta 2019, 250, 413–425. [Google Scholar] [CrossRef]
- Marshall, R.; Kombrink, A.; Motteram, J.; Loza-Reyes, E.; Lucas, J.; Hammond-Kosack, K.E.; Thomma, B.P.H.J.; Rudd, J.J. Analysis of Two in Planta Expressed LysM Effector Homologs from the Fungus Mycosphaerella graminicola Reveals Novel Functional Properties and Varying Contributions to Virulence on Wheat. Plant Physiol. 2011, 156, 756–769. [Google Scholar] [CrossRef]
- Divon, H.H.; Fluhr, R. Nutrition acquisition strategies during fungal infection of plants. FEMS Microbiol. Lett. 2007, 266, 65–74. [Google Scholar] [CrossRef]
- Harris, L.J.; Balcerzak, M.; Johnston, A.; Schneiderman, D.; Ouellet, T. Host-preferential Fusarium graminearum gene expression during infection of wheat, barley, and maize. Fungal Biol. 2016, 120, 111–123. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hours Post-Inoculation | % of Infection Sites with Autofluorescent and/or Browning Cells in Leaves of the Coffee Genotypes | |
|---|---|---|
| Híbrido de Timor CIFC 832/1 (R) | Caturra CIFC 19/1 (S) | |
| 10 | 0 | 0 |
| 17 | 18 ± 3 | 30 ± 5 |
| 24 | 65 ± 13 | 30 ± 5 |
| 48 | 73 ± 8 | 30 ± 5 |
| 72 | 83 ± 8 | 28 ± 6 |
| 96 | 93 ± 3 | 28 ± 3 |
| Libraries | Raw Reads | Trimmed Reads | Mapped Reads against Coffee | Unmapped Reads against Coffee | H. vastatrix Reads * |
|---|---|---|---|---|---|
| Uredospores | |||||
| Hydrated (HU) | 19,095,474 | 15,120,725 | 224,132 | 14,896,593 | 10,685,403 |
| Germinated (GU) | 11,854,206 | 9,780,528 | 111,336 | 9,669,192 | 6,116,869 |
| CIFC 19/1 (S) | |||||
| 12 h | 24,937,788 | 21,535,680 | 14,662,195 | 6,873,485 | 1,040,374 |
| 24 h | 19,149,370 | 15,285,996 | 9,698,668 | 5,587,328 | 552,489 |
| 96 h | 13,416,692 | 10,677,026 | 6,380,984 | 4,296,042 | 397,579 |
| 17 d | 27,295,036 | 20,334,878 | 10,244,133 | 10,090,745 | 3,912,843 |
| CIFC 832/1 (R) | |||||
| 12 h | 22,328,618 | 19,101,578 | 14,698,398 | 4,403,180 | 666,275 |
| 24 h | 31,061,884 | 24,885,905 | 20,409,662 | 4,476,243 | 648,268 |
| 96 h | 12,359,240 | 9,438,008 | 6,460,518 | 2,977,490 | 172,604 |
| 17 d | 18,684,474 | 16,204,815 | 12,188,572 | 4,016,243 | 140,861 |
| Total | 200,182,782 | 162,365,139 | 95,078,598 | 67,286,541 | 24,333,565 |
| Experiment | Reference (Control) | Upregulated | Downregulated | Total |
|---|---|---|---|---|
| HdT (Resistant) 12 hpi | Spores a | 170 | 168 | 338 |
| HdT (Resistant) 24 hpi | Spores | 157 | 133 | 290 |
| HdT (Resistant) 96 hpi | Spores | 69 | 45 | 114 |
| HdT (Resistant) 17 dpi | Spores | 24 | 13 | 37 |
| Cat (Susceptible) 12 hpi | Spores | 248 | 203 | 451 |
| Cat (Susceptible) 24 hpi | Spores | 227 | 184 | 411 |
| Cat (Susceptible) 96 hpi | Spores | 179 | 112 | 291 |
| Cat (Susceptible) 17 dpi | Spores | 312 | 236 | 548 |
| Cat (Susceptible) 12 hpi | HdT (Resistant) 12 hpi | 263 | 154 | 417 |
| Cat (Susceptible) 24 hpi | HdT (Resistant) 24 hpi | 178 | 208 | 386 |
| Cat (Susceptible) 96 hpi | HdT (Resistant) 96 hpi | 144 | 86 | 230 |
| Cat (Susceptible) 17 dpi | HdT (Resistant) 17 dpi | 0 | 10 | 10 |
| Total | 1971 | 1552 | 3523 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castro, I.S.L.; Freitas-Lopes, R.d.L.; Ferreira, S.d.S.; Maciel, T.E.F.; Florez, J.C.; Zambolim, E.M.; Zambolim, L.; Caixeta, E.T. Transcriptome Analysis Uncovers the Gene Expression Profile of Hemileia vastatrix (Race XXXIII) during the Interactions with Resistant and Susceptible Coffee. Agronomy 2022, 12, 444. https://doi.org/10.3390/agronomy12020444
Castro ISL, Freitas-Lopes RdL, Ferreira SdS, Maciel TEF, Florez JC, Zambolim EM, Zambolim L, Caixeta ET. Transcriptome Analysis Uncovers the Gene Expression Profile of Hemileia vastatrix (Race XXXIII) during the Interactions with Resistant and Susceptible Coffee. Agronomy. 2022; 12(2):444. https://doi.org/10.3390/agronomy12020444
Chicago/Turabian StyleCastro, Isabel Samila Lima, Rejane do Livramento Freitas-Lopes, Sávio de Siqueira Ferreira, Talles Eduardo Ferreira Maciel, Juan Carlos Florez, Eunize Maciel Zambolim, Laércio Zambolim, and Eveline Teixeira Caixeta. 2022. "Transcriptome Analysis Uncovers the Gene Expression Profile of Hemileia vastatrix (Race XXXIII) during the Interactions with Resistant and Susceptible Coffee" Agronomy 12, no. 2: 444. https://doi.org/10.3390/agronomy12020444
APA StyleCastro, I. S. L., Freitas-Lopes, R. d. L., Ferreira, S. d. S., Maciel, T. E. F., Florez, J. C., Zambolim, E. M., Zambolim, L., & Caixeta, E. T. (2022). Transcriptome Analysis Uncovers the Gene Expression Profile of Hemileia vastatrix (Race XXXIII) during the Interactions with Resistant and Susceptible Coffee. Agronomy, 12(2), 444. https://doi.org/10.3390/agronomy12020444