
Protein Profiling of Pollen–Pistil Interactions in Almond (Prunus dulcis) and Identification of a Transcription Regulator Presumably Involved in Self-Incompatibility

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Controlled Pollinations
2.2. iTRAQ and 2D-nano-HPLC-ESI-MS/MS Analyses
2.2.1. Protein Extraction, Digestion, and Tagging with iTRAQ-4-plex® Reagent
2.2.2. Liquid Chromatography and Mass Spectrometer Analysis
2.2.3. Data Analysis of Mass Spectrometry
2.3. RNA Isolation and cDNA Synthesis
2.4. Full-Length cDNA Amplification and Sequence Analysis
2.5. RT-qPCR Analysis
3. Results
3.1. iTRAQ Analysis
3.2. Differentially Expressed Proteins in Unpollinated Pistils
3.3. Differentially Expressed Proteins after the Compatible Pollination
3.4. Differentially Expressed Proteins after the Incompatible Pollination
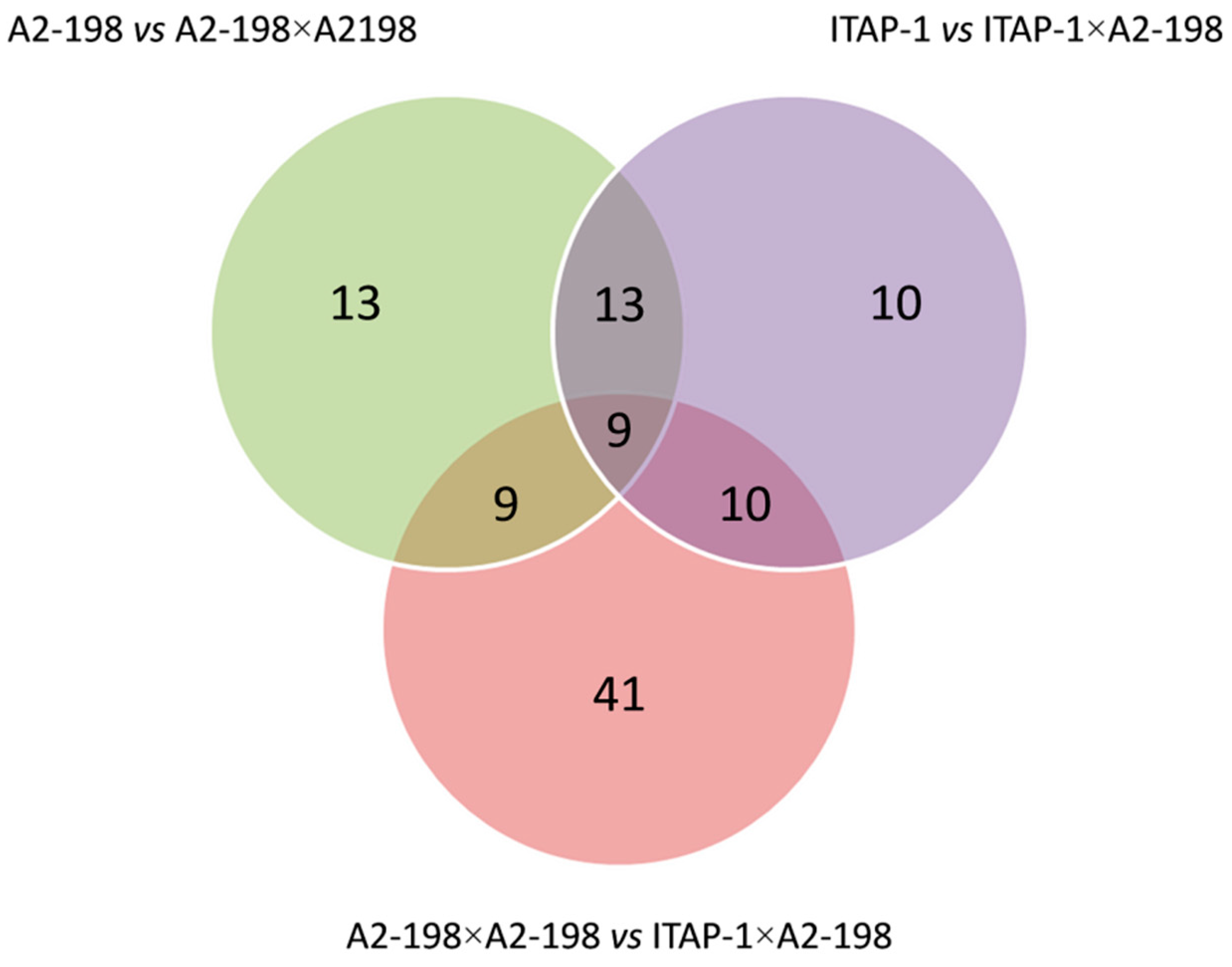
3.5. Differentially Expressed Proteins in Compatible and Incompatible Pollinations
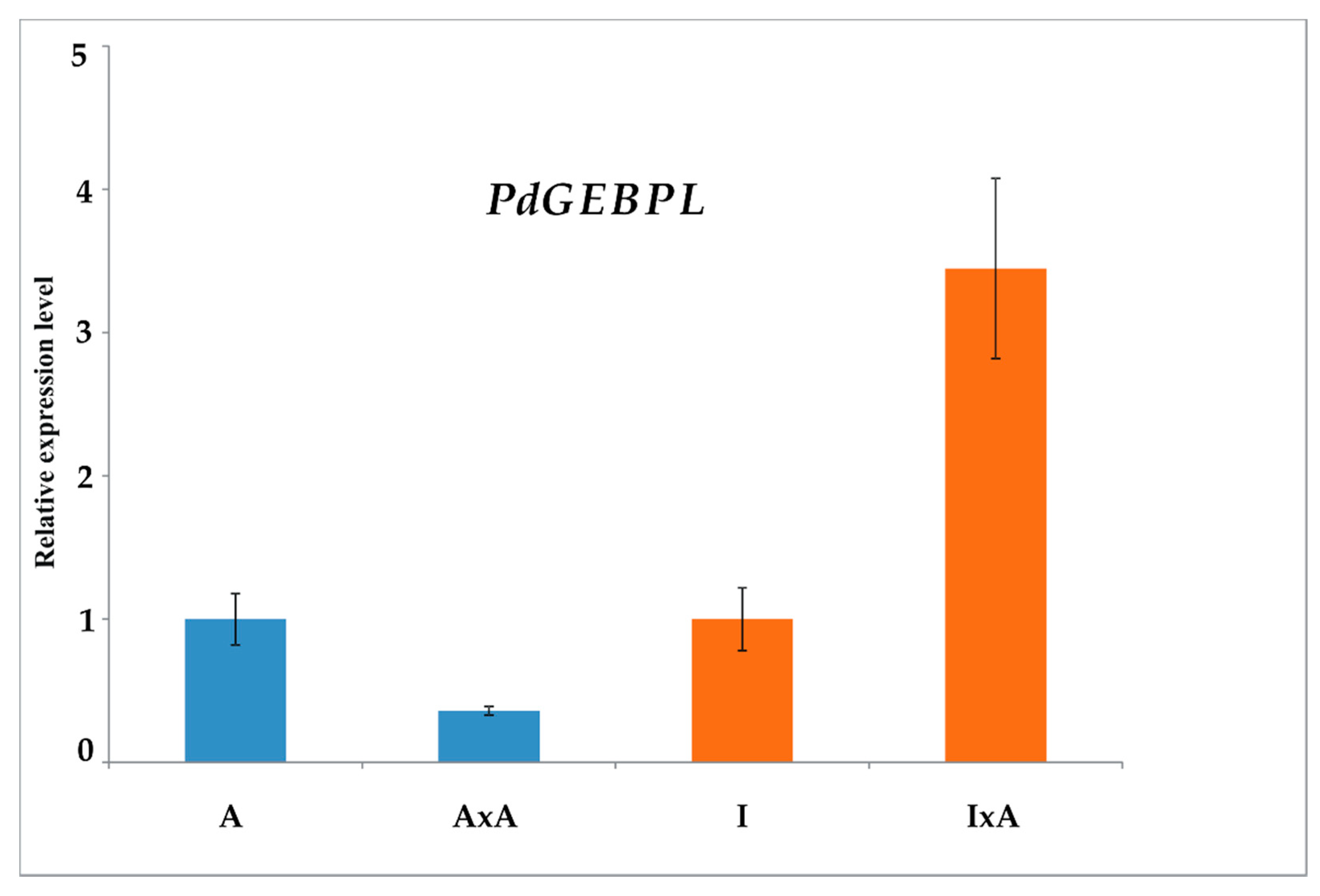
3.6. M5WQU9 Protein and Analysis of Its Expression by RT-qPCR
4. Discussion
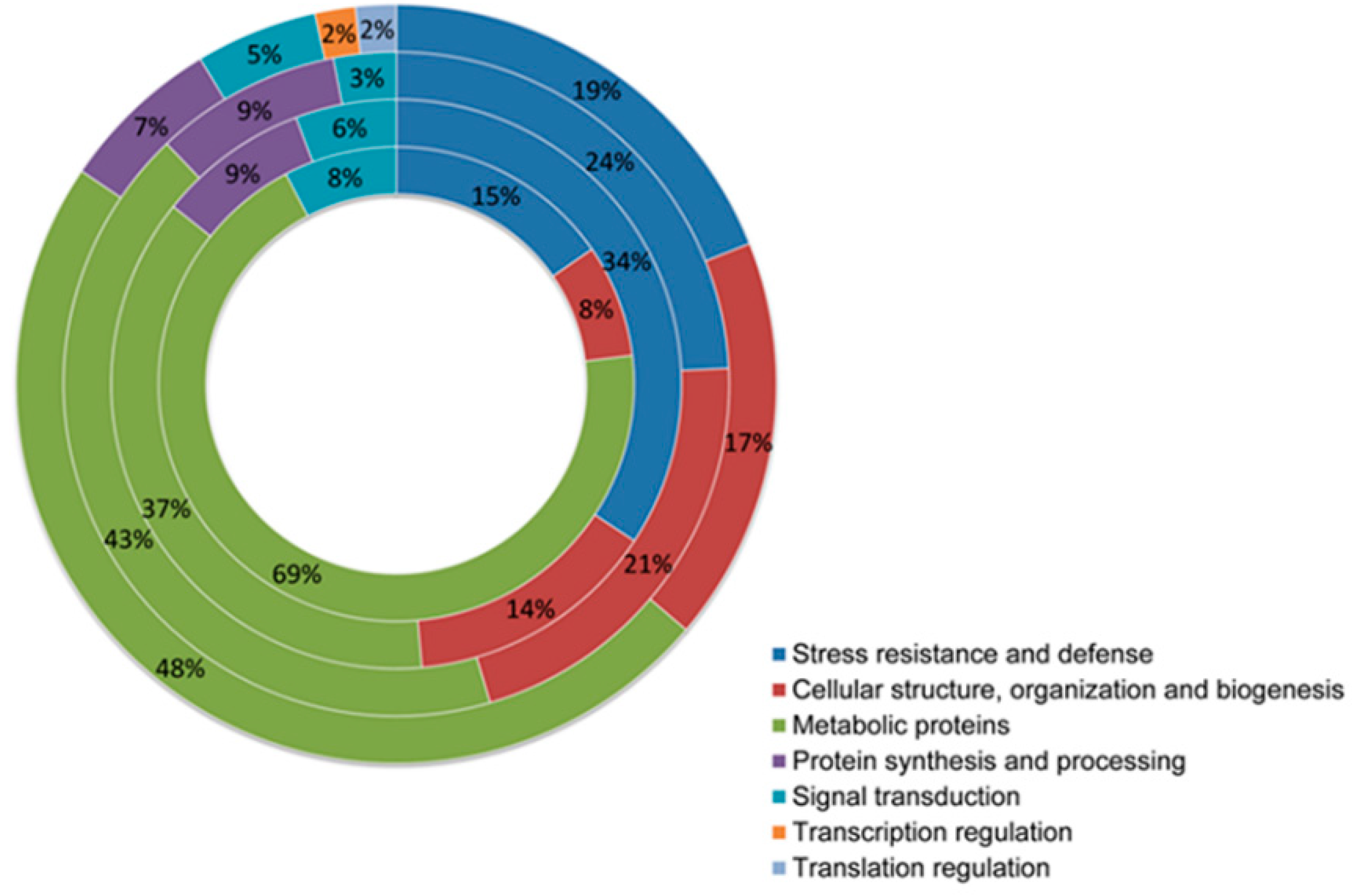
4.1. Classification of Differentially Expressed Proteins
4.2. Characterization of Differentially Expressed Proteins
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tao, R.; Iezzoni, A.F. The S-RNase-based gametophytic self-incompatibility system in Prunus exhibits distinct genetic and molecular features. Sci. Hortic. 2010, 124, 423–433. [Google Scholar] [CrossRef]
- Martí, A.F.i.; Howad, W.; Tao, R.; Segura, J.M.A.; Arús, P.; Socias i Company, R. Identification of quantitative trait loci associated with self-compatibility in a Prunus species. Tree Genet. Genomes 2011, 7, 629–639. [Google Scholar] [CrossRef]
- Bošković, R.I.; Tobutt, K.R.; Ortega, E.; Sutherland, B.G.; Godini, A. Self-(in)compatibility of the almonds P. dulcis and P. webbii: Detection and cloning of ‘wild-type Sf’ and new self-compatibility alleles encoding inactive S-RNases. Mol. Genet. Genomics 2007, 278, 665–676. [Google Scholar] [CrossRef] [PubMed]
- Godini, A. Ipotesi sulla comparsa dell’ autocompatibilitá nel mandorlo. Rivista di Scienza e Tecnica Agraria 1979, 19, 3–10. [Google Scholar]
- Kodad, O.; Alonso, J.M.; Fernández i Marti, A.; Oliveira, M.M.; Socias i Company, R. Molecular and physiological identification of new S-alleles associated with self-(in)compatibility in local Spanish almond cultivars. Sci. Hortic. 2010, 123, 308–311. [Google Scholar] [CrossRef]
- Martínez-García, P.J.; Mañas, F.; López, P.; Dicenta, F.; Ortega, E. Molecular and phenotypic characterization of the S-locus and determination of flowering time in new ‘Marcona’ and ‘Desmayo Largueta’-type almond (Prunus dulcis) selections. Euphytica 2011, 177, 67–78. [Google Scholar] [CrossRef]
- Gómez, E.M.; Buti, M.; Sargent, D.J.; Dicenta, F.; Ortega, E. Transcriptomic analysis of pollen-pistil interactions in almond (Prunus dulcis) identifies candidate genes for components of gametophytic self-incompatibility. Tree Genet. Genomes 2019, 15, 53. [Google Scholar] [CrossRef]
- Cao, X.; Feng, J.; Wang, D.; Sun, J.; Lu, X.; Liu, H. Primary style protein expression in the self-incompatible/self-compatible apricot by the 2D-DIGE technique. Gene 2012, 503, 110–117. [Google Scholar] [CrossRef]
- Feng, J.; Chen, X.; Yuan, Z.; He, T.; Zhang, L.; Wu, Y.; Liu, W.; Liang, Q. Proteome comparison following self- and across-pollination in self-incompatible apricot (Prunus armeniaca L.). Protein J. 2006, 25, 328–335. [Google Scholar] [CrossRef]
- Feng, J.R.; Chen, X.S.; Yuan, Z.H.; Zhang, L.J.; Ci, Z.J.; Liu, X.L.; Zhang, C.Y. Primary molecular features of self-incompatible and self-compatible F1 seedling from apricot (Prunus armeniaca L.) Katy × Xinshiji. Mol. Biol. Rep. 2009, 36, 263–272. [Google Scholar] [CrossRef]
- Gómez, E.M.; Dicenta, F.; Martínez-García, P.J.; Ortega, E. iTRAQ-based quantitative proteomic analysis of pistils and anthers from self-incompatible and self-compatible almonds with the Sf haplotype. Mol. Breed. 2015, 35, 120. [Google Scholar] [CrossRef]
- Lénárt, J.; Gere, A.; Causon, T.; Hann, S.; Dernovics, M.; Németh, O.; Hegedűs, A.; Halász, J. LC–MS based metabolic fingerprinting of apricot pistils after self-compatible and self-incompatible pollinations. Plant Mol. Biol. 2021, 105, 435–447. [Google Scholar] [CrossRef] [PubMed]
- Martínez-García, P.J.; Gómez, E.M.; Casado-Vela, J.; Elortza, F.; Dicenta, F.; Ortega, E. Differential protein expression in compatible and incompatible pollen-pistil interactions in almond [Prunus dulcis (Miller) D.A. Webb] by 2D-DIGE and HPLC-MS/MS. J. Hort. Sci. Biotech. 2015, 90, 71–77. [Google Scholar] [CrossRef]
- Dresselhaus, T.; Franklin-Tong, N. Male-female crosstalk during pollen germination, tube growth and guidance, and double fertilization. Mol. Plant 2013, 6, 1018–1036. [Google Scholar] [CrossRef]
- Kim, M.-J.; Jeon, B.W.; Oh, E.; Seo, P.J.; Kim, J. Peptide signaling during plant reproduction. Trends Plant Sci. 2021, 26, 822–835. [Google Scholar] [CrossRef]
- Ross, P.L.; Huang, Y.L.N.; Marchese, J.N.; Williamson, B.; Parker, K.; Hattan, S.; Khainovski, N.; Pillai, S.; Dey, S.; Daniels, S.; et al. Multiplexed protein quantitation in Saccharomyces cerevisiae using amine-reactive isobaric tagging reagents. Mol. Cell. Proteom. 2004, 3, 1154–1169. [Google Scholar] [CrossRef]
- Evans, C.; Noirel, J.; Ow, S.Y.; Salim, M.; Pereira-Medrano, A.G.; Couto, N.; Pandhal, J.; Smith, D.; Pham, T.K.; Karunakaran, E.; et al. An insight into iTRAQ: Where do we stand now? Anal. Bioanal. Chem. 2012, 404, 1011–1027. [Google Scholar] [CrossRef]
- Nägele, E.; Vollmer, M.; Hörth, P. Two-dimensional nano-liquid chromatography-mass spectrometry system for applications in proteomics. J. Chromatogr. A 2003, 1009, 197–205. [Google Scholar] [CrossRef]
- Felipe, A.J. Stadi fenologici del mandorlo. In Proceedings of the 3rd GREMPA Colloquium, Valenzano, Bari, Italy, 3–7 October 1977; pp. 60–61. [Google Scholar]
- Ortega, E.; Dicenta, F. Inheritance of self-compatibility in almond: Breeding strategies to assure self-compatibility in the progeny. Theor. Appl. Genet. 2003, 106, 904–911. [Google Scholar] [CrossRef]
- Gómez, E.M. Factores Moleculares Implicados En El Sistema De Incompatibilidad Floral En Almendro [Prunus dulcis (Miller) D.A. Webb]. Ph.D. Thesis, University of Murcia, Murcia, Spain, 2017. [Google Scholar]
- Remy, P. Contribution a l’étude du pollen des arbres fruitiers a noyau, genre Prunus. Ann. Amélior. Plantes 1953, 3, 351–388. [Google Scholar]
- Ortega, E.; Egea, J.; Cánovas, J.A.; Dicenta, F. Pollen tube dynamics following half- and fully-compatible pollinations in self-compatible almond cultivars. Sex. Plant Reprod. 2002, 15, 47–51. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Méndez, L.; Ciordia, S.; Fernández, M.S.; Juárez, S.; Ramos, A.; Pazos, M.; Gallardo, J.M.; Torres, J.L.; Nogués, M.R.; Medina, I. Changes in liver proteins of rats fed standard and high-fat and sucrose diets induced by fish omega-3 PUFAs and their combination with grape polyphenols according to quantitative proteomics. J. Nutr. Biochem. 2017, 41, 84–97. [Google Scholar] [CrossRef] [PubMed]
- Verde, I.; Abbott, A.G.; Scalabrin, S.; Jung, S.; Shu, S.; Marroni, F.; Zhebentyayeva, T.; Dettori, M.T.; Grimwood, J.; Cattonaro, F.; et al. The high-quality draft genome of peach (Prunus persica) identifies unique patterns of genetic diversity, domestication and genome evolution. Nat. Genet. 2013, 45, 487–494. [Google Scholar] [CrossRef]
- Cachi, A.M.; Wunsch, A. Characterization and mapping of non-S gametophytic self-compatibility in sweet cherry (Prunus avium L.). J. Exp. Bot. 2011, 62, 1847–1856. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Sanz, J.V.; Zuriaga, E.; López, I.; Badenes, M.L.; Romero, C. Self-(in)compatibility in apricot germplasm is controlled by two major loci, S and M. BMC Plant Biol. 2017, 17, 82. [Google Scholar] [CrossRef]
- Gambino, G.; Perrone, I.; Gribaudo, I. A rapid and effective method for RNA extraction from different tissues of grapevine and other woody plants. Phytochem. Anal. 2008, 19, 520–525. [Google Scholar] [CrossRef]
- Undergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S. G. Primer3-new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef]
- Tong, Z.; Gao, Z.; Wang, F.; Zhou, J.; Zhang, Z. Selection of reliable reference genes for gene expression studies in peach using real-time PCR. BMC Mol. Biol. 2009, 10, 71. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, 2002–2007. [Google Scholar] [CrossRef]
- Chalivendra, S.C.; López-Casado, G.; Kumar, A.; Kassenbrock, A.R.; Royer, S.; Tovar-Mèndez, A.; Covey, P.A.; Dempsey, L.A.; Randle, A.M.; Stack, S.M.; et al. Developmental onset of reproductive barriers and associated proteome changes in stigma/styles of Solanum pennellii. J. Exp. Bot. 2013, 64, 265–279. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.H.; Chen, G.L.; Huang, J. Shot-gun proteome and transcriptome mapping of the jujube floral organ and identification of a pollen-specific S-locus F-box gene. Peer J. 2017, 5, e3588. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Wang, M.; Zhao, L. Dissecting stylar responses to self-pollination in wild tomato self-compatible and self-incompatible species using comparative proteomics. Plant Physiol. Biochem. 2016, 106, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Marshall, E.; Costa, L.M.; Gutiérrez-Marcos, J. Cysteine-rich peptides (CRPs) mediate diverse aspects of cell-cell communication in plant reproduction and development. J. Exp. Bot. 2011, 62, 1677–1686. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wang, K.; Li, S.; Yang, P. Exploration of rice pistil responses during early post-pollination through a combined proteomic and transcriptomic analysis. J. Proteom. 2016, 131, 214–226. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, A.; Li, W.; Jiang, Y.; Song, S.; Li, Y.; Chen, R. Comparative proteomic analysis of eggplant (Solanum melongena L.) heterostylous pistil development. PLoS ONE 2017, 12, e0179018. [Google Scholar] [CrossRef]
- Samuel, M.A.; Tang, W.Q.; Jamshed, M.; Northey, J.; Patel, D.; Smith, D.; Siu, K.W.M.; Muench, D.G.; Wang, Z.Y.; Goring, D.R. Proteomic analysis of Brassica stigmatic proteins following the self-incompatibility reaction reveals a role for microtubule dynamics during pollen responses. Mol. Cell Proteom. 2011, 10, M111.011338. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.Q.; Ma, R.C. Generation and analysis of expressed sequence tags from almond (Prunus dulcis Mill.) pistils. Sex. Plant Reprod. 2003, 16, 197–207. [Google Scholar] [CrossRef]
- Sassa, H.; Hirano, H. Identification of a new class of pistil-specific proteins of Petunia inflata that is structurally similar to, but functionally distinct from, the self-incompatibility factor HT. Mol. Genet. Genom. 2006, 275, 97–104. [Google Scholar] [CrossRef]
- Timperio, A.M.; Egidi, M.G.; Zolla, L. Proteomics applied on plant abiotic stresses: Role of heat shock proteins (HSP). J. Protem. 2008, 71, 391–411. [Google Scholar] [CrossRef]
- Sassa, H.; Hirano, H. Style-specific and developmentally regulated accumulation of a glycosylated thaumatin/PR5-like protein in Japanese pear (Pyrus serotina Rehd. ) Planta 1998, 205, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Kuboyama, T. A novel thaumatin-like protein gene of tobacco is specifically expressed in the transmitting tissue of stigma and style. Sex. Plant Reprod. 1998, 11, 251–256. [Google Scholar] [CrossRef]
- Grenier, J.; Potvin, C.; Trudel, J.; Asselin, A. Some thaumatin-like proteins hydrolyse polymeric b-1,3-glucans. Plant J. 1999, 19, 473–480. [Google Scholar] [CrossRef]
- Nishikawa, S.-i.; Zinkl, G.M.; Swanson, R.J.; Maruyama, D.; Preuss, D. Callose (β-1,3 glucan) is essential for Arabidopsis pollen wall patterning, but not tube growth. BMC Plant Biol. 2005, 5, 22. [Google Scholar] [CrossRef] [PubMed]
- Afzal, A.J.; Wood, A.J.; Lightfoot, D.A. Plant receptor-like serine threonine kinases: Roles in signaling and plant defense. Mol. Plant Microbe Interact 2008, 21, 507–517. [Google Scholar] [CrossRef]
- Losada, J.M.; Herrero, M. Glycoprotein composition along the pistil of Malus x domestica and the modulation of pollen tube growth. BMC Plant Biol. 2014, 14, 1. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, K.; Yin, G.; Liu, X.; Liu, M.; Cao, N.; Duan, Y.; Gao, H.; Wang, W.; Ge, W.; et al. Pollen-expressed Leucine-Rich Repeat extensins are essential for pollen germination and growth. Plant Physiol. 2018, 176, 1993–2006. [Google Scholar] [CrossRef] [PubMed]
- Menezes, S.P.; Silva, E.M.d.A.; Lima, E.M.; Sousa, A.O.d.; Andrade, B.S.; Lemos, L.S.L.; Gramacho, K.P.; Gesteira, A.d.S.; Pirovani, C.P.; Micheli, F. The pathogenesis-related protein PR-4b from Theobroma cacao presents RNase activity, Ca2+ and Mg2+ dependent-DNase activity and antifungal action on Moniliophthora perniciosa. BMC Plant Biol. 2014, 14, 161. [Google Scholar]
- Ori, N.; Sessa, G.; Lotan, T.; Himmelhoch, S.; Fluhr, R. A major stylar matrix polypetide (sp41) is a member of the pathogenesis-related proteins superclass. EMBO J. 1990, 9, 3429–3436. [Google Scholar] [CrossRef]
- Constabel, C.P.; Brisson, N. Stigma and vascular-specific expression of the PR-10a gene of potato: A novel pattern of expression of a pathogenesis-related gene. Mol. Plant-Microbe Interact 1995, 8, 104–113. [Google Scholar] [CrossRef]
- Eldik, G.J.V.; Wingens, M.; Ruiter, R.K.; Herpen, M.M.V.; Schrauwen, J.A.; Wullems, G.J. Molecular analysis of a pistil-specific gene expressed in the stigma and stylar cortex of Solanum tuberosum. Plant Mol. Biol. 1996, 30, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Rose, J.K.C.; Braam, J.; Fry, S.C.; Nishitani, K. The XTH family of enzymes involved in xyloglucan endotransglucosylation and endohydrolysis: Current perspectives and a new unifying nomenclature. Plant Cell Physiol. 2002, 43, 1421–1435. [Google Scholar] [CrossRef] [PubMed]
- Mollet, J.C.; Leroux, C.; Dardelle, F.; Lehner, A. Cell wall composition, biosynthesis and remodeling during pollen tube growth. Plants 2013, 2, 107–147. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhang, Q.; Zhao, K.; Luo, Q.; Bao, S.; Liu, H.; Men, S. The Arabidopsis GPR1 gene negatively affects pollen germination, pollen tube growth, and gametophyte senescence. Int. J. Mol. Sci. 2017, 18, 1303. [Google Scholar] [CrossRef]
- Curaba, J.; Herzog, M.; Vachon, G. GeBP, the first member of a new gene family in Arabidopsis, encodes a nuclear protein with DNA-binding activity and is regulated by KNAT1. Plant J. 2003, 33, 305–317. [Google Scholar] [CrossRef]
- Chevalier, F.; Perazza, D.; Laporte, F.; Le Hénanf, G.; Hornitschek, P.; Bonneville, J.M.; Herzog, M.; Vachon, G. GeBP and GeBP-like proteins are noncanonical Leucine-Zipper transcription factors that regulate cytokinin response in Arabidopsis. Plant Physiol. 2008, 146, 1142–1154. [Google Scholar] [CrossRef][Green Version]
- Kovaleva, L.; Zakharova, E. Hormonal status of the pollen-pistil system at the progamic phase of fertilization after compatible and incompatible pollination in Petunia hybrida L. Sex Plant Reprod. 2003, 16, 191–196. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Comparison a | DEPs | Up-Regulated b | Down-Regulated c | Unidentified |
|---|---|---|---|---|
| A vs. I | 13 | 6 | 7 | - |
| A × A vs. A | 44 | 37 | 7 | 9 |
| I × A vs. I | 42 | 36 | 6 | 9 |
| A × A vs. I × A | 69 | 26 | 43 | 11 |
| A vs. I a | A × A vs. A b | I × A vs. I c | A × A vs. I × A d | Description e | Function | GDR Gen f | Scaffold g |
|---|---|---|---|---|---|---|---|
| Stress resistance and defense | |||||||
| M5VM54 | Putative LRR receptor-like serine/threonine-protein kinase | Pathogenesis process | ppa007248m | 8 | |||
| M5W1Q5 | M5W1Q5 | M5W1Q5 | M5W1Q5 | Pathogenesis-related protein PR-4 | Pathogenesis process | ppa012991m | 6 |
| M5WR86 | M5WR86 | Pathogenesis-related thaumatin-like protein | Pathogenesis process | ppa019480m | 5 | ||
| M5WT96 | M5WT96 | M5WT96 | Pathogenesis-related thaumatin-like protein | Pathogenesis process | ppa024543m | 5 | |
| Cellular structure, organization and biogenesis | |||||||
| M5WH02 | M5WH02 | Xyloglucan endotransglucosylase | Hydrolase activity | ppa009685m | 4 | ||
| M5WJ79 | M5WJ79 | Xyloglucan endotransglucosylase | Hydrolase activity | ppa009472m | 6 | ||
| M5WT30 | M5WT30 | Xyloglucan endotransglucosylase | Hydrolase activity | ppa009608m | 3 | ||
| Signal transduction | |||||||
| M5XF62 | M5XF62 | GTP-binding nuclear protein | GTP binding | ppa007452m | 1 | ||
| Transcription regulation | |||||||
| M5WQU9 | GLABROUS1 enhancer-binding protein-like * | Transcription regulation | ppa006690m | 3 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez, E.M.; Prudencio, Á.S.; Ortega, E. Protein Profiling of Pollen–Pistil Interactions in Almond (Prunus dulcis) and Identification of a Transcription Regulator Presumably Involved in Self-Incompatibility. Agronomy 2022, 12, 345. https://doi.org/10.3390/agronomy12020345
Gómez EM, Prudencio ÁS, Ortega E. Protein Profiling of Pollen–Pistil Interactions in Almond (Prunus dulcis) and Identification of a Transcription Regulator Presumably Involved in Self-Incompatibility. Agronomy. 2022; 12(2):345. https://doi.org/10.3390/agronomy12020345
Chicago/Turabian StyleGómez, Eva M., Ángela S. Prudencio, and Encarnación Ortega. 2022. "Protein Profiling of Pollen–Pistil Interactions in Almond (Prunus dulcis) and Identification of a Transcription Regulator Presumably Involved in Self-Incompatibility" Agronomy 12, no. 2: 345. https://doi.org/10.3390/agronomy12020345
APA StyleGómez, E. M., Prudencio, Á. S., & Ortega, E. (2022). Protein Profiling of Pollen–Pistil Interactions in Almond (Prunus dulcis) and Identification of a Transcription Regulator Presumably Involved in Self-Incompatibility. Agronomy, 12(2), 345. https://doi.org/10.3390/agronomy12020345