4.1. In Vitro Mycelial Growth Inhibition
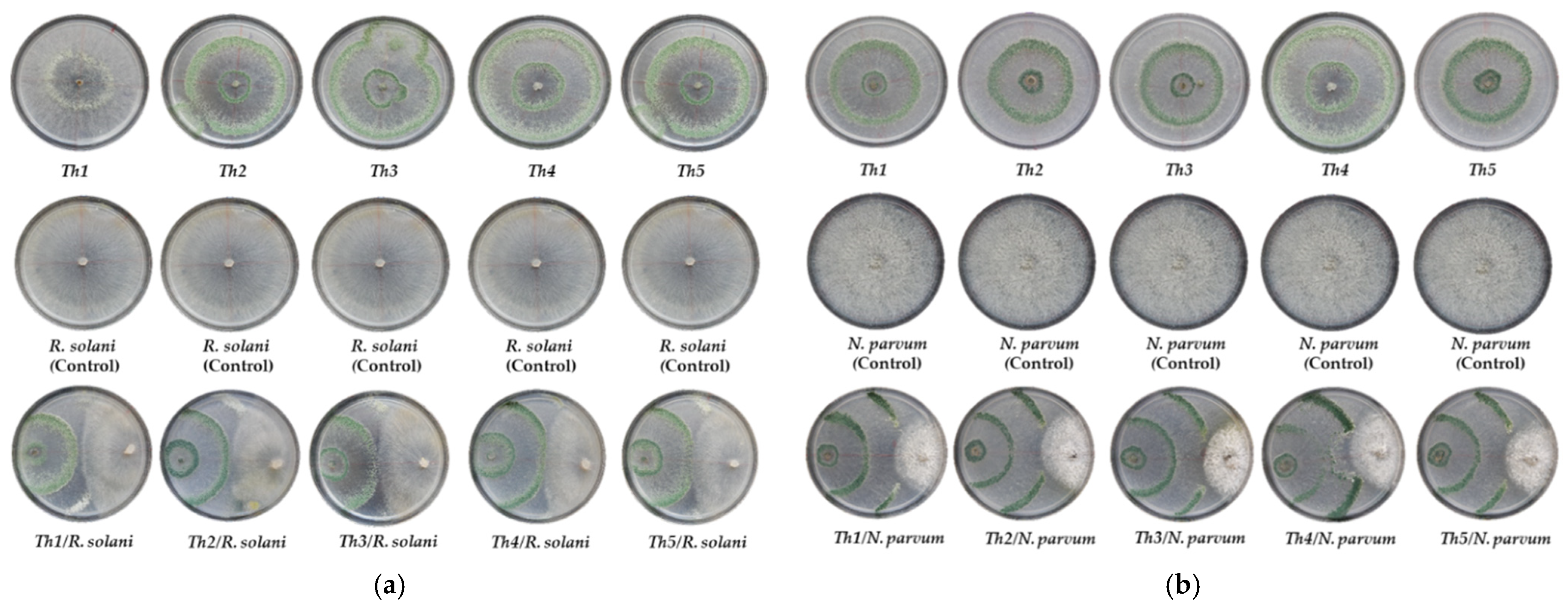
Plate confrontation assays demonstrated the ability of
T. harzianum strains to inhibit the mycelial growth of the two selected grapevine pathogens. In general terms, the highest inhibition values were recorded against
N. parvum, representing almost twice those recorded against
R. solani. Numerous studies have evaluated in vitro the antagonistic potential of several
Trichoderma species, a well-known group of hyperparasitic mitosporic fungi [
19,
20]. In this sense, the values obtained in the present study seem to be relatively discrete in comparison with PIRG data obtained from the confrontations of species of the mentioned genus against other plant pathogens [
21,
22,
23], in which the average rates of inhibition of microbial growth in dual confrontations with
T. harzianum were usually higher than 50%, sometimes reaching PIRG values above 99%.
One possible explanation is that both
N. parvum and
R. solani species have simple nutritional requirements and usually show high-speed growth rates. This is especially true for isolates of the genus
Rhizoctonia, a polyphagous basidiomycete with great colonizing capacity and very high vegetative growth rates (with practically absent reproductive mechanisms) [
24]. This would hinder its inhibition by other antagonists, especially by those that, like
Trichoderma, base their mode of action mainly on the colonization and parasitism of the mycelium of the pathogen to be controlled, always accompanied by an effective occupation of the culture medium. Thus, in the case of
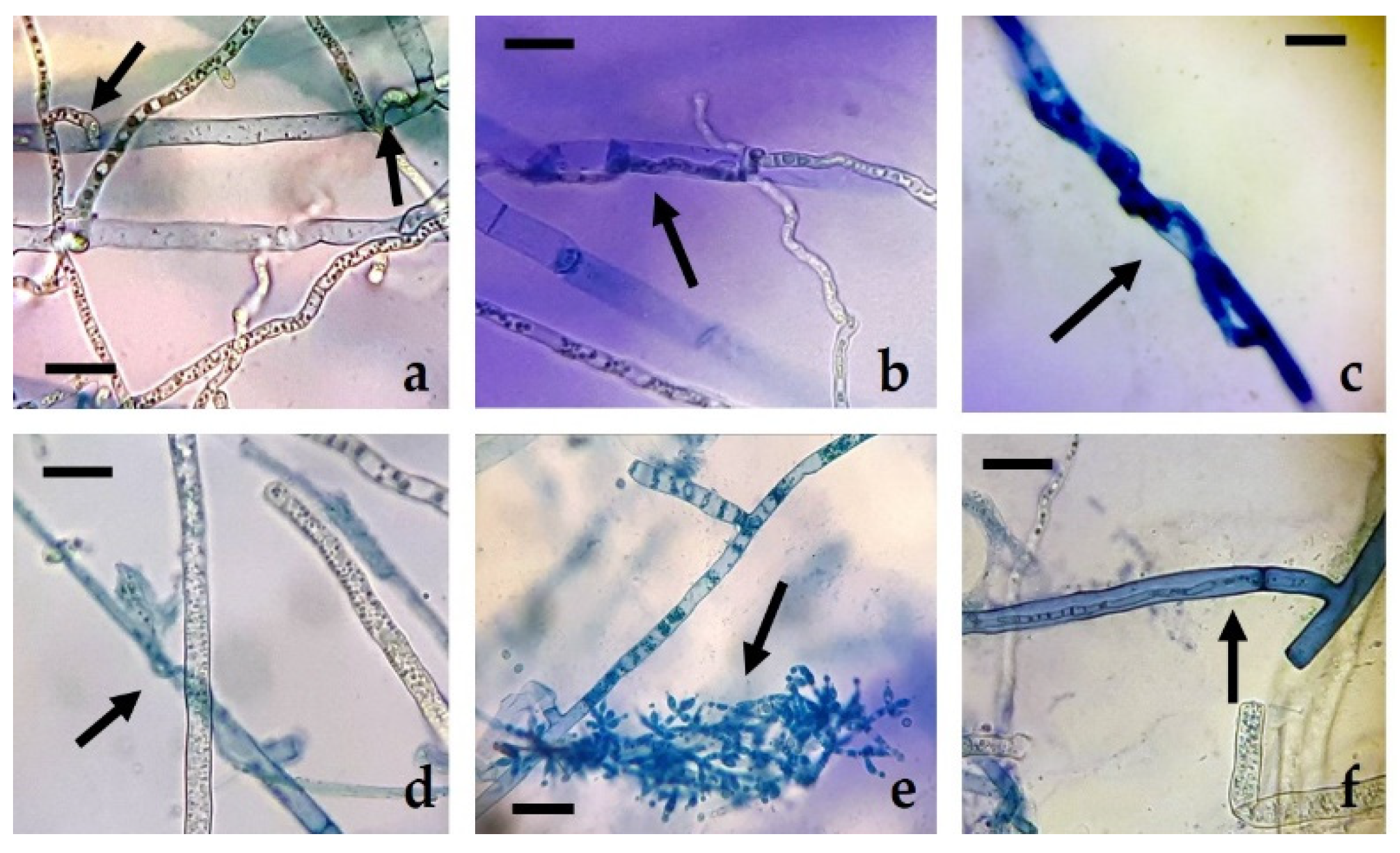
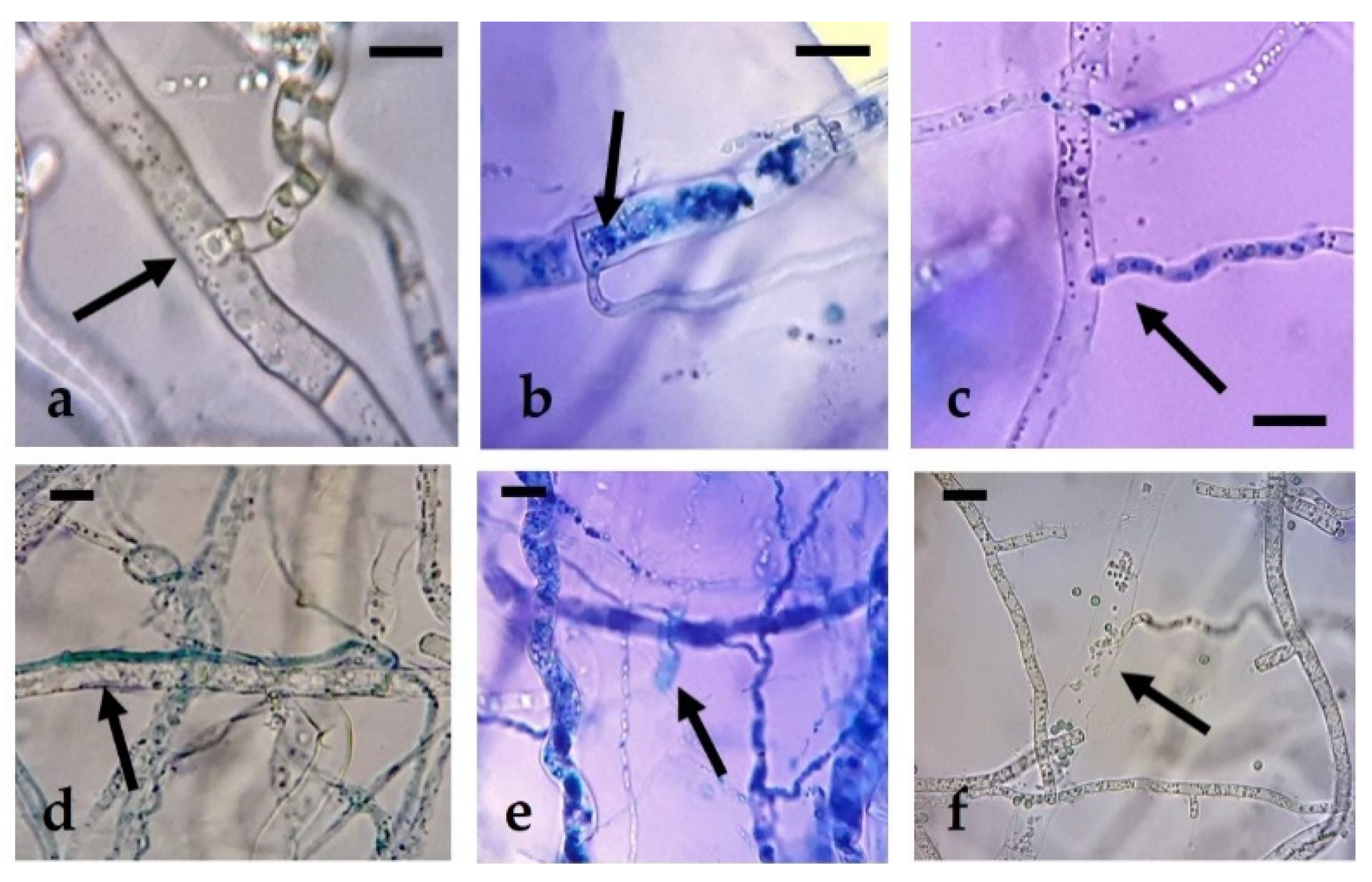
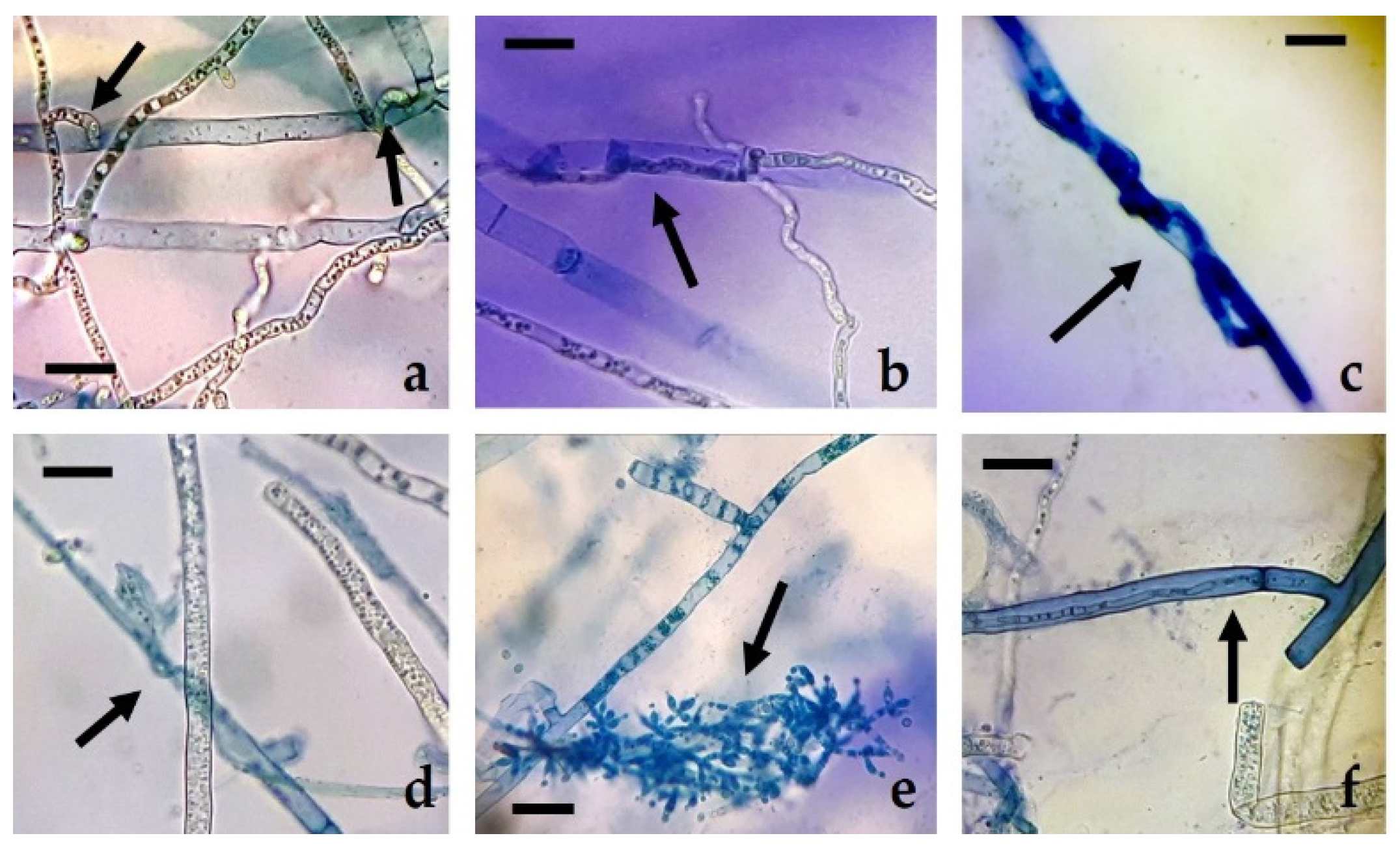
R. solani, the presence of hyperparasitism phenomena (enzymatic lysis of the mycelium of
R. solani) and sporulation of the different isolates of
Trichoderma on the mycelium of the pathogen can be observed, but not beyond half of the Petri dish (
Figure 4), due to the rapid growth and colonization capacity of
R. solani, with rates similar to those of
Trichoderma. Siameto et al. [
25], in a study on the growth inhibition of selected soil pathogens in Africa, found that
T. harzianum inhibited the growth of
R. solani with the highest PIRG being 61.55%, while the lowest was 25.88%; and, according to Guedez et al. [
26], in a study on the in vitro growth-inhibitory activity of different strains of
T. harzianum against
R. solani and two other tomato pathogens, rates between 62 and 72% were recorded for inhibition against this fungus, well above those obtained here. However, the aggressiveness, virulence, or speed of colonization of isolates of a species as biologically complex as
R. solani largely depends on the type of anastomosis group and/or subgroup in question [
24,
26].
In the case of
N. parvum, as indicated above, PIRG values were shown to be clearly higher (
Figure 4). These values seem to be in line with the mycelial growth inhibition rates observed in certain species of the grapevine pathogenic
Botrysphaeriaceae family in plate confrontations with different isolates of the genus
Trichoderma. Marraschi et al. [
27] evaluated the potential of different BCAs and determined that the tested
Trichoderma species and commercial formulations could inhibit the growth of a member of the mentioned ascomycetous family such as
Lasiodiplodia theobromae (Pat.) Griffon & Maubl. at rates above 75%. Urbez-Torres et al. [
28] studied different isolates of
Trichoderma spp. that inhibited the growth of several species of
Botryosphaeriaceae in a range from 44.5 to 74.3%, with
T. atroviride being the most effective, close to
T. harzianum. Plata-Caudillo [
29] obtained mycelial growth inhibition values of
N. parvum around 50% in direct plate confrontations with several isolates and formulations based on
T. harzianum, in line with the values obtained in the present work. Mutawila et al. [
30] observed mycelial growth inhibition rates against
N. parvum of 45–50%, although using extracts containing a metabolite of interest previously extracted from different
Trichoderma strains. Finally, Kotze et al. [
31], in an in vitro and in vivo evaluation of the antagonistic activity of different microorganisms and formulations, reported, in the case of plate confrontations with
T. harzianum versus
N. parvum, the existence of hyperparasitism of the former towards the latter, based on the presence of hyphal interactions, enzymatic lysis, and occupation of space and nutrients in Petri dish. These hyphal interactions were also observed in the confrontations of the present study, although they were not particularly abundant along the whole contact surface between the colonies.
4.2. Bioassays of Trichoderma harzianum against Neofusicoccum parvum in Plants
Neofusicoccum parvum has been repeatedly isolated from nurseries throughout the world’s wine-growing areas in the last two decades and has been the subject of numerous investigations related to its epidemiology and characterization, as well as its control, including biological methods [
12,
32,
33]. Most of the approaches related to the biocontrol of
N. parvum and related
Botryosphaeriaceae species have been based either on the protection of pruning wounds with antagonist-based formulations or other alternative methods to chemical fungicides [
31,
34,
35], or with the application of these biocontrol agents in the different stages of production of grafted plants in nurseries [
9,
12,
36].
In this sense, most of the investigations on the control of this and other pathologies of young grapevine plants, have been focused on grafted plants in the nursery and/or adult plants in the field, so the methodology and biometric results obtained after the treatment of grapevine plants germinated from seeds with different isolates of T. harzianum in the present study represents a novelty in this type of research. Nevertheless, the biometric data did not provide conclusive results regarding the protective effect of the different antagonistic strains used, given that a disparate behavior was observed among isolates, type of application, or measured parameter. Nevertheless, the reported data indicate, in agreement with the majority of studies based on the application of microbial antagonists at the root level, that the parameters most related to the development and activity of this type of tissue (root length and total weight) were the ones that offered better values compared to control plants inoculated with the pathogen.
Regarding the protection tests on grafted plants, successive treatments showed good root biomass lengths and lower necrosis (compared to controls). Several researchers claim that successive treatments with antagonists reduce the incidence of
N. parvum. In this sense, Pintos et al. [
37] concluded that inoculating
Trichoderma in plants three days before the pathogen reduced more than twice the length of necrosis caused by the latter, being more effective than inoculating three days after the pathogen. Similarly, Kotze et al. [
31] demonstrated a reduction of
N. parvum damage in pruning wounds by applying
Trichoderma products seven days before exposure, when the plant was healthy.
4.3. Bioassays of Trichoderma harzianum against Rhizoctonia solani in Plants
Members of the so-called
Rhizoctonia species complex are considered a mixture of filamentous fungi, having in common the possession of a non-spored imperfect state, usually referred to as the
Rhizoctonia anamorph [
5,
6,
38,
39,
40]. Among them,
Rhizoctonia solani is considered a very destructive plant pathogen, having a broad host range and causing diseases in a great variety of crops. Although
R. solani has long been known as one of the main pathogens associated with grapevine wood in young plants in greenhouses [
5,
6,
38,
39,
40], to date, there have been few studies on the biological protection of grapevine plants with microbial antagonists or other alternative methods against it. Crous et al. [
39] examined the effects of thermotherapy treatments on the prevalence and incidence of a number of pathogens associated with apparently healthy grapevine seedlings in nurseries, including
R. solani, concluding that hot water treatments did not significantly reduce its presence and the presence of other associated fungal pathogens. On the other hand, Ziedan et al. [
41] tested the efficacy of a series of bacterial and fungal antagonistic strains such as
T. harzianum against infection by
Fusarium oxysporum E.F.Sm. & Swingle and
R. solani in grapevine plants of the variety ‘Thompson Seedless’, concluding that both the application of these antagonists by immersion of the root system of the plants and the incorporation of the same to the culture substrate managed to reduce the incidence of root rot or the colonization of the roots by the pathogens. The results obtained in the present study on seedlings from germinated seed, although they cannot be compared with those reported in the studies mentioned above based on grafted plants, seem to agree with those previously cited on the fact that the protective effects of the antagonistic strains of
T. harzianum are basically associated to the root system of the treated plants, where successive treatments with the different strains of
T. harzianum reflected higher root development associated to lower colonization rates by the pathogen, compared to control plants artificially infected only with
R. solani.
Regarding biological control trials on grafted plants, it was shown that simultaneous applications of certain strains of
T. harzianum obtained higher values of root and aerial biomass length than controls with
R. solani. This is consistent with the findings of authors such as Marais [
38], Walker [
42], and Hemida et al. [
43], who stated that
R. solani infections in young grapevine plants are associated with a reduction in root biomass and the existence of active root rot, in addition to general retardation of plant growth.
4.4. Trichoderma harzianum as a Growth Promoter
Along with the protective effects against specific plant pathogens, another important aspect related to biocontrol trials with
Trichoderma is the ability of the different species of this genus to promote and stimulate plant growth, widely studied in different plant species [
19], including grapevine [
44,
45,
46,
47].
In the present study, the inoculation of Trichoderma protective strains was carried out at two levels: (1) on grapevine seedlings germinated from seed and (2) on grafted plants coming from the protection experiments against N. parvum and R. solani.
In the case of seedlings, the results obtained suggested that the different strains of T. harzianum were not able to increase the different biometric parameters analyzed compared to control plants without any microbial inoculant, except for T. harzianum 5, which promoted root development. A possible explanation for this could be the scarce development of the root systems of this type of plant, where the culture system based on a small container with inert substrate could be a limitation for the incorporation and colonization of the plant by the mentioned isolates of T. harzianum.
Regarding
T. harzianum growth promotion assays on grafted grapevine plants, biometric increases were recorded in plants from
T. harzianum control experiments with the two mentioned pathogens. This increase was especially significant in root biomass, regardless of the statistical significance of this type of interaction. This type of beneficial effects, especially at the root system level, has been previously reported for this antagonistic genus in numerous plant hosts, including grapevine, where some authors (e.g., Di Marco and Osti [
44]) observed significant increases in root biomass in grapevine plants treated with
Trichoderma, especially at the time of rooting, suggesting another relevant feature of this genus from the biotechnological point of view: its potential use as a microbial fertilizer [
48,
49]. In this sense, in the study of Di Marco et al. [
12], it was observed that grapevine roots developed four times more compared to controls when treated with
Trichoderma. The mentioned authors concluded that the grapevines treated with the mentioned genus presented a more developed root system, which improved the absorption of water and nutrients and endowed them with a greater tolerance to stress-related diseases. Similarly, Fourie et al. [
47] observed that the early shoot growth of
Trichoderma treated grapevine plants was visibly better than that of untreated control plants. Regarding the effect of
Trichoderma on rooting, these authors found that the fresh root weight of grapevine increased by 41.7% after monthly treatments with
Trichoderma in the greenhouse soil.


 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}