
Wood-Ash Fertiliser and Distance from Drainage Ditch Affect the Succession and Biodiversity of Vascular Plant Species in Tree Plantings on Marginal Organic Soil




Abstract
1. Introduction
2. Materials and Methods
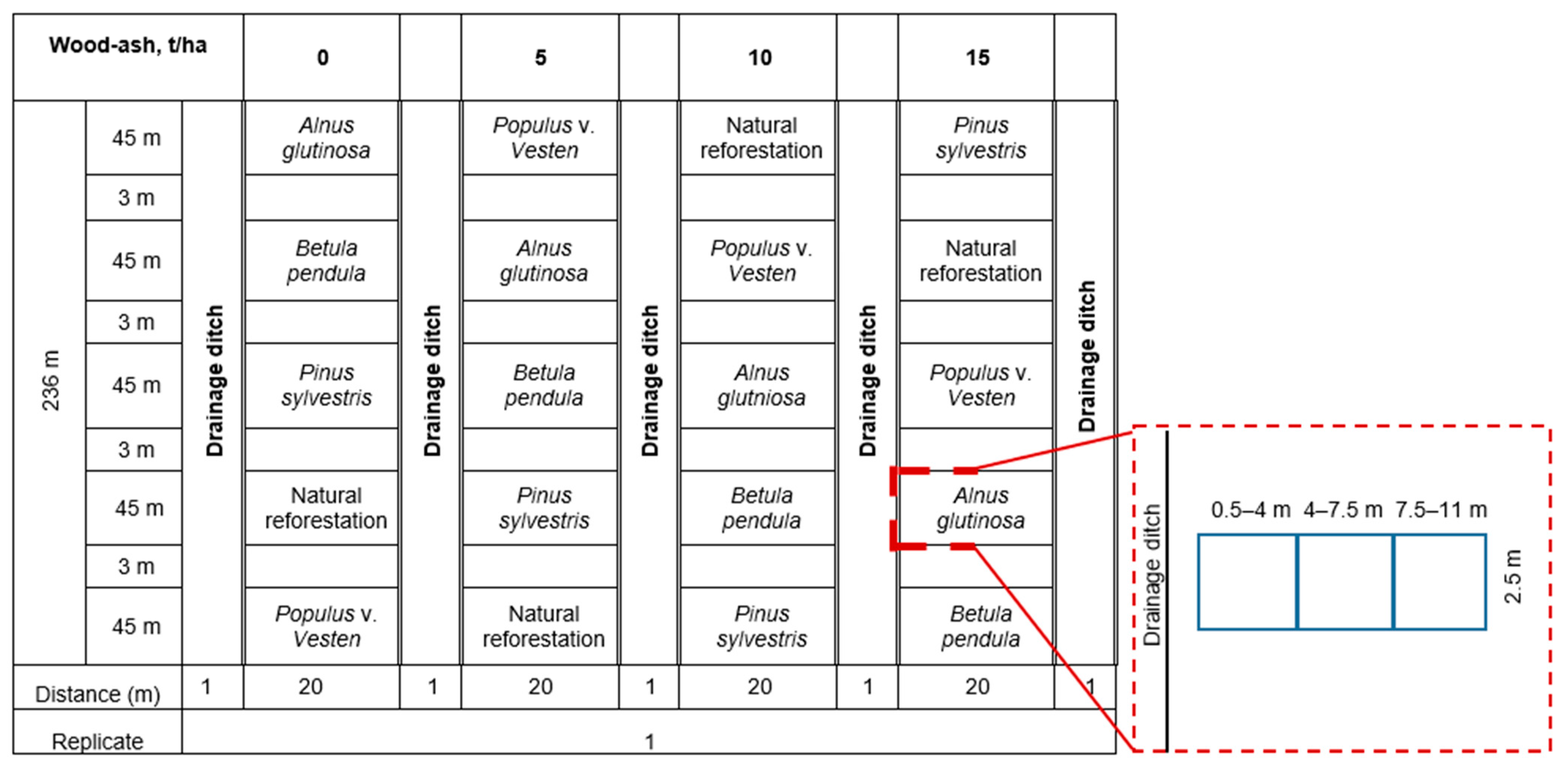
Study Site and Design
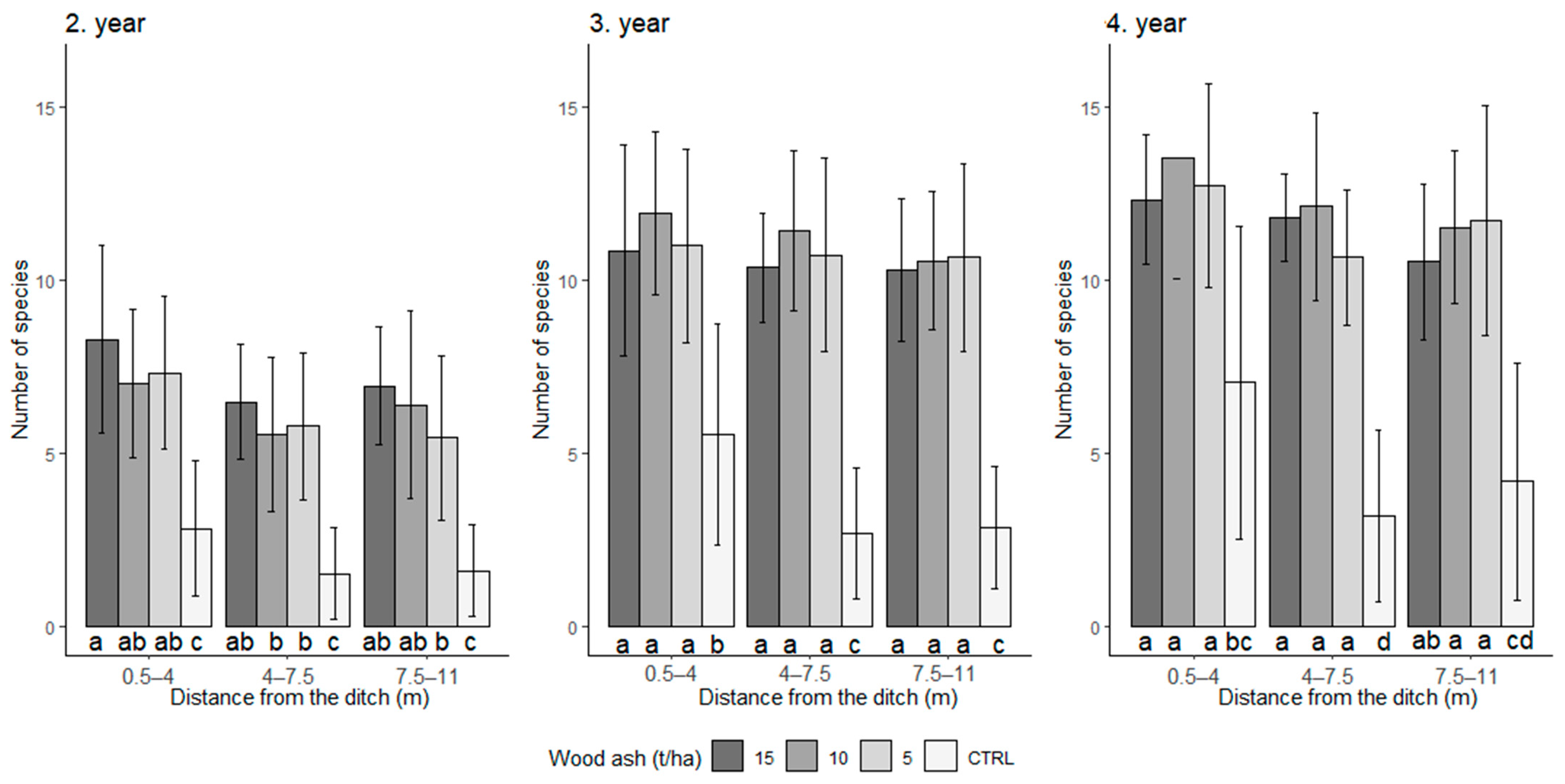
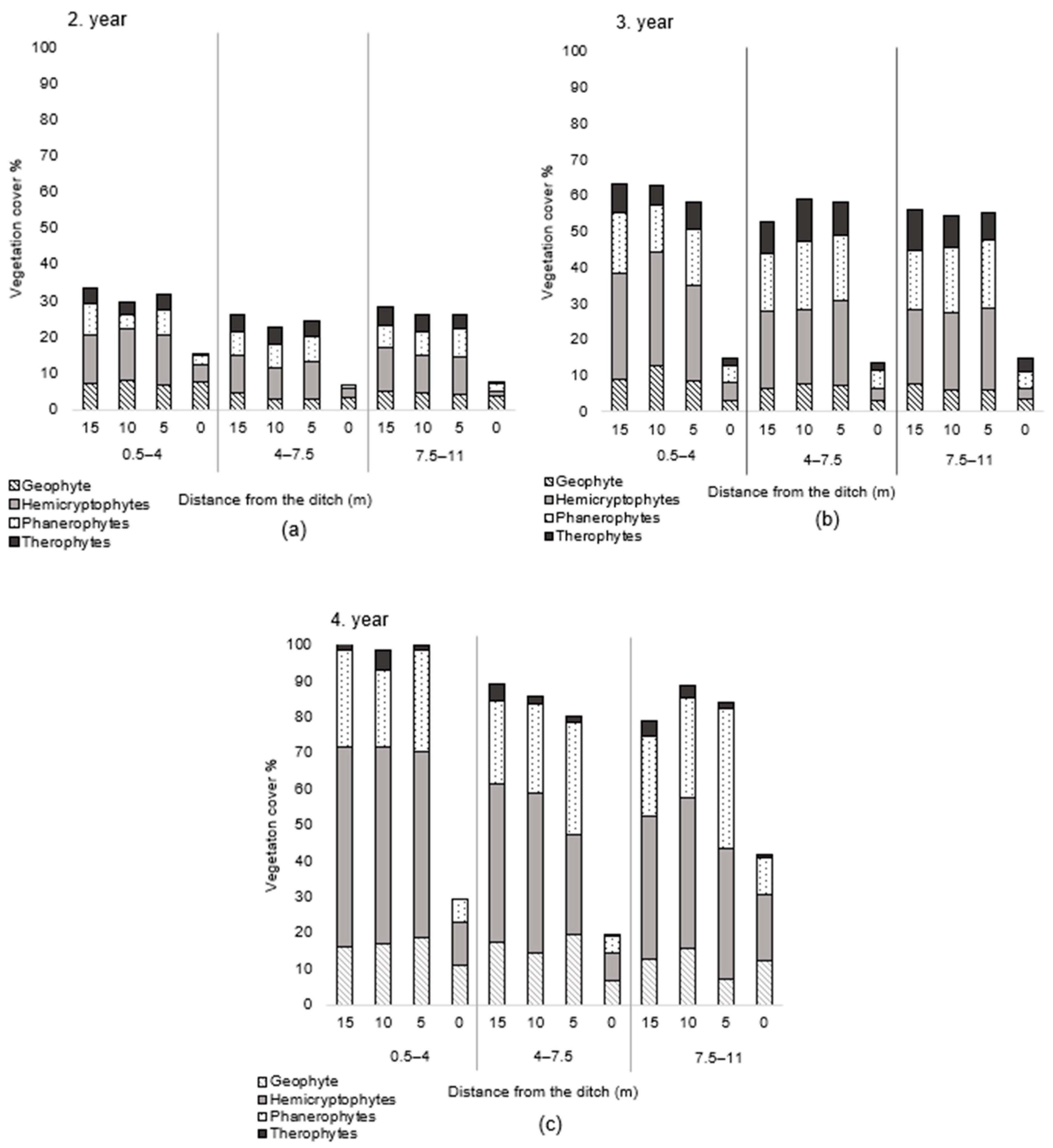
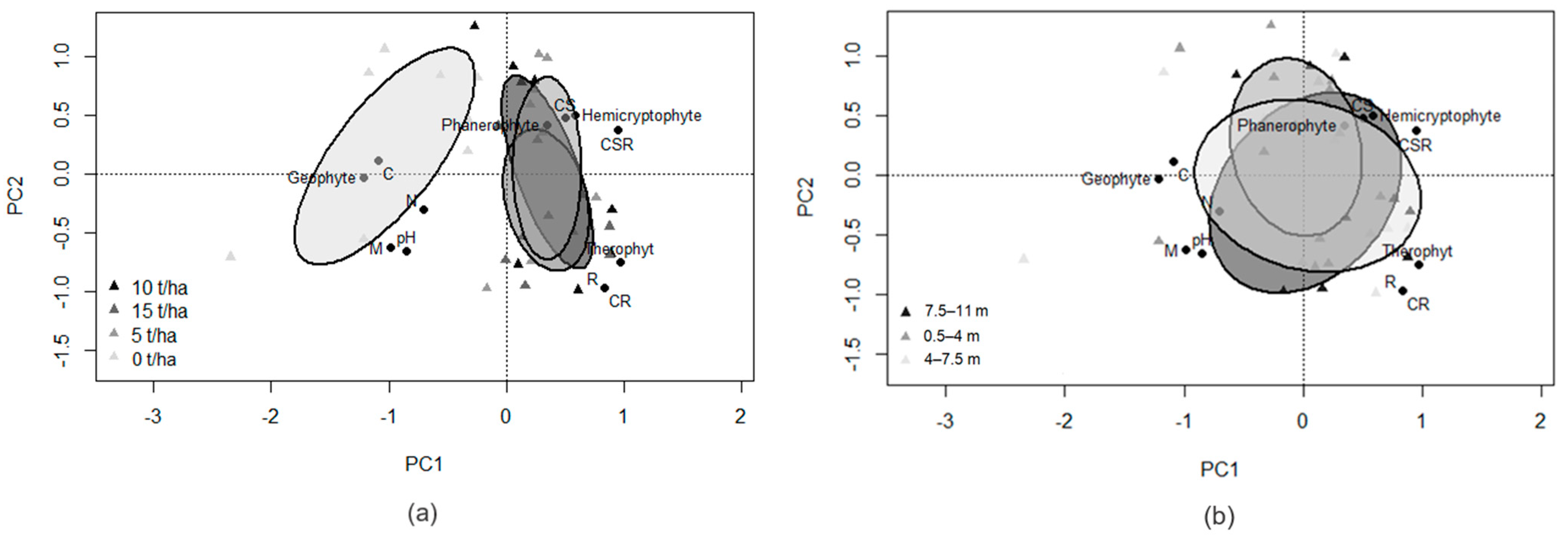
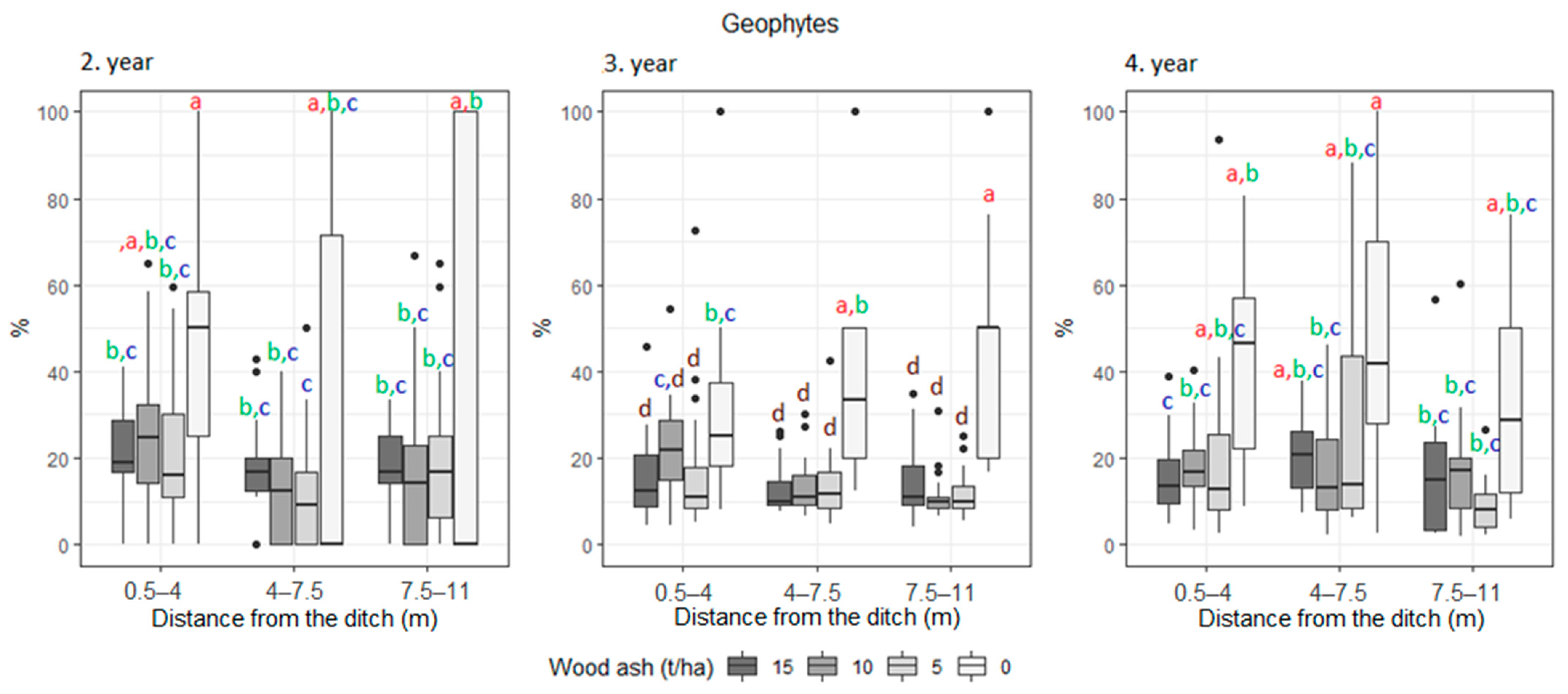
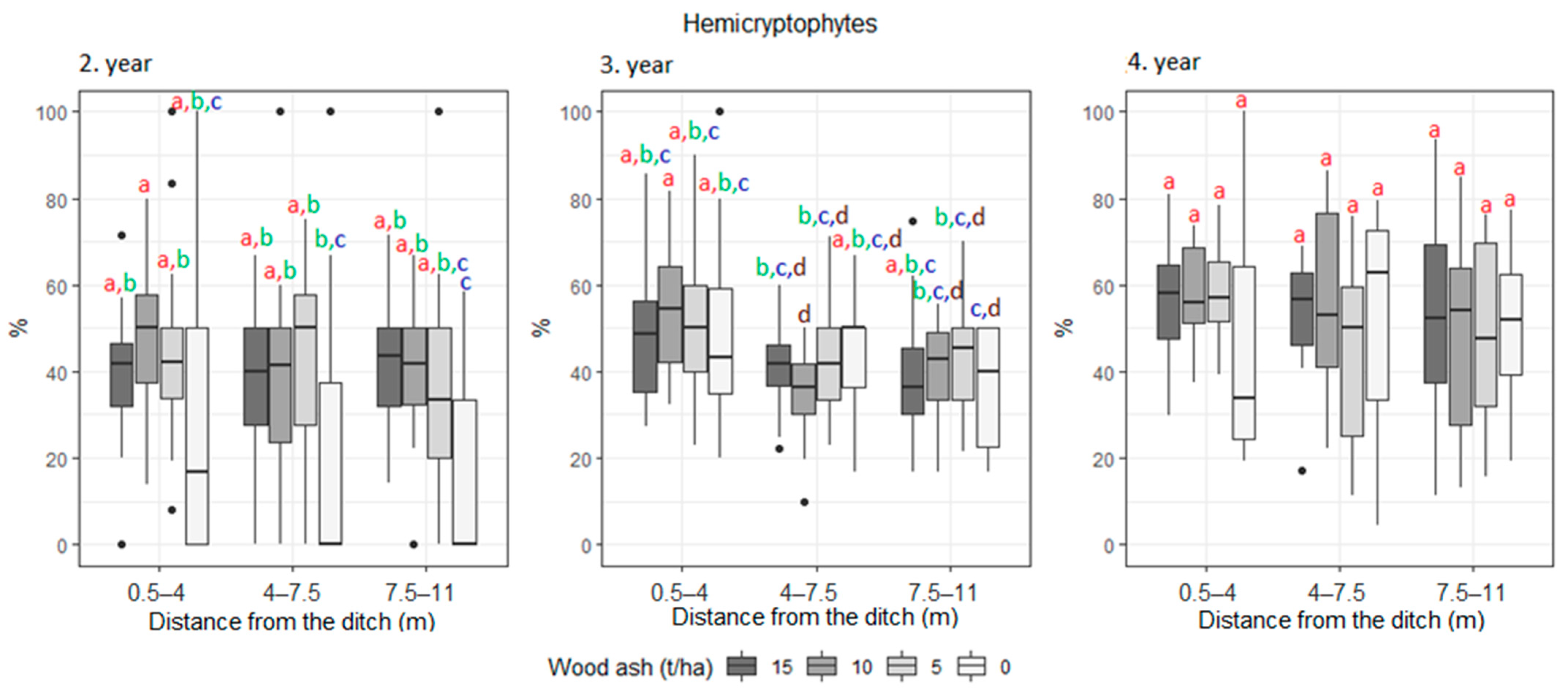
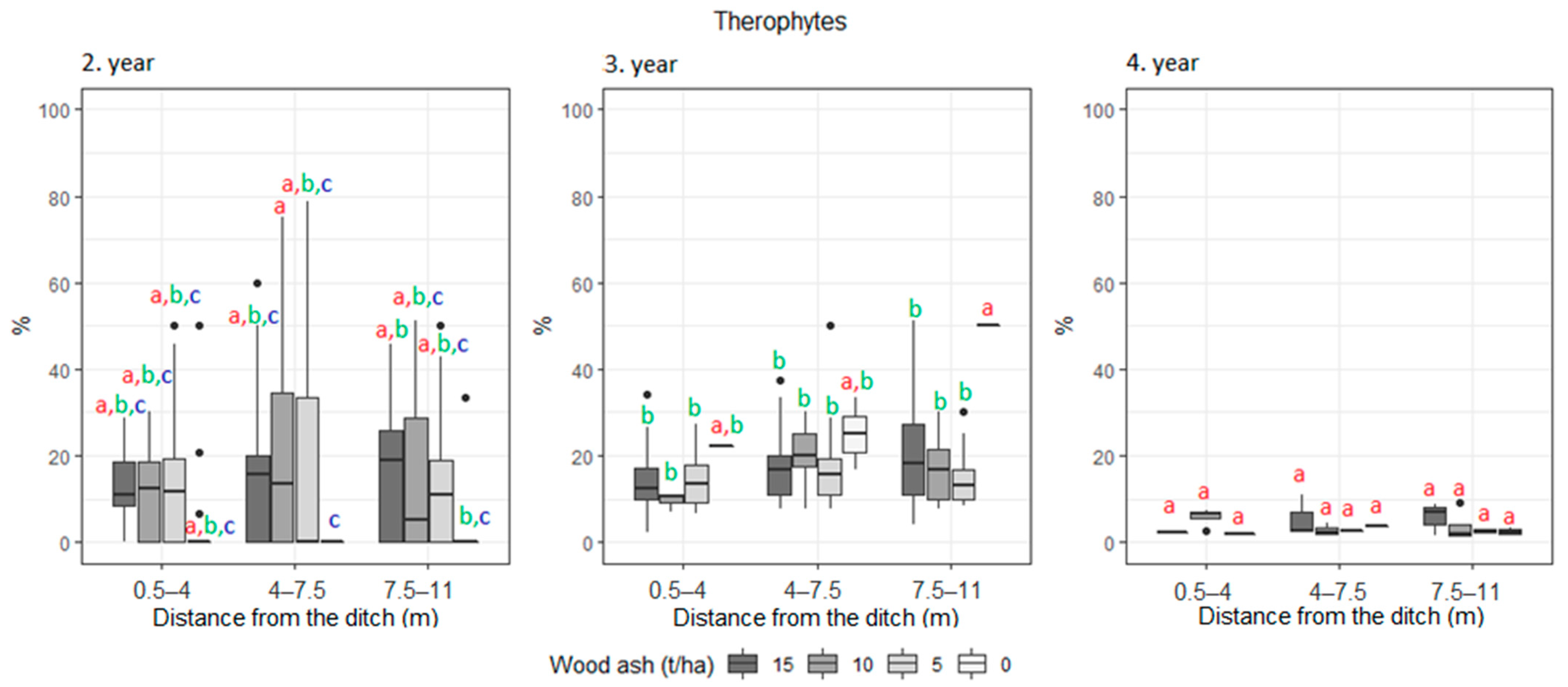
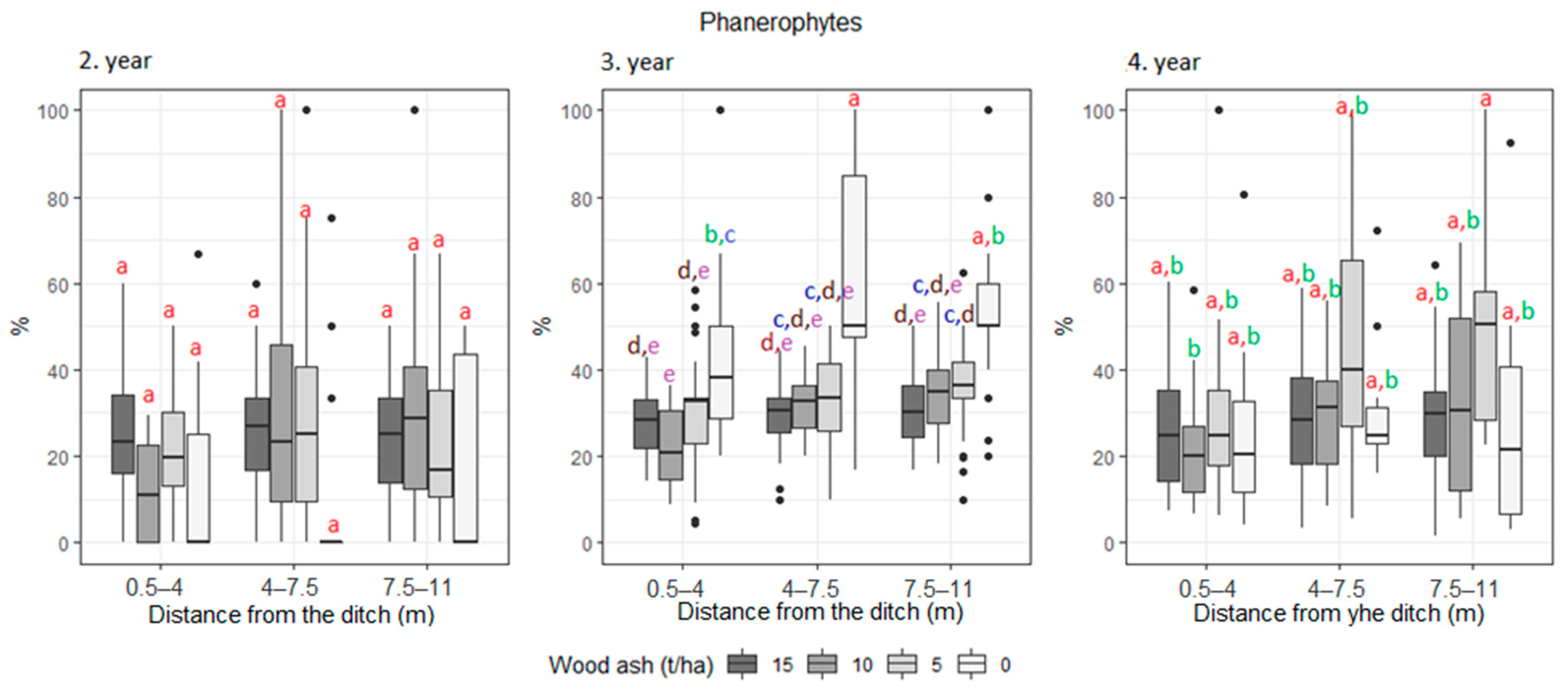
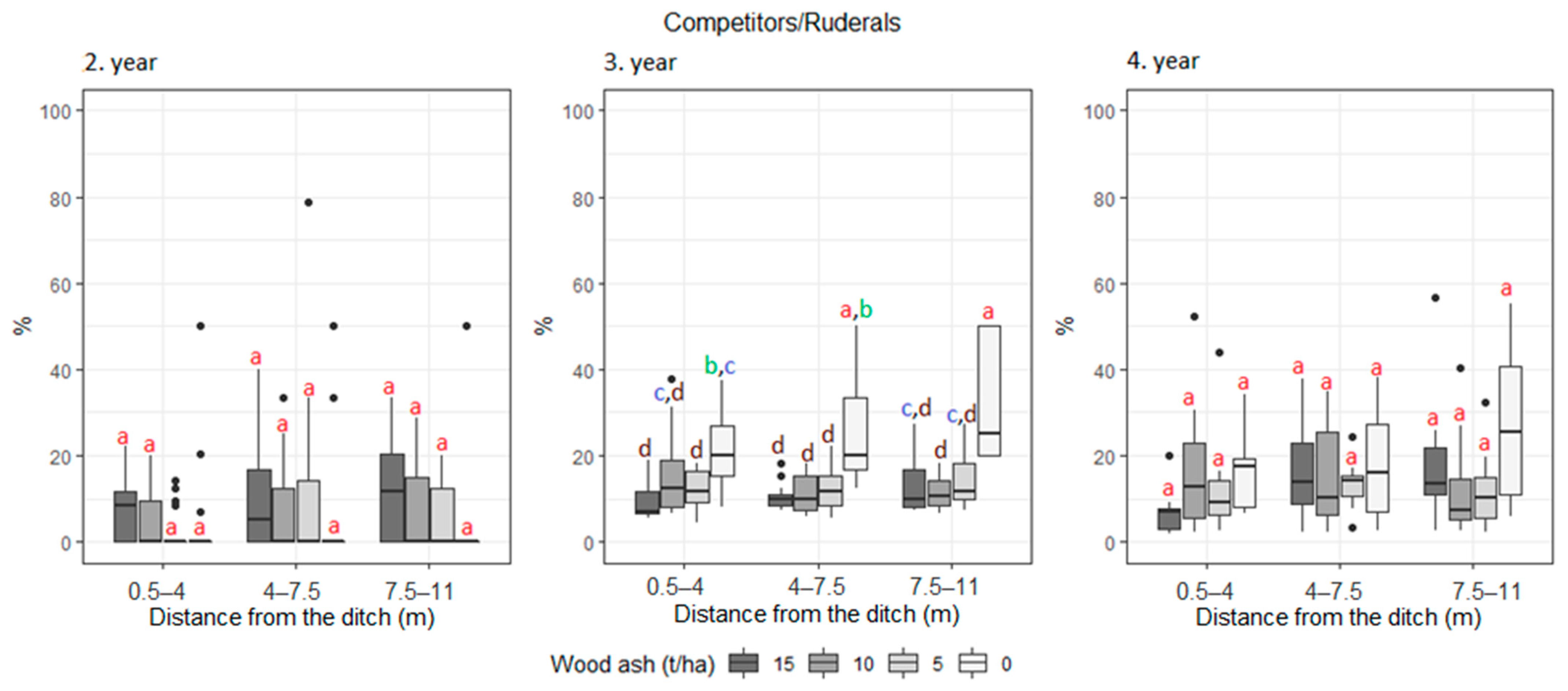
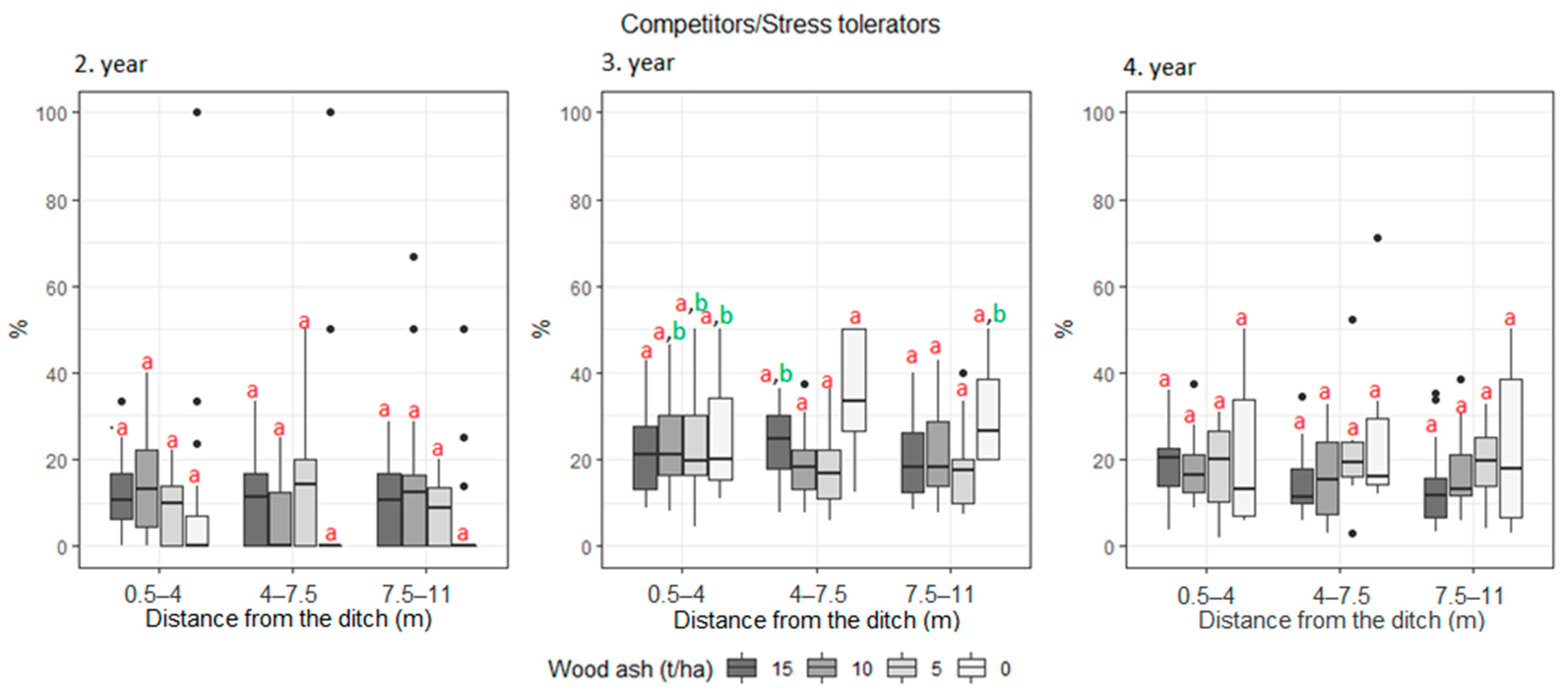
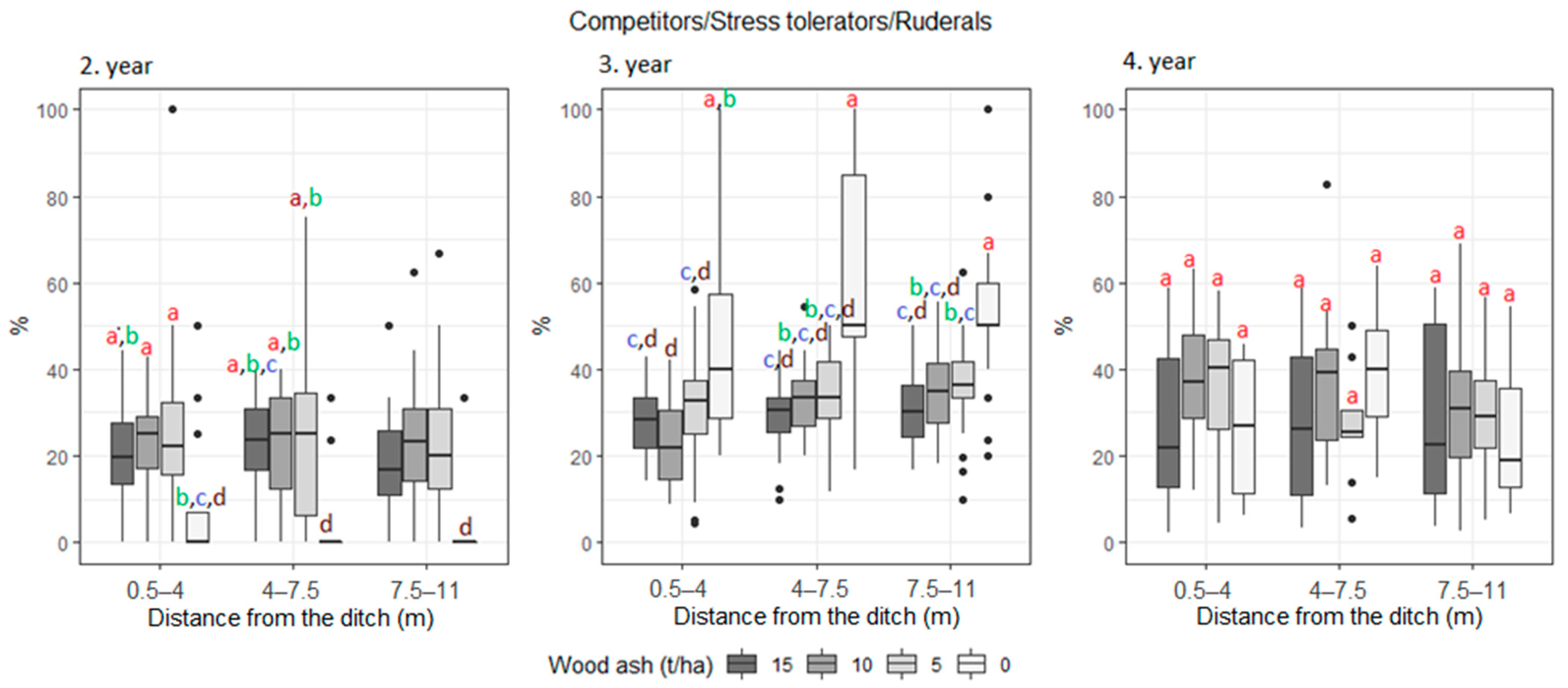
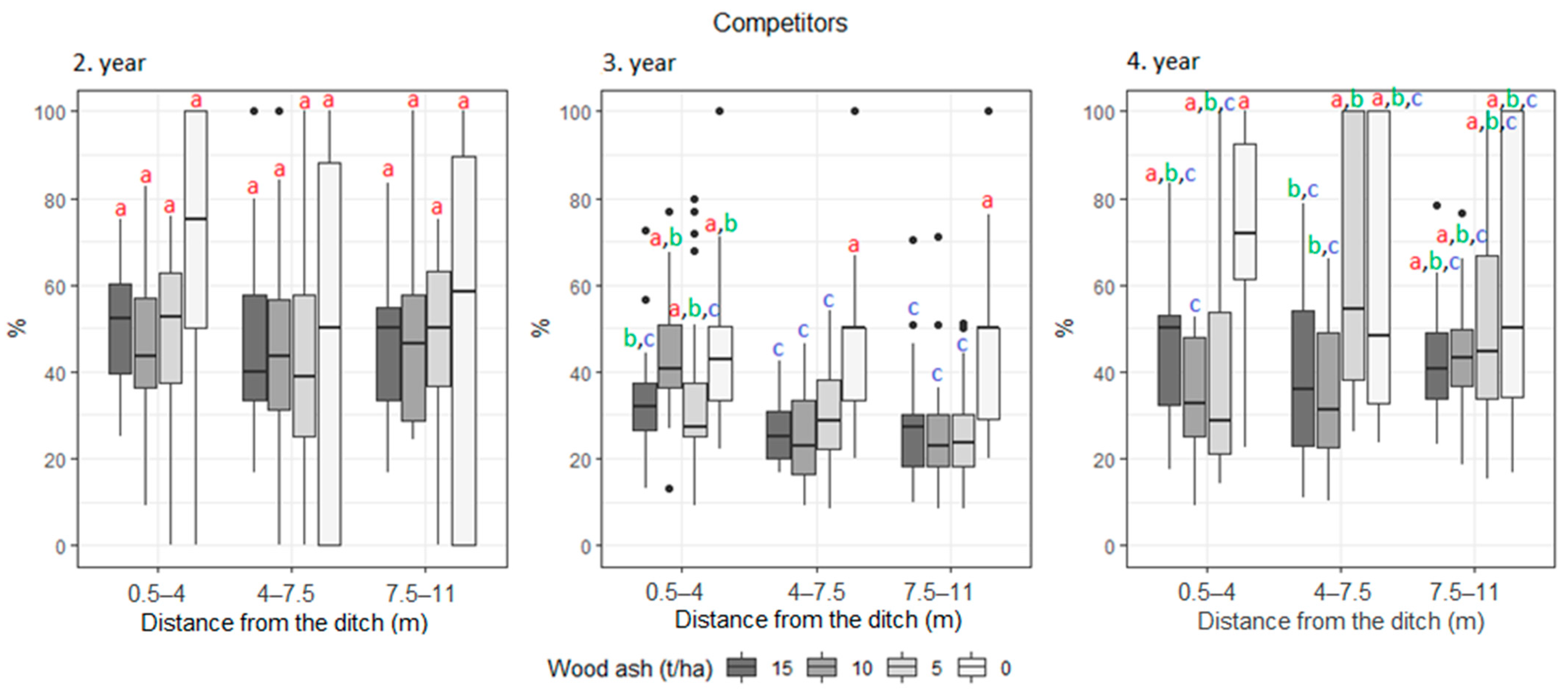
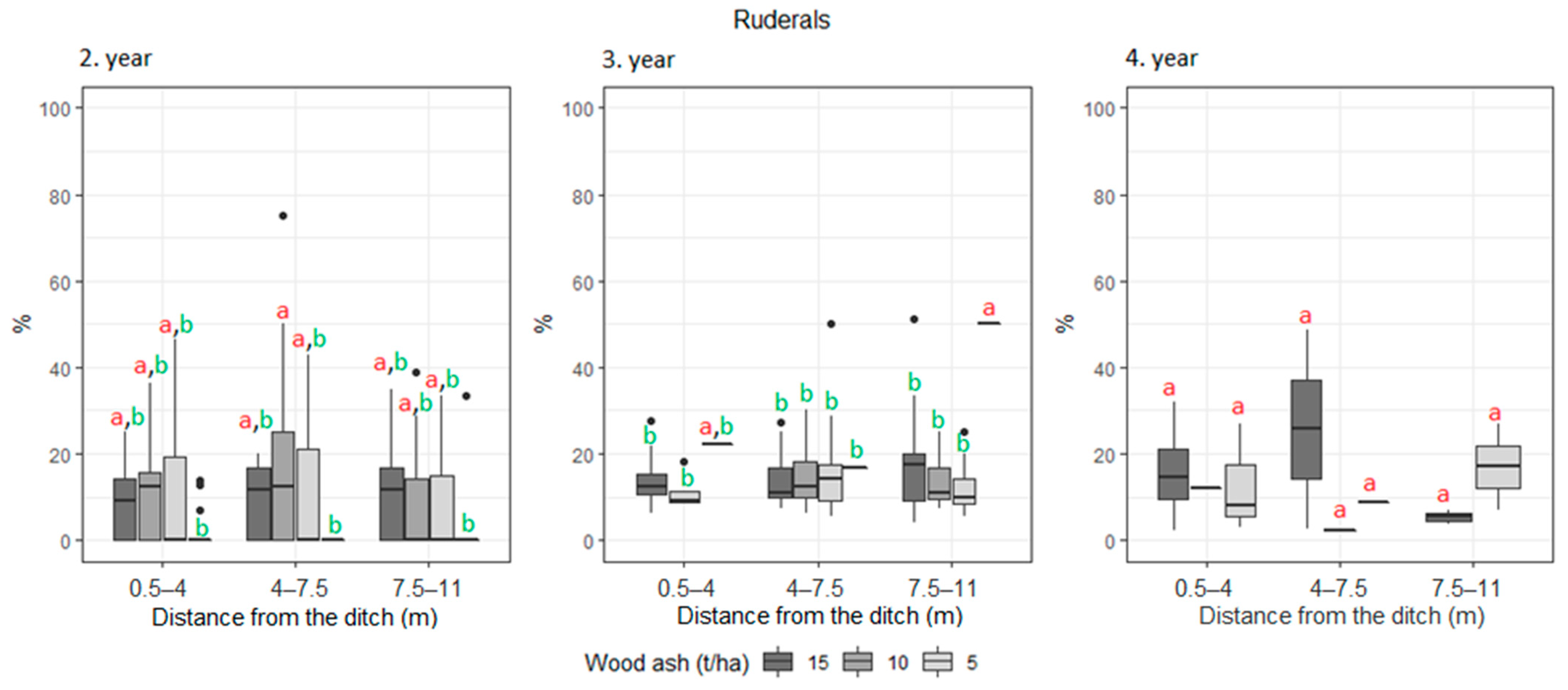
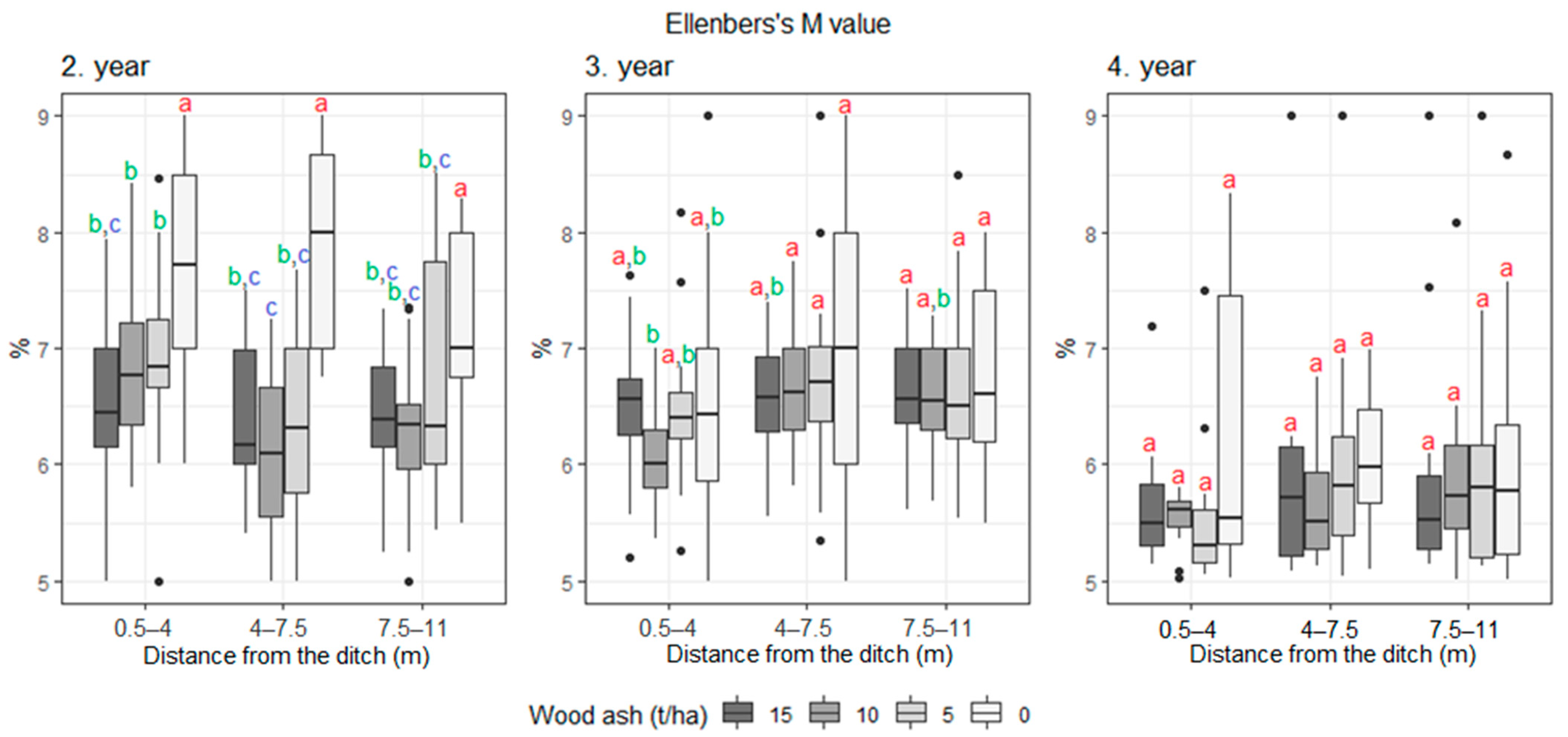
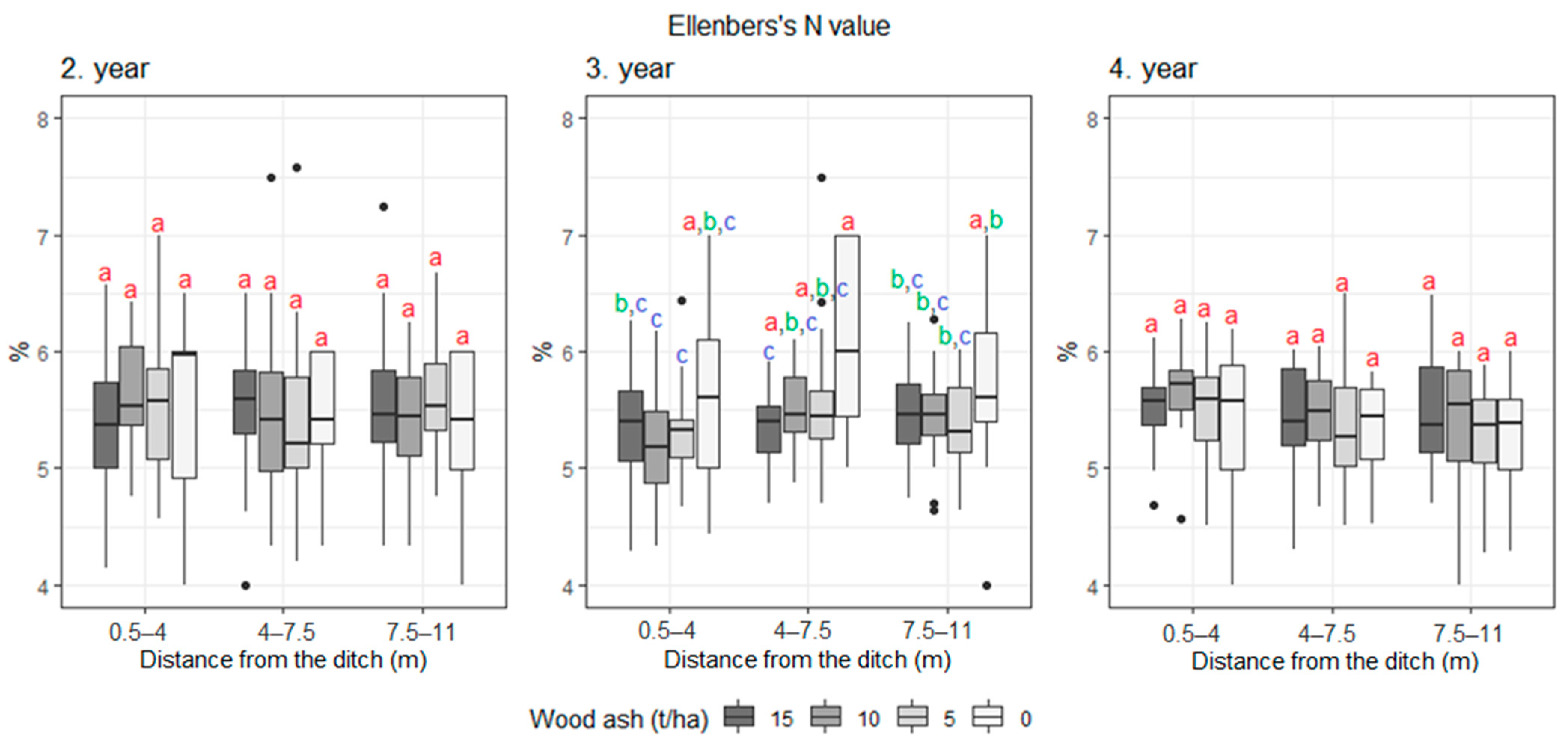
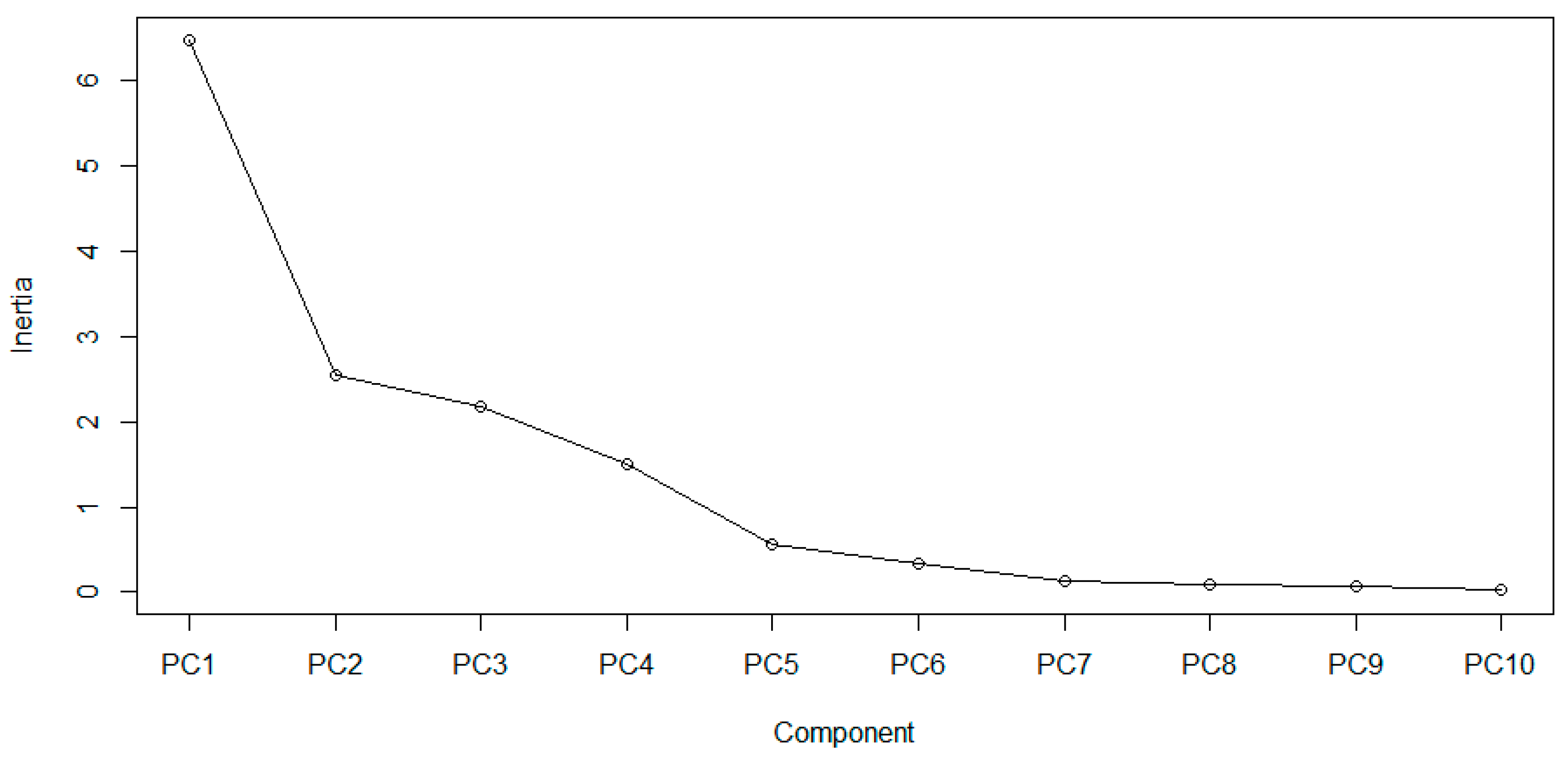
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Species | 2019 | 2018 | 2017 | No. | Species | 2019 | 2018 | 2017 |
|---|---|---|---|---|---|---|---|---|---|
| 1. | Agrostis capillaris | x | x | x | 43. | Luzula pilosa | x | ||
| 2. | Arabidopsis thaliana | x | x | 44. | Lycopus europaeus | x | x | x | |
| 3. | Arctium lappa | x | x | 45. | Matricaria perforata | x | x | ||
| 4. | Arctium tomentosum | x | 46. | Mycelis muralis | x | ||||
| 5. | Barbara stricta | x | 47. | Persicaria maculosa | x | ||||
| 6. | Betula pendula | x | x | x | 48. | Petasites hybridus | x | ||
| 7. | Betula pubescens | x | x | x | 49. | Phragmites australis | x | x | x |
| 8. | Bidens tripartita | x | x | x | 50. | Picea abies | x | x | x |
| 9. | Brassica campestris | x | x | 51. | Picris hieracioides | x | x | x | |
| 10. | Calamagrostis canescens | x | x | x | 52. | Pinus sylvestris | x | x | x |
| 11. | Calluna vulgaris | x | x | 53. | Plantago lanceolata | x | |||
| 12. | Carex cespitosa | x | x | x | 54. | Plantago major | x | x | |
| 13. | Carex hirta | x | 55. | Poa palustris | x | x | x | ||
| 14. | carex pseudocyperus | x | 56. | Polygonum sp. | x | ||||
| 15. | Carex vesicaria | x | 57. | Polytrihum sp. | x | x | |||
| 16. | Cerastium holosteoides | x | x | 58. | Populus tremula | x | x | x | |
| 17. | Chamerion angustifolium | x | x | x | 59. | Rubus idaeus | x | x | x |
| 18. | Chenopodium album | x | x | 60. | Rumex acetosa | x | x | ||
| 19. | Cirsium arvense | x | x | x | 61. | Rumex acetosella | x | x | x |
| 20. | Cirsium aucale | x | 62. | Rumex longifolius | x | ||||
| 21. | Cirsium oleraceum | x | x | 63. | sagina nodosa | x | |||
| 22. | Cirsium palustre | x | 64. | Salix alba | x | ||||
| 23. | Crepis biennis | x | x | 65. | Salix caprea | x | x | x | |
| 24. | Echinochloa crusgalli | x | x | 66. | Salix myrsinifolia | x | |||
| 25. | Eirophorum polystachion | x | 67. | Salix rosmarinifolia | x | ||||
| 26. | Epilobium parviflorum | x | x | x | 68. | Salix sp. | x | x | |
| 27. | Equisetum arvense | x | 69. | Salix starkeana | x | ||||
| 28. | Equisetum fluviatile | x | 70. | Salix triandra | x | ||||
| 29. | Equisetum sylvaticum | x | 71. | Salix viminalis | x | ||||
| 30. | Erigon canadesis | x | x | x | 72. | Scirpus sylvaticus | x | ||
| 31. | Eriophorum vaginatum | x | x | x | 73. | Senecio sylvaticus | x | x | |
| 32. | Eupatorium cannabinum | x | x | x | 74. | Silene vulgaris | x | ||
| 33. | Festuca rubra | x | 75. | Solidago cannadensis | x | x | x | ||
| 34. | Fragaria vesca | x | x | x | 76. | Sonchus asper | x | x | |
| 35. | Frangula alnus | x | x | 77. | Stellaria media | x | x | ||
| 36. | Gnaphalium uliginosum | x | 78. | Taraxacum officinale | x | x | x | ||
| 37. | Hieracium pilosella | x | x | x | 79. | Trifolium repens | x | ||
| 38. | Juncus articulatus | x | x | x | 80. | Tripleurospermum inodorum | x | ||
| 39. | Juncus effusus | x | x | x | 81. | Tussilafgo farfara | x | x | x |
| 40. | Juncus tenuis | x | x | x | 82. | Typha latifolia | x | x | x |
| 41. | Lamium album | x | x | 83. | Utrica dioica | x | |||
| 42. | Linaria vulgaris | x | x | 84. | Valeriana officinalis | x |
References
- Xu, J.; Morris, P.J.; Liu, J.; Holden, J. PEATMAP: Refining Estimates of Global Peatland Distriubution Based on a Meta-Analysis. Catena 2018, 160, 134–140. [Google Scholar] [CrossRef]
- Clymo, R.S. The Limits to Peat Bog Growth. Philosophical Transactions of the Royal Society of London. Ser. B Biol. Sci. 1984, 303, 605–654. [Google Scholar] [CrossRef]
- Yli-Petays, M.; Laine, J.; Vasander, H.; Tuittila, E.-S. Carbon Gas Exchange of a Re-Vegetated Cut-Away Peatland Five Decades after Abandonment. For. Ecol. Manag. 2007, 12, 177–190. [Google Scholar]
- Lazdiņš, A.; Lupikis, A. Life Restore Project Contribution to the Greenhouse Gas Emission Accounts in Latvia. In Sustainable and Responsible After-Use of Peat Extraction Areas; Baltijas krasti: Riga, Latvia, 2019; pp. 21–52. [Google Scholar]
- Huotari, N.; Tillman-Sutela, E.; Kauppi, A.; Kubin, E. Fertilization Ensures Rapid Formation of Ground Vegetation on Cut-Away Peatlands. Can. J. For. Res. 2006, 37, 874–883. [Google Scholar] [CrossRef]
- Tuittila, E.-S.; Komulainen, V.-M.; Vasander, H.; Laine, J. Restored Cut-Away Peatland as a Sink for Atmospheric CO2. Oecologia 1999, 120, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Pētersons, J.; Lazdiņš, A.; Kasakovskis, A. Life Restore Database on Areas Affected by Peat Extraction. In Sustainable and Responsible After-Use of Peat Extraction Areas; Baltijas krasti: Riga, Latvia, 2019; pp. 122–128. [Google Scholar]
- Mehmood, M.A.; Ibrahim, M.; Rashid, U.; Nawaz, M.; Shafaqat, A.; Hussain, A.; Gull, M. Biomass Production for Bioenergy Using Marginal Lands. Sustain. Prod. Consum. 2017, 9, 3–21. [Google Scholar] [CrossRef]
- Gong, J.; Shurpali, N.J.; Kellomäki, S.; Wang, K.; Zhang, C.; Salem, M.M.A. High Sensitivity of Peat Moisture Content to Seasonal Climate in Acutaway Peatland Cultivated with a Perennial Crop (Phalarisarundinaceae, L.): A Modeling Study. Agric. For. Meteorol. 2012, 180, 225–235. [Google Scholar] [CrossRef]
- Ots, K.; Orru, M.; Tilk, M.; Kuura, L.; Aguraijuja, K. Afforestation of Cutaway Peatlands: Effect of Wood Ash on Biomass Formation and Carbon Balance. For. Stud. 2017, 67, 17–36. [Google Scholar] [CrossRef]
- Padur, K.; Ilomets, M.; Põder, T. Identification of the Criteria for Decision Making of Cut-Away Peatland Reuse. Environ. Manag. 2017, 59, 505–521. [Google Scholar] [CrossRef]
- Neimane, S.; Celma, S.; Butlers, A.; Lazdiņa, D. Conversion of an Industrial Cutaway Peatland to a Betulacea Family Tree Species Plantation. Agron. Res. 2019, 17, 741–753. [Google Scholar] [CrossRef]
- Kikamägi, K.; Ots, K.; Kuznetsova, T.; Pototski, A. The Growth and Nutrients Status of Conifers on Ash-Treated Cutaway Peatland. Trees Struct. Funct. 2013, 28, 53–64. [Google Scholar] [CrossRef]
- Neimane, S.; Celma, S.; Zusevica, A.; Lazdina, D.; Ievinsh, G. The Effect of Wood Ash Application on Growth, Leaf Morphological and Physiological Traits of Trees Planted in a Cutaway Peatland. Mires Peat 2021, 27, 12. [Google Scholar] [CrossRef]
- Hytönen, J.; Aro, L. Biomass and Nutrition of Naturally Regenerated and Coppiced Birch on Cutaway Peatland During 37 Years. Silva Fenn. 2012, 46, 377–394. [Google Scholar] [CrossRef][Green Version]
- Wilson, L.; Wilson, J.M.; Holden, J.; Johnstone, I.; Armstrong, A.; Morris, M. Recovery of Water Tables in Welsh Blanket Bog after Drain Blocking: Discharge Rates, Time Scales and the Influence of Local Conditions. J. Hydrol. 2010, 391, 377–386. [Google Scholar] [CrossRef]
- Gatis, N.; Luscombe, D.J.; Grand-Clement, E.; Hartley, I.P.; Anderson, K.; Smith, D.; Brazier, R.E. The Effect of Drainage Ditches on Vegetation Diversity and CO2 Fluxes in a Molinia caerulea- Dominated Peatland. Ecohydrology 2016, 9, 407–420. [Google Scholar] [CrossRef]
- Renou-Wilson, F.; Farrell, E.P. Reclaiming Peatlands for Forestry. In Restoration of Boreal and Temperate Forests; CRC Press: New York, NY, USA, 2004; pp. 541–557. [Google Scholar]
- Strack, M.; Keith, A.M.; Xu, B. Growing Season Carbon Dioxide and Methane Exchange at a Restored Peatland on the Western Boreal Plain. Ecol. Eng. 2014, 64, 231–239. [Google Scholar] [CrossRef]
- Silvan, N.; Hytönen, J. Impact of Ash-Fertilization and Soil Preparation on Soil Respiration and Vegetation Colonization on Cutaway Peatlands. Am. J. Clim. Change 2016, 5, 178–192. [Google Scholar] [CrossRef]
- Kelemen, K.; Mihók, B.; Galhidy, L.; Standovár, T. Dynamic Response of Herbaceous Vegetation to Gap Opening in a Central European Beech Stand. Silva Fenn. 2012, 46, 53–65. [Google Scholar] [CrossRef]
- Thrippleton, T.; Bugmann, H.; Kramer-Priewasser, K.; Snell, R.S. Herbaceous Understorey: An Overlooked Player in Forest Landscape Dynamics? Ecosystems 2016, 19, 1240–1254. [Google Scholar] [CrossRef]
- Gilliam, F.S. Excess Nitrogen in Temperate Forest Ecosystems Decreases Herbaceous Layer Diversity and Shifts Control from Soil to Canopy Structure. Forests 2019, 10, 66. [Google Scholar] [CrossRef]
- Augusto, L.; Dupouey, J.-L.; Ranger, J. Effects of Tree Species on Understory Vegetation and Environmental Conditions in Temperate Forests. Ann. For. Sci. 2003, 60, 823–831. [Google Scholar] [CrossRef]
- Kou, M.; García-Fayos, P.; Hu, S.; Jiao, J. The Effect of Robinia pseudoacacia Afforestation on Soil and Vegetation Properties in the Loess Plateau (China): A Chronosequence Approach. For. Ecol. Manag. 2016, 375, 146–158. [Google Scholar] [CrossRef]
- Huotari, N.; Tillman-Sutela, E.; Kubin, E. Ground Vegetation Has a Major Role in Element Dynamics in an Ash-Fertilized Cut-Away Peatland. For. Ecol. Manag. 2011, 261, 2081–2088. [Google Scholar] [CrossRef]
- Mabry, C.M.; Golay, M.G.; Thompson, J.R. Seasonal Storage of Nutrients by Perennial Herbaceous Species in Undisturbed and Disturbed Deciduous Forests. Appl. Veg. Sci. 2009, 11, 37–44. [Google Scholar] [CrossRef]
- Weigel, R.; Gilles, J.; Klisz, M.; Manthey, M.; Kreyling, J. Forest Understory Vegetation Is More Related to Soil than to Climate towards the Cold Distribution Margin of European Beech. J. Veg. Sci. 2019, 30, 746–755. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, H.Y.H.; Tayor, A.R. Positive Species Diversity and Above-ground Biomass Relationships Are Ubiquitous across Forest Strata despite Interference from Overstory Trees. Funct. Ecol. 2017, 31, 419–426. [Google Scholar] [CrossRef]
- Burgess-Conforti, J.R.; Moore, P.A., Jr.; Owens, P.R.; Miller, D.M.; Ashworth, A.J.; Hays, P.D.; Evans-White, M.A.; Anderson, K.R. Are Soils beneath Coniferous Tree Stands More Acidic than Soils beneath Deciduous Tree Stands? Environ. Sci. Pollut. Res. 2019, 26, 14920–14929. [Google Scholar] [CrossRef]
- Lazdiņa, D.; Neimane, S.; Celma, S. Afforestation Demo Site. In Sustainable and Responsible After-Use of Peat Extraction Areas; Baltijas krasti: Riga, Latvia, 2019; pp. 208–220. [Google Scholar]
- Ellenberg, H.; Weber, H.E.; Düll, R.; Wirth, V.; Werner, W.; Paulißen, D. Zeigerwerte von Pflanzen in Mitteleuropa, 2nd ed.; Verlag Erich GoltzeKG: Göttingen, Germany, 1991. [Google Scholar]
- Chytrý, M.; Tichý, L.; Dřevojan, P.; Sádlo, J.; Zelený, D. Ellenberg-Type Indicator Values for the Czech Flora. Preslia 2018, 90, 83–103. [Google Scholar] [CrossRef]
- Diekmann, M. Use and Improvement of Ellenberg’s Indicator Values in Decidous Forests of Boreal-Nemoral Zone in Sweeden. Ecography 1995, 18, 178–189. [Google Scholar] [CrossRef]
- Kleyer, M.; Bekker, R.M.; Knevel, I.C.; Bakker, J.P.; Sonnenschein, M.; Poschlod, P.; van Groenendael, J.; Klimeš, L.; Klimešová, J.; Klotz, S.; et al. The LEDA Traitbase: A Database of Life History Traits of Northwest European Flora. J. Ecol. 2008, 96, 1266–1274. [Google Scholar] [CrossRef]
- Niklas, K. Life Forms. In Encyclopedia of Ecology; Elsevier: Amsterdam, The Netherlands, 2008; pp. 2160–2167. [Google Scholar]
- Saugier, B. Plant Strategies and Vegetation Processes. Plant Sci. 2001, 161, 813. [Google Scholar] [CrossRef]
- RStudio Team. RStudio: Integrated Development for R; RStudio: Boston, MA, USA, 2019. [Google Scholar]
- R Core Team. R: A Language and Environment; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan Community Ecology Package Version 2.5-7. November 2020. Available online: https://cran.r-project.org (accessed on 10 December 2021).
- Bellamy, P.E.; Stephen, L.; Maclean, L.S.; Grant, M.C. Response of Blanket Bog Vegetation to Drain-Blocking. Appl. Veg. Sci. 2012, 15, 129–135. [Google Scholar] [CrossRef]
- Price, J.; Heathwaite, A.L.; Baird, A.J. Hydrological Processes in Abandoned and Restored Peatlands: An Overview of Management Approaches. Wetl. Ecol. Manag. 2003, 11, 65–83. [Google Scholar] [CrossRef]
- Leung, A.K.; Garg, A.; Coo, J.L.; Ng, C.W.W.; Hau, B.C.H. Effects of the Roots of Cynodon dactylon and Schefflera heptaphylla on Water Infiltration Rate and Soil Hydraulic Conductivity. Hydrol. Process. 2015, 29, 3342–3354. [Google Scholar] [CrossRef]
- Campbell, D.R.; Lavoie, C.; Rochefort, L. Wind Erosion and Surface Stability in Abandoned Milled. Can. J. Soil Sci. 2002, 82, 89–95. [Google Scholar] [CrossRef]
- Dafni, A.; Cohen, D.; Noy-Mier, I. Life-Cycle Variation in Geophytes. Ann. Mo. Bot. Gard. 1981, 68, 652–660. [Google Scholar] [CrossRef]
- Marks, M.; Lapin, B.; Randall, J. Phragmites australis (P. communis): Threats, Management and Monitoring. Nat. Areas J. 1994, 6, 285–294. [Google Scholar] [CrossRef]
- Myerscough, P.J.; Whitehead, F. Comparative Biology of Tussilago farfara L., Chamaenerion angustifolium (L.) Scop., Epilobium montanum L., and Epilobium adenocaulon Hausskn. II. Growth and Ecology. New Phytol. 1967, 66, 785–823. [Google Scholar] [CrossRef]
- Schaffers, A.P.; Sýkora, K.V. Reliability of Ellenberg Indicator Values for Moisture, Nitrogen and Soil Reaction: A Comparison with Field Measurements. J. Veg. Sci. 2000, 11, 225–244. [Google Scholar] [CrossRef]






| Plant Functional Traits | Wood-Ash Dose (t/ha) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 5 | 10 | 15 | 0 | 5 | 10 | 15 | 0 | 5 | 10 | 15 | ||
| 2. Year | 3. Year | 4. Year | |||||||||||
| UAST (%) | Competitors | 50.4 | 53.8 | 47.1 | 44.9 | 27.8 | 34.6 | 35.9 | 26.7 | 51.7 | 53.0 | 45.5 | 46.4 |
| Ruderals | 5.5 | 9.2 | 8.2 | 12.3 | 7 | 6.4 | 3.5 | 12.4 | 0 | 1 | 1.7 | 1.4 | |
| Competitors/Ruderals Ruderals | 5.3 | 7.6 | 5.3 | 9.1 | 7.3 | 6.8 | 7.3 | 6.4 | 10.2 | 9.7 | 8.8 | 11.1 | |
| Competitors/Stress tolerant Stress tolerant | 6.2 | 11.6 | 11.7 | 10.4 | 13.5 | 17.4 | 16.9 | 17.9 | 14.6 | 13.2 | 16.6 | 13.5 | |
| Competitors/Stress tolerant/Ruderals | 17.9 | 15.7 | 19.9 | 22.6 | 39.6 | 31.3 | 32.0 | 32.0 | 22.6 | 20.6 | 27.3 | 27.5 | |
| Growth form (%) | Geophyte | 23.7 | 26.7 | 24.6 | 13.2 | 15.5 | 16.2 | 18.6 | 12.5 | 19.2 | 23.8 | 22.9 | 17.5 |
| Therophyte | 8.3 | 13.7 | 9.7 | 18.1 | 10.0 | 9.2 | 4.6 | 16.4 | 0.6 | 0.4 | 0.8 | 0.6 | |
| Hemicryptophyte | 30.5 | 35.7 | 40.9 | 40.3 | 33.7 | 42.6 | 42.2 | 38.6 | 43.3 | 46.1 | 51.3 | 47.0 | |
| Phanerophyte | 22.7 | 22.1 | 17.5 | 28.2 | 39.7 | 30.5 | 32.1 | 31.3 | 36.9 | 27.3 | 25.0 | 34.9 | |
| Ellenberg’s value | Moisture | 6.6 | 6.9 | 6.5 | 6.2 | 6.5 | 6.5 | 6.2 | 6.6 | 6.9 | 6.8 | 6.9 | 7.0 |
| Nitrogen | 5.0 | 5.3 | 5.1 | 5.3 | 5.4 | 5.4 | 5.3 | 5.4 | 5.9 | 5.5 | 5.6 | 5.8 | |
| Plant Functional Traits | Distance from Drainage Ditch (m) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 0.5–4 | 4–7.5 | 7.5–11 | 0.5–4 | 4–7.5 | 7.5–11 | 0.5–4 | 4–7.5 | 7.5–11 | ||
| 2. Year | 3. Year | 4. Year | ||||||||
| UAST (%) | Competitors | 52.6 | 45.3 | 48.8 | 37.6 | 28.4 | 27.9 | 49.6 | 49 | 48.8 |
| Ruderals | 9.1 | 10.2 | 8 | 3.1 | 9.7 | 9.2 | 1.9 | 1.1 | 0.8 | |
| Competitors/Ruderals | 4.6 | 8.5 | 7.8 | 8.5 | 6 | 6.5 | 8.5 | 10.8 | 10.7 | |
| Competitors/Stress tolerant | 11.6 | 10.3 | 9 | 20 | 15.1 | 14.7 | 15.6 | 13.7 | 14.1 | |
| Competitors/Stress tolerant/Ruderals | 19.4 | 20.2 | 17.9 | 28.1 | 36.1 | 36.8 | 25.5 | 25.4 | 23.8 | |
| Growth form (%) | Geophyte | 27.9 | 16.4 | 21.3 | 17.7 | 14.74 | 14.6 | 22.7 | 21.8 | 18 |
| Therophyte | 11 | 15 | 12.5 | 4.3 | 13.2 | 12.8 | 0.5 | 0.5 | 0.7 | |
| Hemicryptophyte | 40.6 | 37.6 | 34.1 | 48.2 | 34.8 | 43.9 | 51 | 44.7 | 45.1 | |
| Phanerophyte | 19.2 | 25.2 | 23.6 | 27.6 | 35.6 | 36.9 | 25.8 | 32 | 34.4 | |
| Ellenberg’s value | Moisture | 7 | 6.1 | 6.5 | 6.2 | 6.6 | 6.6 | 6.8 | 7 | 7 |
| Nitrogen | 5.4 | 5 | 5.2 | 5.3 | 5.5 | 5.4 | 5.5 | 5.9 | 5.8 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuševica, A.; Celma, S.; Neimane, S.; von Cossel, M.; Lazdina, D. Wood-Ash Fertiliser and Distance from Drainage Ditch Affect the Succession and Biodiversity of Vascular Plant Species in Tree Plantings on Marginal Organic Soil. Agronomy 2022, 12, 421. https://doi.org/10.3390/agronomy12020421
Zuševica A, Celma S, Neimane S, von Cossel M, Lazdina D. Wood-Ash Fertiliser and Distance from Drainage Ditch Affect the Succession and Biodiversity of Vascular Plant Species in Tree Plantings on Marginal Organic Soil. Agronomy. 2022; 12(2):421. https://doi.org/10.3390/agronomy12020421
Chicago/Turabian StyleZuševica, Austra, Santa Celma, Santa Neimane, Moritz von Cossel, and Dagnija Lazdina. 2022. "Wood-Ash Fertiliser and Distance from Drainage Ditch Affect the Succession and Biodiversity of Vascular Plant Species in Tree Plantings on Marginal Organic Soil" Agronomy 12, no. 2: 421. https://doi.org/10.3390/agronomy12020421
APA StyleZuševica, A., Celma, S., Neimane, S., von Cossel, M., & Lazdina, D. (2022). Wood-Ash Fertiliser and Distance from Drainage Ditch Affect the Succession and Biodiversity of Vascular Plant Species in Tree Plantings on Marginal Organic Soil. Agronomy, 12(2), 421. https://doi.org/10.3390/agronomy12020421