
Effects of Phenological Changes on Plant Production—From the View of Stipa krylovii

1
State Key Laboratory of Severe Weather, Hebei Gucheng Agricultural Meteorology National Observation and Research Station, Chinese Academy of Meteorological Sciences, Beijing 100081, China
2
Collaborative Innovation Center on Forecast and Evaluation of Meteorological Disasters (CIC-FEMD), Nanjing University of Information Science & Technology, Nanjing 210044, China
3
Joint Eco-Meteorological Laboratory, Chinese Academy of Meteorological Sciences and Zhengzhou University, Zhengzhou 450001, China
4
College of Resources and Environmental Sciences, China Agricultural University, Beijing 100193, China
*
Authors to whom correspondence should be addressed.
Agronomy 2022, 12(12), 3208; https://doi.org/10.3390/agronomy12123208
Submission received: 25 October 2022
/
Revised: 13 December 2022
/
Accepted: 14 December 2022
/
Published: 16 December 2022
(This article belongs to the Section Grassland and Pasture Science)
Abstract
:Global warming has changed plant phenology and induced variations in the productivity of terrestrial ecosystems. Recent studies have shown inconsistent results regarding the influence of phenological changes on plant production. We carried out a three-year in situ experiment in Inner Mongolia and used Stipa krylovii as an example to examine the phenological changes and their importance to plant production under changes in temperature and precipitation. We found that precipitation, temperature, and their interactions had no significant impact on the start of the growing season (SOS) or vegetative growth length (VGL). Precipitation had significant impacts on the end of the growing season (EOS), the length of the growing season (LOS), and reproductive growth length (RGL). The precipitation addition treatments of T2.0W + 50% (2 °C warming and 50% precipitation addition) and T1.5W + 50% (1.5 °C warming and 50% precipitation addition) significantly delayed the EOS by 6.7 d and 5.4 d, and significantly prolonged the LOS by 9.3 d and 9.3 d, respectively. Precipitation significantly changed the net CO2 assimilation rate (Pn) of the heading stage. There was no significant difference in the dry mass among all the treatments. The SOS and VGL had significant negative impacts on the dry mass of Stipa krylovii, while temperature, precipitation, and the EOS had no significant direct effect on it. Our results imply that the SOS was more important than the EOS in regulating the plant production of Stipa krylovii. This study can facilitate the understanding of the response of productivity to phenological dynamics and improve the accuracy of simulating the terrestrial ecosystem carbon budget.
1. Introduction
Plant phenology is a traditional science that studies the time of annually recurring plant life cycle events and their biotic and abiotic drivers [1,2,3,4]. As one of the most reliable bioindicators that can reflect the impact of climate change on vegetation, phenology is highly sensitive to climate change [5,6]. For vegetation in Europe from 1959 to 1996, the start of the growing season (SOS) was advanced by 6.3 days (−0.21 day/year), whereas the end of the growing season (EOS) was delayed by 4.5 days (+0.15 day/year), jointly this extended the growing season by 10.8 days [7]. In the recent three decades, 65% of SOS in the northern hemisphere grassland ecosystems has been advanced, and 58% of EOS has been delayed [8]. Climatic factors, including temperature, precipitation, and their interactions, are the dominant drivers of the variations in the SOS and EOS [8,9,10]. In Europe, the temperature of the preceding months resulted in an advance in spring/summer phenology of 2.5 days °C−1, and there was a delay in the EOS of 1.0 day °C−1 in fall. From 1971 to 2000, 78% of all leaf unfolding, flowering, and fruit ripening records advanced (30% significantly) [11]. For midlatitude (30° N–55° N) grasslands in the northern hemisphere, 23.2% of the SOS in this region was significantly advanced, and 20.5% of the EOS was significantly postponed during 1981–2014, leading to a significant prolongation trend of growing season length in 22.7% of this area. The dominant trends of the SOS and EOS in most of the region were closely related to the changes in air temperature and precipitation [12]. Currently, plant phenology variation has attracted extensive attention, but our knowledge of the exact effects of temperature, precipitation, and their interaction on phenology is incomplete.
A variety of studies have suggested that changes in plant phenology are indeed responsible for variations in ecosystem productivity and carbon sequestration [9,13,14]. A dominant positive correlation has been observed between the length of growing season (LOS) and annual gross primary productivity/net primary productivity. A one-day lengthening of the LOS increased the annual gross primary productivity by 5.8 g C m−2 yr−1 per day and the net primary productivity by 2.8 g C m−2 yr−1 per day [15]. The sensitivity of net ecosystem exchange of evergreen needleleaf forests to the LOS was 3.4 g C m−2 d−1, the net ecosystem exchange of deciduous broadleaf forests had a higher sensitivity of 5.8 g C m−2 d−1, and the net ecosystem exchange of grass/crop was the most sensitive to the LOS (7.9 g C m−2 d−1) [16]. Variability in net ecosystem exchange was significantly related to the LOS for the savanna, grassland, and tree canopy in California. However, summer drought in both mid and high latitude is probably responsible for the lower net CO2 uptake, which offsets the CO2 increase during spring [17]. In spring and autumn, gross ecosystem photosynthesis increased by the extended LOS was cancelled out by the simultaneously enhanced ecosystem respiration [18]. In northern terrestrial ecosystems, autumn warming can prolong the LOS and increase photosynthesis and respiration. Nevertheless, respiration is increased more than photosynthesis, and the CO2 lost due to autumn warming offsets 90% of the CO2 gained by early spring [19]. Thus, it is highly debatable whether a prolonged LOS will increase productivity.
As one of the most widely distributed vegetation types worldwide, grassland accounts for 40.5% of the land area (excluding Greenland and Antarctica) and plays an important role in the global carbon cycle [12,20]. Compared with forest ecosystems, grasslands, especially arid and semiarid grasslands, are more sensitive to precipitation changes and have more complex responses to climate changes [5,21]. Since 1970, the growth rate of global surface temperature has exceeded any other 50 years during the past 2000 years. The global surface temperature in 2011–2020 was 1.09 °C higher than that in 1850–1900. At least until the middle of this century, the global surface temperature is expected to continue to rise, which could also strengthen the global water cycle, such as the severity of global monsoon precipitation and dry/wet events [22]. This will inevitably lead to changes in the SOS, EOS, and LOS of grasslands, which may influence the productivity of the ecosystem, especially the arid and semiarid grassland ecosystems, and then feed back to the global climate. The typical grassland, which accounts for 10% of the total grassland of China, is fragile and sensitive to climate change [23]. Stipa krylovii is the dominant perennial grass species in the typical grassland ecosystem of north China, and it is very sensitive to precipitation change. Therefore, the study of the phenological changes of Stipa krylovii under the background of global climate change and the importance of these changes to plant production is of great significance. In this study, we studied the dominant species Stipa krylovii through a three-year simulation experiment in situ to clarify: (1) the effects of temperature and precipitation on the phenology of Stipa krylovii; (2) the changes in photosynthesis and plant production of Stipa krylovii under different temperature and precipitation conditions; and (3) the importance of the variations in the SOS and EOS to plant production.
2. Materials and Methods
2.1. Study Site
The experiment was carried out in Xilinhot, Inner Mongolia, China, at Xilinhot National Climate Observatory (44°08′03′′ N, 116°19′43′′ E, 990 m a.s.l.). The study site is a typical semiarid grassland ecosystem in northern China. This region is characterized as a temperate semiarid continental climate. A long-term (1955–2015) climate record indicates a mean annual temperature of 2.5 °C and a mean annual precipitation of 283.6 mm. The soil is chestnut soil. The experimental site is located on grassland dominated by Stipa krylovii, accompanied by Leymus chinensis and Cleistogenes squarrosa [24].
2.2. Experimental Design
Representative plots with uniform vegetation distributions were selected to carry out our experiment. We set five treatments: which include ambient temperature and precipitation (T0W0), 1.5 °C warming and 50% precipitation reduction (T1.5W − 50%), 1.5 °C warming and 50% precipitation addition (T1.5W + 50%), 2 °C warming and 50% precipitation reduction (T2.0W − 50%), and 2 °C warming and 50% precipitation addition (T2.0W + 50%). There were 4 replicates for each of the five treatments. Twenty 2 m × 2 m plots which were arranged in a 4 × 5 matrix and separated by a 2 m buffer were laid out in a randomized complete block design (Figure S1). The steel sheets were buried 1 m into the soil and protruded 0.3 m aboveground around each plot to prevent the horizontal exchange and infiltration of soil water and nutrients. The experiment began on 18 April 2019.
To simulate climate warming, we used 1 m-long infrared radiation lamps (Beijing Shiji Xingyuan Lighting Technology Co., Ltd., Chaoyang, Beijing, China), which continuously warmed the plots 24 h a day with different powers (800 W for 1.5 °C warming and 1000 W for 2.0 °C warming). Lamps were installed in the 135°-angle iron sheets and hung 2 m aboveground in the center of each warming plot. In the T0W0 plots, the same iron sheets, excluding lamps, were deployed to minimize the differences between plots.
Precipitation treatments adopted the rainfall manipulation [25] (Figure S1). The rainfall shelter used a metal frame supporting V-shaped clear acrylic bands with more than 95% light transmittance. We applied 100% perforated acrylic bands in the T0W0 treatment and 50% perforated bands and 50% acrylic bands in the T1.5W − 50% and T2.0W − 50% plots. Precipitation collected in T1.5W − 50% and T2.0W − 50% plots was evenly sprayed into each T1.5W + 50% and T2.0W + 50% plot every time it rained [24].
2.3. Soil Temperature and Water Content Measurements
In the center of each plot, an ECH2O measuring system with an EM50 data collector and three 5TM sensors (METER, Pullman, WA, USA) was installed to measure and record the soil temperature and water content. The three sensors were buried at soil depths of 0–10 cm, 10–20 cm, and 20–30 cm in each plot. Soil temperature and water content were monitored and recorded automatically every 30 min, 24 h a day.
2.4. Phenology Observation
We observed the phenology of Stipa krylovii twice a day and recorded each phenological stage [26]. The start of the growing season (SOS) was defined as the date when 50% of the Stipa krylovii restored their elasticity and turned from yellow to green. The end of the growing season (EOS) was defined as the date when two-thirds of the aboveground part of 50% of plants in the plot withered and turned yellow. The heading stage was defined as the date when 50% of plants exposed aristae from the leaf sheath. The flowering stage was defined as the date when 50% of the individual plants had anthers and dispersed their pollen. The seed formation stage was defined as the date when the upper panicles of 50% of individuals turned yellow and seeds hardened. The length of the growing season (LOS) was defined as the length between the SOS and EOS. Vegetative growth length (VGL) was defined as the length between the SOS and heading stage. Reproductive growth length (RGL) was defined as the length between the heading stage and EOS.
2.5. Leaf Gas Exchange Parameters and Dry Mass Measurements
Every year, we selected three plots from each treatment and one representative and healthy plant of each phenological stage from each selected plot to measure the net CO2 assimilation rate (Pn) (5 times a year) with an open gas exchange system (LI-6400, Li-COR Inc., Lincoln, NE, USA), which has a leaf chamber fluorometer attachment (LI6400-40, LCF). During the measurement, the saturated photosynthetic photon flux density, which was supplied by a red-blue LED, was fixed at 1500 μmol·m−2·s−1, the concentration of CO2 was set at 400 μmol·mol−1, the temperature was maintained at 25 °C, and the air relative humidity was controlled between 50% and 70% [27,28]. At the end of the growing season, the aboveground parts of the selected plants were clipped after the measurement, oven-dried at 80 °C for at least 48 h to constant weight, and weighed to obtain the dry mass (DM) [24].
2.6. Statistical Analysis
A three-way ANOVA with LSD (least significant difference) tests was used to test the main and interactive effects of year, temperature, and precipitation on soil temperature, soil water content, phenology, the net CO2 assimilation rate (Pn) at each stage, and dry mass of Stipa krylovii. A structural equation model (SEM) was conducted by Amos 21.0 (Amos Development, Spring House, PA, USA) to explore the effects of the start of the growing season (SOS) and the end of the growing season (EOS) on the plant production of Stipa krylovii. The statistical significance level was set at p-values less than 0.05 unless stated otherwise.
3. Results
3.1. Changes in Soil Water Content and Temperature
In the experimental period (from 18 April 2019 to 2 November 2021), the dynamics of the soil temperature (ST) and water content (SWC) of the five treatments were similar (Figure S1A,B). Temperature, precipitation, and year significantly changed SWC. The treatments of 50% precipitation addition significantly enhanced the SWC of the three growing seasons (May to November), while 50% precipitation reduction significantly decreased it (Table S1, Figure S2). Year and precipitation significantly changed ST, and the 1.5 °C and 2 °C warming treatments significantly increased ST in the three growing seasons (Table S1, Figure S2).
3.2. Changes in the Phenology of Stipa krylovii
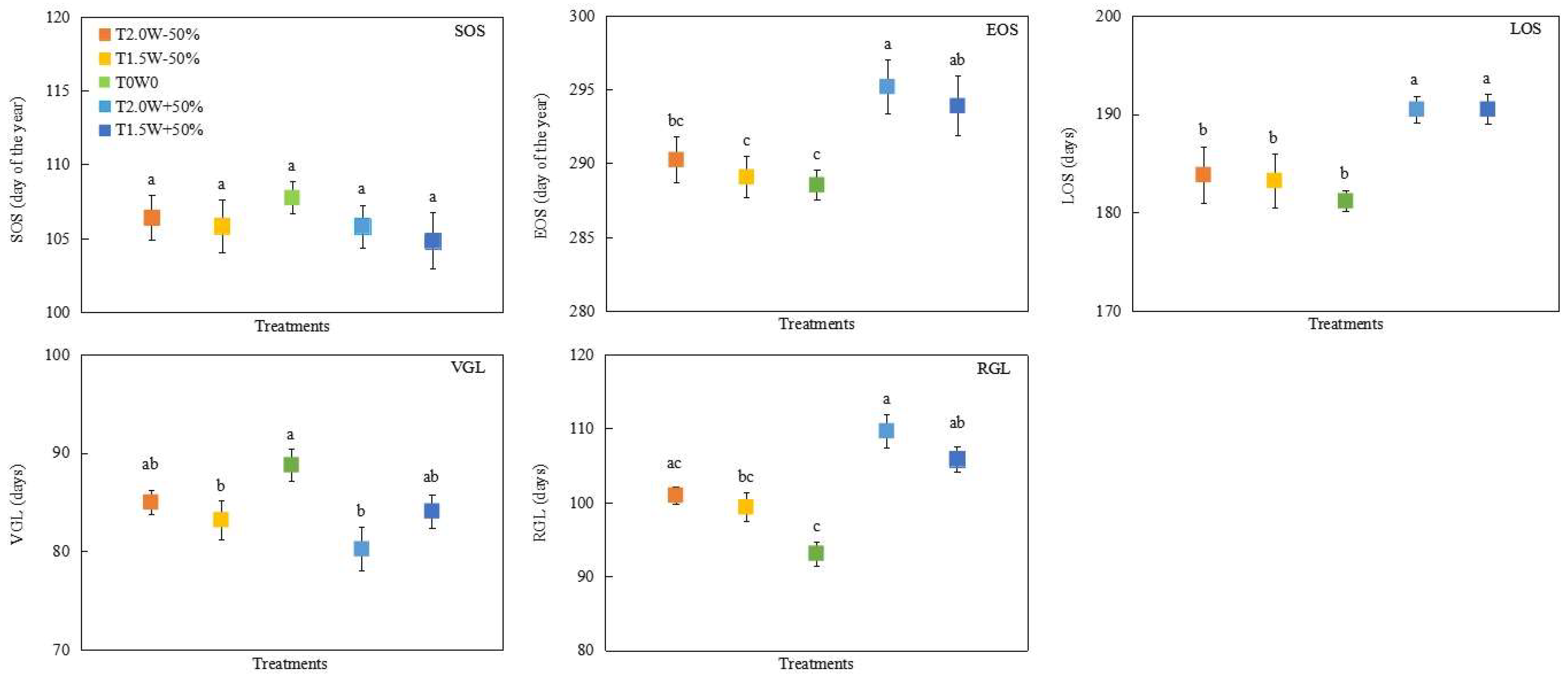
Year significantly influenced the start of the growing season (SOS) and vegetative growth length (VGL), while precipitation, temperature, and their interactions had no significant impact on them (Table 1). Year, precipitation, and their interactions had significant impacts on the end of the growing season (EOS), the length of growing season (LOS), and reproductive growth length (RGL), but temperature had no significant effect on them (Table 1). Precipitation addition treatments (T2.0W + 50% and T1.5W + 50%) significantly delayed the EOS and prolonged the LOS (Table 1, Figure 1). The EOS of T2.0W + 50% and T1.5W + 50% were 6.65 d and 5.35 d later than that of T0W0, respectively (Figure 1). The LOS of T2.0W + 50% and T1.5W + 50% were 9.32 d and 9.32 d longer than that of T0W0, respectively (Figure 1). The VGL of T0W0 was significantly longer than the VGL of T1.5W − 50% and T2.0W + 50%, and the RGL of T0W0 was significantly shorter than those of T1.5W + 50% and T2.0W + 50% (Figure 1).
3.3. Responses of the Net CO2 Assimilation Rate at Different Stages
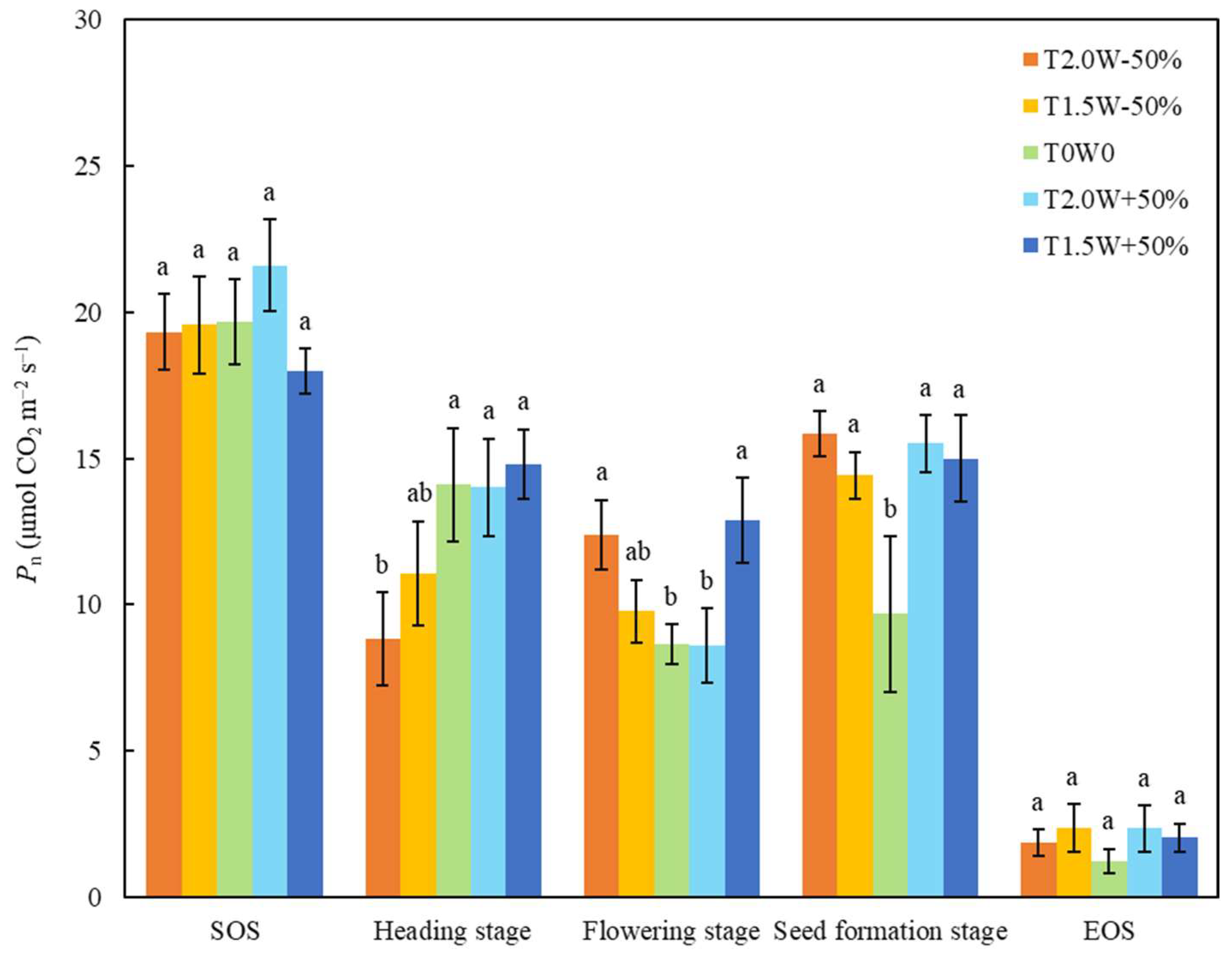
Year significantly changed the net CO2 assimilation rate (Pn) of all the stages except the flowering stage (Table 2). Precipitation significantly changed the Pn of the heading stage (Pnh), and the Pnh of the T2.0W − 50% treatment was significantly lower than those of the other treatments (Table 2, Figure 2). The main effects of year, temperature, and precipitation were not significant on Pn at the flowering stage (Pnf), but the interactive effects were significant (Table 2). The Pnf values of T2.0W − 50% and T1.5W + 50% were significantly higher than those of T0W0 and T2.0W + 50% (Figure 2). The Pn at the seed formation stage (Pns) in the T0W0 plots were significantly lower than that in the other treatments (Figure 2).
3.4. Changes in the Above-Ground Dry Mass of Stipa krylovii
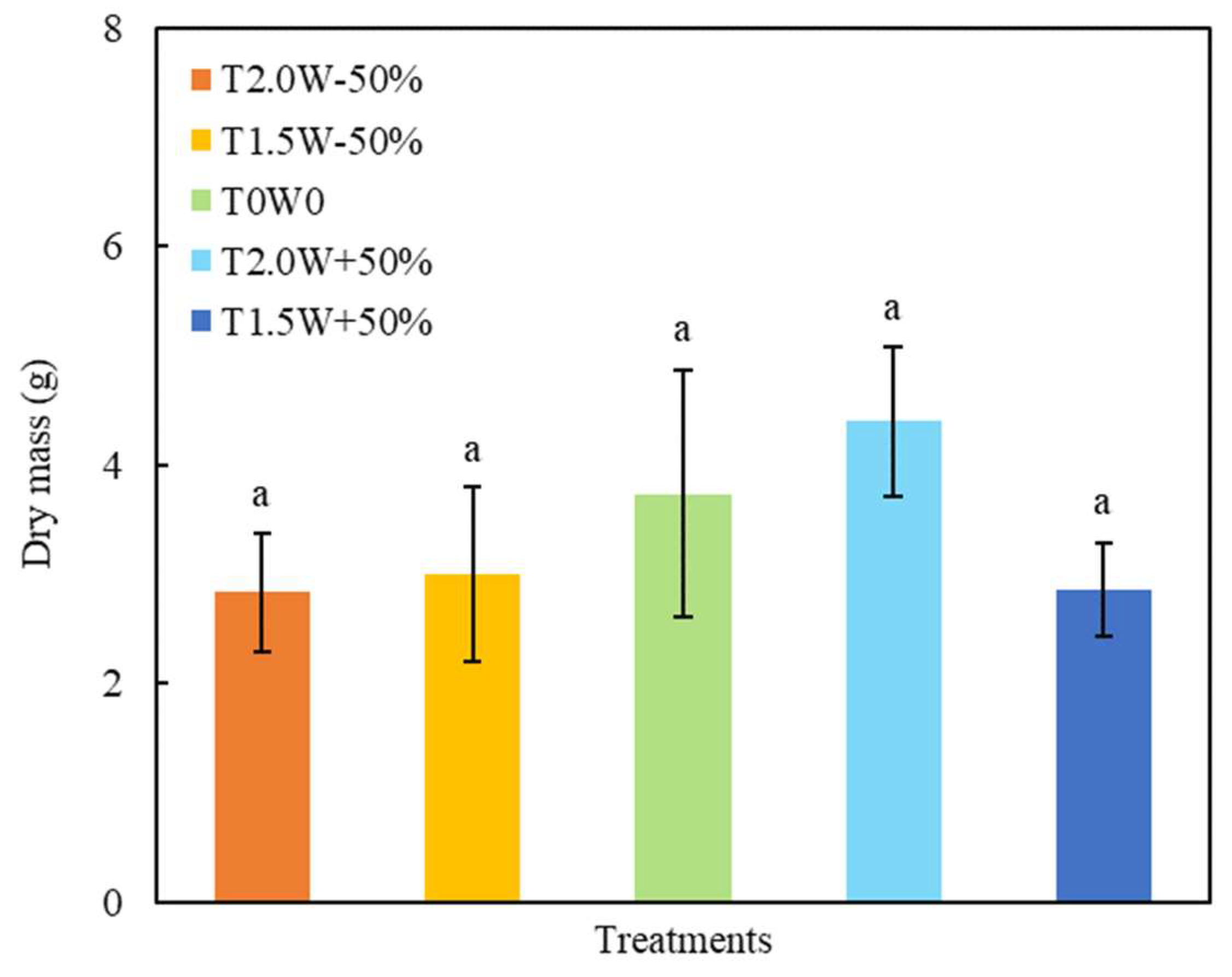
The dry mass was only significantly influenced by year (Table 2). There was no significant difference among the dry mass of all the treatments (Figure 3). The dry masses of the T2.0W − 50%, T1.5W − 50%, T0W0, T2.0W + 50%, and T1.5W + 50% plots were 2.83 g, 3.00 g, 3.73 g, 4.40 g, and 2.86 g, respectively.
3.5. Factors Affecting Plant Production of Stipa krylovii
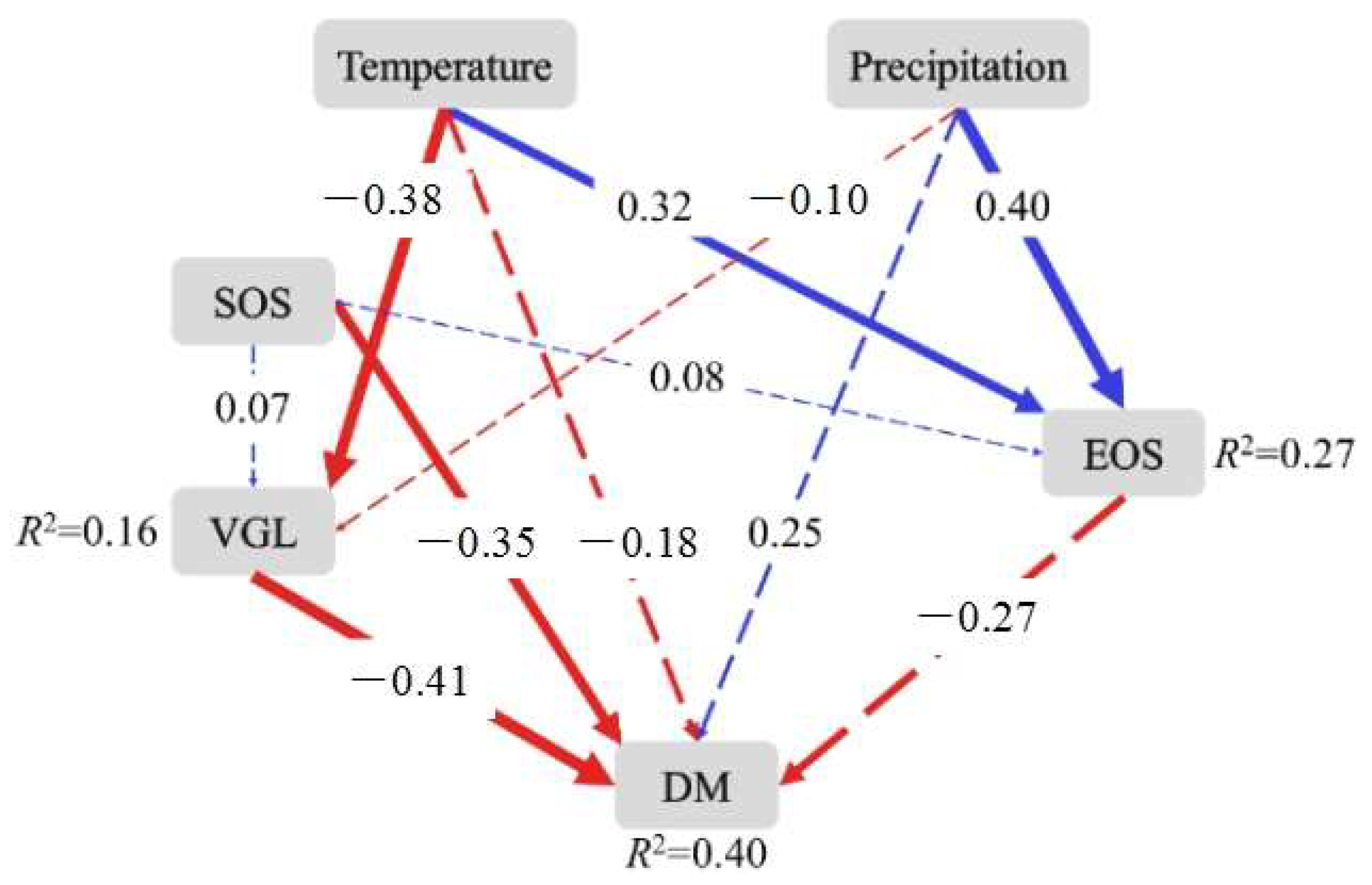
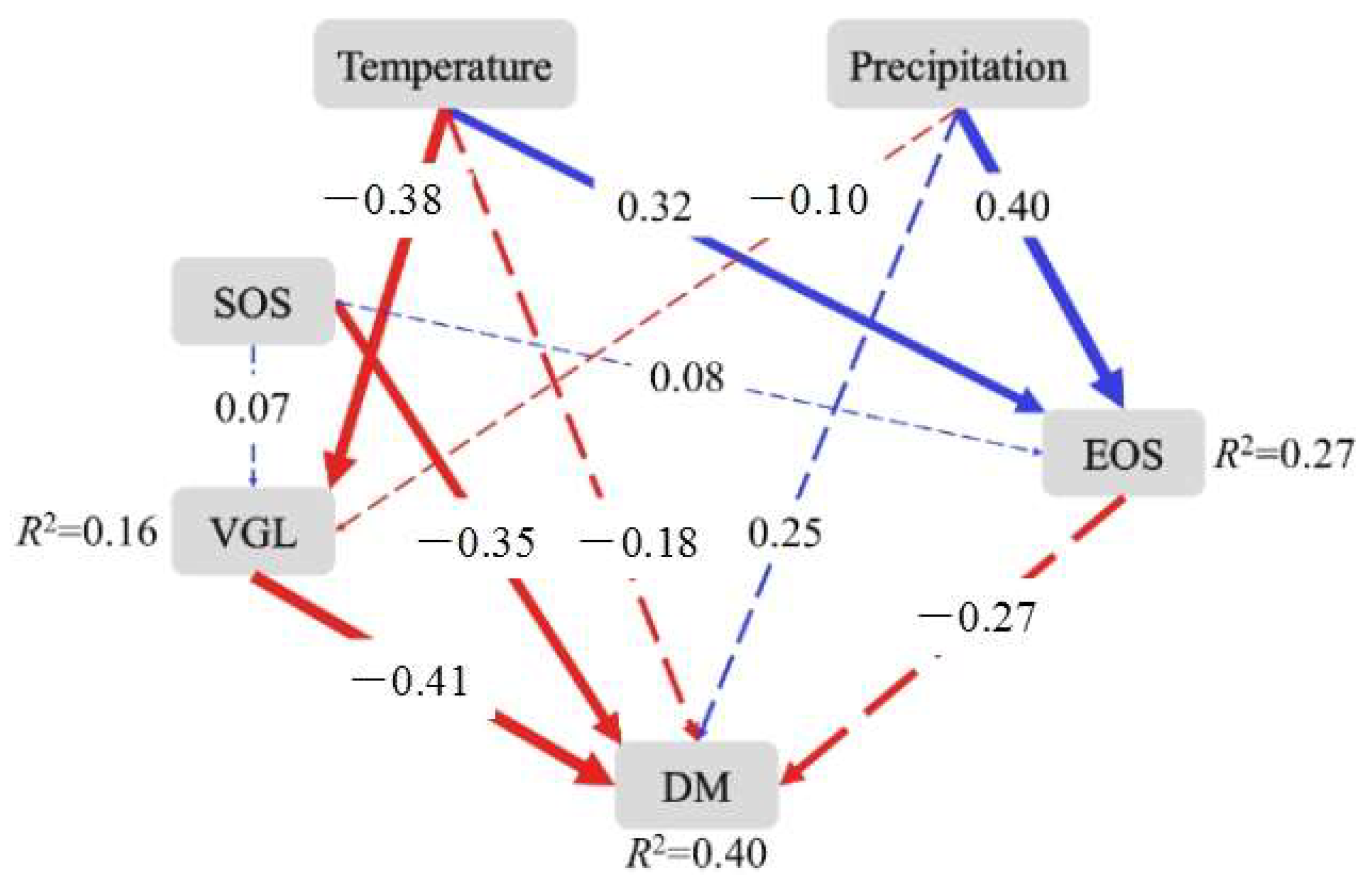
Structural equation models (SEM) were established to analyze the direct and indirect effects of the SOS and EOS on the dry mass of Stipa krylovii, with an explanation of total variance in dry mass of 40%. The results of the SEM models showed that the SOS and vegetative growth length (VGL) had significant negative relationships with the dry mass of Stipa krylovii, with standardized path coefficients of −0.35 and −0.41, respectively. Temperature, precipitation, and the EOS had no significant relationship with dry mass (Figure 4). In addition, temperature and precipitation had significant positive impacts on the EOS, and temperature had significantly negative impacts on the VGL.
4. Discussion
We found that precipitation and temperature had no significant impact on the SOS, which is different from previous research. According to previous studies, temperature has a dual effect on spring phenology: Low temperature is necessary to induce and break endodormancy, and high temperature can break ecodormancy to promote the growth of buds [29,30]. In winter and spring (from 1985 to 2003), with the increase in temperature, the SOS of Leymus chinensis and Stipa krylovii was postponed [23]. Precipitation changed the SOS of plants in the semi-arid grassland of southern Africa [31]. The observation indicated that precipitation was related to SOS [32]. This inconformity may be caused by other factors such as wind speed, relative humidity, sunshine hours, etc. The control factors of SOS and its regulation mechanism are worthy of attention. In the future, we will further study the influence of other factors on SOS, other than temperature and precipitation, and find out the factors that cause the interannual variation of SOS.
Our results showed that precipitation had significant effects on the EOS, which is consistent with previous studies. The research pointed out that an increase in temperature in summer and autumn significantly delayed the timing of leaf senescence of European beech, delaying it by 6–8 d for every 1 °C increase in temperature [33]. Without the limitation of water and nutrients, temperature may be the main factor controlling the leaf senescence of European beech. In Finnish Lapland, no correlation between the EOS and climatic factors by investigating plant phenology from 1997 to 2006 was found [34]. At middle and high latitudes, temperature and photoperiod are the two key regulators of phenology, but at a regional scale, water limitations may be the driver [35]. The phenology of arid areas was more sensitive to interannual variation in preseason precipitation than that of mesic areas [36].
In the present study, precipitation addition treatments significantly delayed the EOS and prolonged the LOS, but there was no significant change in the dry mass of Stipa krylovii among all treatments. Further analysis showed that the SOS and VGL had significant correlations with dry mass, while the EOS had no significant correlation with dry mass. These results indicate that the SOS plays a more important role in regulating the plant production of Stipa krylovii than the EOS. Plant phenology has been regarded as a regulator of the global carbon cycle [37,38,39]. Many previous studies have proposed that the productivity increase might be attributable to the earlier SOS or later EOS [40,41,42,43]. However, respiration can offset the improvement of productivity due to the extended LOS [18,19]. The extended LOS caused by the delayed EOS will not enhance carbon gain in forests because of the limit of the declines in photosynthetic capacity [44]. On the other hand, the response models of plant productivity to phenological changes depended on the location [45]. An earlier SOS may increase plant productivity and can also decrease productivity, and the same goes for the later EOS. Moreover, the relationship between phenological dynamics and productivity may also be influenced by local climatic (such as temperature and precipitation), topographic conditions, and many other factors (such as photoperiod, winter chilling, permafrost degradation and snowmelt, soil moisture, nutrient limitation, and human disturbance) [46]. This study can provide new evidence for the accurate estimation of the terrestrial ecosystem carbon budget.
5. Conclusions
Plant phenology is of vital importance to the carbon cycle of terrestrial ecosystems. We conducted a three-year in situ simulation experiment in the Stipa krylovii steppe to explore the influence of phenological changes on plant production. The results suggested that precipitation, temperature, and their interactions did not significantly influence the start of the growing season (SOS) or vegetative growth length (VGL). Precipitation significantly changed the end of the growing season (EOS) and the length of the growing season (LOS). The precipitation addition treatments of T2.0W + 50% and T1.5W + 50% significantly delayed the EOS by 6.65 d and 5.35 d, and significantly prolonged the LOS by 9.32 d and 9.32 d, respectively. There was no significant difference among the dry mass of all the treatments. The SOS had significant direct impacts on the dry mass of Stipa krylovii, while the EOS had no significant direct effect on it, indicating that the SOS contributed more than the EOS to the variation in plant production. This study can provide reference for the assessment of the plant production and carbon budget of terrestrial ecosystems.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy12123208/s1, Figure S1. The layout of the plots (A) and the plot photo (B). Figure S2. Soil water content (A) and soil temperature (B) of each treatment during the experimental period (from 18 April 2019 to 2 November 2021) and the effects of precipitation and temperature treatments on the soil water content (C) and soil temperature (D) of the three growing seasons (from May to November) (means ± SE). W0: ambient precipitation, W + 50%: 50% precipitation addition, W − 50%: 50% precipitation reduction; T0: ambient temperature, T1.5: 1.5 °C warming, T2.0: 2.0 °C warming. The different lowercase letters show significant differences among precipitation and temperature treatments (p < 0.05). Table S1. Main and interactive effects of the year (Y), temperature (T), and precipitation (Pre) on the soil water content (SWC) and soil temperature (ST).
Author Contributions
H.Y.: Conceptualization, Methodology, Software, Formal analysis, Investigation, Writing—original draft, Visualization. G.Z.: Conceptualization, Methodology, Writing—review and editing, Supervision, Project administration, Funding acquisition. X.L.: Design of manipulation experiment and long-term experimental investigation, resources. Q.H.: Conceptualization, Methodology, Writing—review and editing, Project administration, Funding acquisition. M.Z.: Methodology, Investigation, Resources, Writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This study is supported by the National Natural Science Foundation of China (No. 42130514), National Key Research and Development Program of China (No. 2018YFA0606103), and the Basic Research Fund of Chinese Academy of Meteorological Sciences (2020Z004, 2022Y015).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors wish to thank Mingxin, Hu, Siqi Wang, Shiya Zhang, Wenjie Gu, Ruochen Li, Xingyang Song Erhua Liu, Huailin Zhou, Chunli Dong, Jianghua Su, Shilin Cui, Junjie He, Zhenjie Guo, Ping Yin, Huirong Li, Ruojing Wang, Tianhe Lan, Liwei Zhang, Jing Xu, Yanyan Lu, Guolan Zhang, Yingshun Wang and Ruiyan Dong for their assistance in the manipulation experiment.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Azizan, F.A.; Astuti, I.S.; Aditya, M.I.; Febbiyanti, T.R.; Williams, A.; Young, A.; Abdul Aziz, A. Using multi-temporal satellite data to analyse phenological responses of rubber (Hevea brasiliensis) to climatic variations in South Sumatra, Indonesia. Remote Sens. 2021, 13, 2932. [Google Scholar] [CrossRef]
- Lang, W.; Chen, X.; Qian, S.; Liu, G.; Piao, S. A new process-based model for predicting autumn phenology: How is leaf senescence controlled by photoperiod and temperature coupling? Agric. For. Meteorol. 2019, 268, 124–135. [Google Scholar] [CrossRef]
- Lieth, H. Phenology and Seasonality Modeling; Springer: Berlin/Heidelberg, Germany, 1974; pp. 3–19. [Google Scholar]
- Ren, S.; Chen, X.; Pan, C. Temperature-precipitation background affects spatial heterogeneity of spring phenology responses to climate change in northern grasslands (30°N–55°N). Agric. For. Meteorol. 2022, 315, 108816. [Google Scholar] [CrossRef]
- Fan, D.; Zhao, X.; Zhu, W.; Sun, W.; Qiu, Y. An improved phenology model for monitoring green-up date variation in Leymus chinensis steppe in Inner Mongolia during 1962–2017. Agric. For. Meteorol. 2020, 291, 108091. [Google Scholar] [CrossRef]
- Fu, Y.H.; Piao, S.; Zhou, X.; Geng, X.; Hao, F.; Vitasse, Y.; Janssens, I.A. Short photoperiod reduces the temperature sensitivity of leaf-out in saplings of Fagus sylvatica but not in horse chestnut. Glob. Change Biol. 2019, 25, 1696–1703. [Google Scholar] [CrossRef]
- Menzel, A. Trends in phenological phases in Europe between 1951 and 1996. Int. J. Biometeorol. 2000, 44, 76–81. [Google Scholar] [CrossRef]
- Cong, N.; Huang, K.; Zhang, Y. Unsynchronized driving mechanisms of spring and autumn phenology over Northern Hemisphere Grasslands. Front. For. Glob. Change 2021, 3, 610162. [Google Scholar] [CrossRef]
- Caffarra, A.; Donnelly, A.; Chuine, I. Modelling the timing of Betula pubescens budburst. II. Integrating complex effects of photoperiod into process-based models. Clim. Res. 2011, 46, 159–170. [Google Scholar] [CrossRef]
- Peng, J.; Wu, C.; Zhang, X.; Wang, X.; Gonsamo, A. Satellite detection of cumulative and lagged effects of drought on autumn leaf senescence over the Northern Hemisphere. Glob. Change Biol. 2019, 25, 2174–2188. [Google Scholar] [CrossRef]
- Menzel, A.; Sparks, T.H.; Estrella, N.; Koch, E.; Aasa, A.; Ahas, R.; Alm-Kübler, K.; Bissolli, P.; Braslavská, O.; Briede, A.; et al. European phenological response to climate change matches the warming pattern. Glob. Change Biol. 2006, 12, 1969–1976. [Google Scholar] [CrossRef]
- Ren, S.; Li, Y.; Peichl, M. Diverse effects of climate at different times on grassland phenology in mid-latitude of the Northern Hemisphere. Ecol. Indic. 2020, 113, 106260. [Google Scholar] [CrossRef]
- Bucher, S.F.; Römermann, C.; Bonser, S. The timing of leaf senescence relates to flowering phenology and functional traits in 17 herbaceous species along elevational gradients. J. Ecol. 2021, 109, 1537–1548. [Google Scholar] [CrossRef]
- Chang, Q.; Xiao, X.; Jiao, W.; Wu, X.; Doughty, R.; Wang, J.; Du, L.; Zou, Z.; Qin, Y. Assessing consistency of spring phenology of snow-covered forests as estimated by vegetation indices, gross primary production, and solar-induced chlorophyll fluorescence. Agric. For. Meteorol. 2019, 275, 305–316. [Google Scholar] [CrossRef]
- Piao, S.; Friedlingstein, P.; Ciais, P.; Viovy, N.; Demarty, J. Growing season extension and its impact on terrestrial carbon cycle in the Northern Hemisphere over the past 2 decades. Glob. Biogeochem. Cycles 2007, 21. [Google Scholar] [CrossRef]
- Churkina, G.; Schimel, D.; Braswell, B.H.; Xiao, X. Spatial analysis of growing season length control over net ecosystem exchange. Glob. Change Biol. 2005, 11, 1777–1787. [Google Scholar] [CrossRef]
- Angert, A.; Biraud, S.; Bonfils, C.; Henning, C.C.; Buermann, W.; Pinzon, J.; Tucker, C.J.; Fung, I. Drier summers cancel out the CO2 uptake enhancement induced by warmer springs. Proc. Natl. Acad. Sci. USA 2005, 102, 10823–10827. [Google Scholar] [CrossRef] [Green Version]
- Richardson, A.D.; Black, T.A.; Ciais, P.; Delbart, N.; Friedl, M.A.; Gobron, N.; Hollinger, D.Y.; Kutsch, W.L.; Longdoz, B.; Luyssaert, S.; et al. Influence of spring and autumn phenological transitions on forest ecosystem productivity. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 3227–3246. [Google Scholar] [CrossRef] [Green Version]
- Piao, S.; Ciais, P.; Friedlingstein, P.; Peylin, P.; Reichstein, M.; Luyssaert, S.; Margolis, H.; Fang, J.; Barr, A.; Chen, A.; et al. Net carbon dioxide losses of northern ecosystems in response to autumn warming. Nature 2008, 451, 49–52. [Google Scholar] [CrossRef]
- Chen, X.; Li, J.; Xu, L.; Liu, L.; Ding, D. Modeling greenup date of dominant grass species in the Inner Mongolian Grassland using air temperature and precipitation data. Int. J. Biometeorol. 2013, 58, 463–471. [Google Scholar] [CrossRef]
- Ren, S.; Yi, S.; Peichl, M.; Wang, X. Diverse responses of vegetation phenology to climate change in different grasslands in Inner Mongolia during 2000–2016. Remote Sens. 2017, 10, 17. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; in press; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., Eds.; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Yuan, W.; Zhou, G.; Wang, Y.; Han, X.; Wang, Y. Simulating phenological characteristics of two dominant grass species in a semi-arid steppe ecosystem. Ecol. Res. 2007, 22, 784–791. [Google Scholar] [CrossRef]
- Yu, H.; Zhou, G.; Lv, X.; He, Q.; Zhou, M. Environmental factors rather than productivity drive autumn leaf senescence: Evidence from a grassland in situ simulation experiment. Agric. For. Meteorol. 2022, 327, 109221. [Google Scholar] [CrossRef]
- Yahdjian, L.; Sala, O.E. A rainout shelter design for intercepting different amounts of rainfall. Oecologia 2002, 133, 95–101. [Google Scholar] [CrossRef] [PubMed]
- National Meteorological Administration. Agrometeorological Observation Criterion; Meteorological Press: Beijing, China, 1993. [Google Scholar]
- Frechette, E.; Chang, C.Y.; Ensminger, I. Variation in the phenology of photosynthesis among eastern white pine provenances in response to warming. Glob. Change Biol. 2020, 26, 5217–5234. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Zhou, G.; Shimizu, H. Are plant growth and photosynthesis limited by pre-drought following rewatering in grass? J. Exp. Bot. 2009, 60, 3737–3749. [Google Scholar] [CrossRef] [Green Version]
- Piao, S.; Liu, Q.; Chen, A.; Janssens, I.A.; Fu, Y.; Dai, J.; Liu, L.; Lian, X.; Shen, M.; Zhu, X. Plant phenology and global climate change: Current progresses and challenges. Glob. Change Biol. 2019, 25, 1922–1940. [Google Scholar] [CrossRef]
- Soolanayakanahally, R.Y.; Guy, R.D.; Silim, S.N.; Song, M. Timing of photoperiodic competency causes phenological mismatch in balsam poplar (Populus balsamifera L.). Plant Cell Environ. 2013, 36, 116–127. [Google Scholar] [CrossRef]
- Cho, M.A.; Ramoelo, A.; Dziba, L. Response of land surface phenology to variation in tree cover during green-up and senescence periods in the semi-arid savanna of Southern Africa. Remote Sens. 2017, 9, 689. [Google Scholar] [CrossRef] [Green Version]
- Shen, M.; Piao, S.; Cong, N.; Zhang, G.; Jassens, I. A Precipitation impacts on vegetation spring phenology on the Tibetan Plateau. Glob. Change Biol. 2015, 21, 3647–3656. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.H.; Piao, S.; Delpierre, N.; Hao, F.; Hanninen, H.; Liu, Y.; Sun, W.; Janssens, I.A.; Campioli, M. Larger temperature response of autumn leaf senescence than spring leaf-out phenology. Glob. Change Biol. 2018, 24, 2159–2168. [Google Scholar] [CrossRef]
- Pudas, E.; Tolvanen, A.; Poikolainen, J.; Sukuvaara, T.; Kubin, E. Timing of plant phenophases in Finnish Lapland in 1997–2006. Boreal Environ. Res. 2008, 13, 31–43. [Google Scholar] [CrossRef] [Green Version]
- Jolly, W.M.; Nemani, R.; Running, S.W. A generalized, bioclimatic index to predict foliar phenology in response to climate. Glob. Change Biol. 2005, 11, 619–632. [Google Scholar] [CrossRef]
- Zhou, J.; Cai, W.; Qin, Y.; Lai, L.; Guan, T.; Zhang, X.; Jiang, L.; Du, H.; Yang, D.; Cong, Z.; et al. Alpine vegetation phenology dynamic over 16 years and its covariation with climate in a semi-arid region of China. Sci. Total Environ. 2016, 572, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Crabbe, R.A.; Dash, J.; Rodriguez-Galiano, V.F.; Janous, D.; Pavelka, M.; Marek, M.V. Extreme warm temperatures alter forest phenology and productivity in Europe. Sci. Total Environ. 2016, 563, 486–495. [Google Scholar] [CrossRef] [PubMed]
- Ganjurjav, H.; Hu, G.; Gornish, E.; Zhang, Y.; Li, Y.; Yan, Y.; Wu, H.; Yan, J.; He, S.; Danjiu, L.; et al. Warming and spring precipitation addition change plant growth pattern but have minor effects on growing season mean gross ecosystem productivity in an alpine meadow. Sci. Total Environ. 2022, 841, 156712. [Google Scholar] [CrossRef]
- Zhou, L.; Wang, Y.; Jia, Q.; Zhou, G. Increasing temperature shortened the carbon uptake period and decreased the cumulative net ecosystem productivity in a maize cropland in Northeast China. Field Crops Res. 2021, 267, 108150. [Google Scholar] [CrossRef]
- Bao, G.; Chen, J.; Chopping, M.; Bao, Y.; Bayarsaikhan, S.; Dorjsuren, A.; Tuya, A.; Jirigala, B.; Qin, Z. Dynamics of net primary productivity on the Mongolian Plateau: Joint regulations of phenology and drought. Int. J. Appl. Earth Obs. Geoinf. 2019, 81, 85–97. [Google Scholar] [CrossRef]
- Ge, W.; Han, J.; Zhang, D.; Wang, F. Divergent impacts of droughts on vegetation phenology and productivity in the Yungui Plateau, southwest China. Ecol. Indic. 2021, 127, 107743. [Google Scholar] [CrossRef]
- Jeong, S.J.; Ho, C.H.; Gim, H.J.; Brown, M.E. Phenology shifts at start vs. end of growing season in temperate vegetation over the Northern Hemisphere for the period 1982–2008. Glob. Change Biol. 2011, 17, 2385–2399. [Google Scholar] [CrossRef]
- Wu, C.; Gough, C.M.; Chen, J.M.; Gonsamo, A. Evidence of autumn phenology control on annual net ecosystem productivity in two temperate deciduous forests. Ecol. Eng. 2013, 60, 88–95. [Google Scholar] [CrossRef]
- Bauerle, W.L.; Oren, R.; Way, D.A.; Qian, S.S.; Stoy, P.C.; Thornton, P.E.; Bowden, J.D.; Hoffman, F.M.; Reynolds, R.F. Photoperiodic regulation of the seasonal pattern of photosynthetic capacity and the implications for carbon cycling. Proc. Natl. Acad. Sci. USA 2012, 109, 8612–8617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Hou, X.; Peng, D.; Gonsamo, A.; Xu, S. Land surface phenology of China’s temperate ecosystems over 1999–2013: Spatial–temporal patterns, interaction effects, covariation with climate and implications for productivity. Agric. For. Meteorol. 2016, 216, 177–187. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, B.; Yang, Q.; Chen, G.; Yang, B.; Lu, L.; Shen, M.; Peng, Y. Responses of net primary productivity to phenological dynamics in the Tibetan Plateau, China. Agric. For. Meteorol. 2017, 232, 235–246. [Google Scholar] [CrossRef]
Figure 1.
Effects of temperature and precipitation treatments on phenological change of Stipa krylovii (means ± SE). T0W0: ambient temperature and precipitation; T1.5W − 50%: 1.5 °C warming and 50% precipitation reduction; T1.5W + 50%: 1.5 °C warming and 50% precipitation addition; T2.0W − 50%: 2 °C warming and 50% precipitation reduction; T2.0W + 50%: 2 °C warming and 50% precipitation addition. SOS represents the start of the growing season; EOS represents the end of the growing season; LOS represents the length of growing season; VGL represents vegetative growth length; RGL represents reproductive growth length. Different lowercase letters were used to indicate significant differences among treatments (p < 0.05).
Figure 1.
Effects of temperature and precipitation treatments on phenological change of Stipa krylovii (means ± SE). T0W0: ambient temperature and precipitation; T1.5W − 50%: 1.5 °C warming and 50% precipitation reduction; T1.5W + 50%: 1.5 °C warming and 50% precipitation addition; T2.0W − 50%: 2 °C warming and 50% precipitation reduction; T2.0W + 50%: 2 °C warming and 50% precipitation addition. SOS represents the start of the growing season; EOS represents the end of the growing season; LOS represents the length of growing season; VGL represents vegetative growth length; RGL represents reproductive growth length. Different lowercase letters were used to indicate significant differences among treatments (p < 0.05).

Figure 2.
Effects of temperature and precipitation treatments on the net CO2 assimilation rate (Pn) at different stages (means ± SE). Different lowercase letters were used to indicate significant differences among treatments (p < 0.05).
Figure 2.
Effects of temperature and precipitation treatments on the net CO2 assimilation rate (Pn) at different stages (means ± SE). Different lowercase letters were used to indicate significant differences among treatments (p < 0.05).
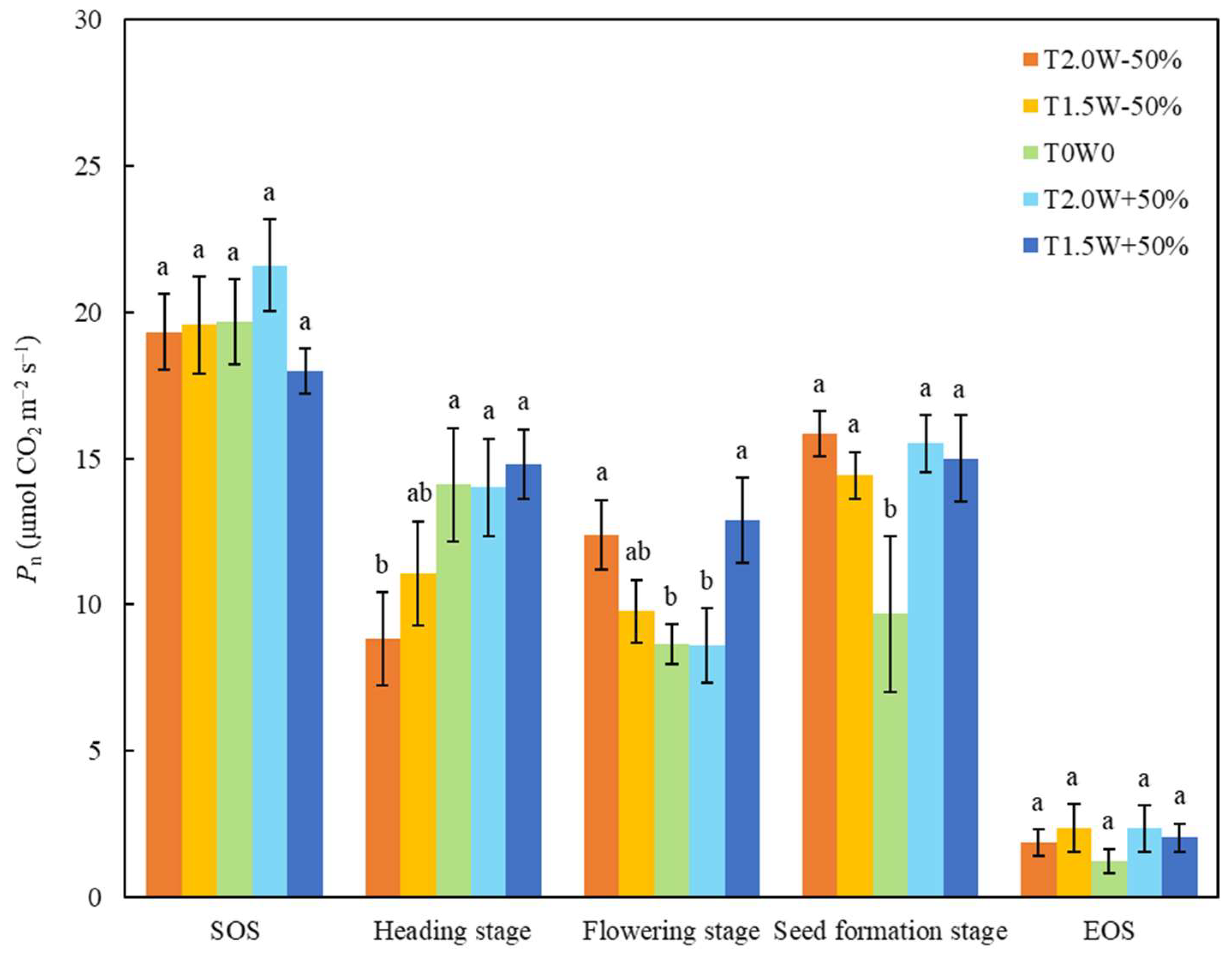
Figure 3.
Effects of temperature and precipitation treatments on dry mass (means ± SE). The letter “a” in the figure was used to indicate significant differences among treatments (p < 0.05).
Figure 3.
Effects of temperature and precipitation treatments on dry mass (means ± SE). The letter “a” in the figure was used to indicate significant differences among treatments (p < 0.05).

Figure 4.
Effects of the start of the growing season (SOS) and the end of the growing season (EOS) on the plant production of Stipa krylovii. VGL represents vegetative growth length; DM represents dry mass. χ2 = 3.27, df = 4, p = 0.51, AI = 49.27, RMSEA (root mean square error of approximation) <0.001. The red and blue arrows indicate negative and positive correlations, respectively, and arrow thickness represents the strength of correlation. The solid and dotted lines indicate significant (p < 0.05) and insignificant effects (p > 0.05), respectively. Values on the arrows indicate standardized path coefficients. The R2 values next to the response variables represent the proportion of variation explained by relationships with other variables.
Figure 4.
Effects of the start of the growing season (SOS) and the end of the growing season (EOS) on the plant production of Stipa krylovii. VGL represents vegetative growth length; DM represents dry mass. χ2 = 3.27, df = 4, p = 0.51, AI = 49.27, RMSEA (root mean square error of approximation) <0.001. The red and blue arrows indicate negative and positive correlations, respectively, and arrow thickness represents the strength of correlation. The solid and dotted lines indicate significant (p < 0.05) and insignificant effects (p > 0.05), respectively. Values on the arrows indicate standardized path coefficients. The R2 values next to the response variables represent the proportion of variation explained by relationships with other variables.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Main and interactive effects of the year (Y), temperature (T), and precipitation (Pre) on the phenology of Stipa krylovii.
Table 1.
Main and interactive effects of the year (Y), temperature (T), and precipitation (Pre) on the phenology of Stipa krylovii.
| SOS | EOS | LOS | VGL | RGL | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| df | F | p | F | p | F | P | F | p | F | p | |
| Y | 2 | 64.24 | <0.001 | 22.22 | <0.001 | 20.84 | <0.001 | 4.16 | <0.05 | 19.64 | <0.001 |
| T | 1 | 0.92 | 0.34 | 1.83 | 0.18 | 0.10 | 0.75 | 0.83 | 0.37 | 0.99 | 0.33 |
| Pre | 1 | 0.92 | 0.34 | 49.57 | <0.001 | 29.38 | <0.001 | 1.35 | 0.25 | 12.53 | <0.001 |
| Y × T | 2 | 1.80 | 0.18 | 0.46 | 0.64 | 0.21 | 0.81 | 0.99 | 0.38 | 0.86 | 0.43 |
| Y × Pre | 2 | 0.83 | 0.44 | 36.99 | <0.001 | 16.58 | <0.001 | 1.19 | 0.32 | 6.76 | <0.001 |
| T × Pre | 1 | 0.06 | 0.80 | 0.01 | 0.93 | 0.02 | 0.90 | 2.90 | 0.10 | 0.12 | 0.73 |
| Y × T × Pre | 2 | 0.23 | 0.80 | 0.26 | 0.77 | 0.03 | 0.98 | 2.56 | 0.09 | 2.16 | 0.13 |
Note: SOS represents the start of the growing season; EOS represents the end of the growing season; LOS represents the length of growing season; VGL represents vegetative growth length; RGL represents reproductive growth length. The values of p < 0.05 are bolded.
Table 2.
Main and interactive effects of the year (Y), temperature (T), and precipitation (Pre) on the net CO2 assimilation rate (Pn) and dry mass of Stipa krylovii.
Table 2.
Main and interactive effects of the year (Y), temperature (T), and precipitation (Pre) on the net CO2 assimilation rate (Pn) and dry mass of Stipa krylovii.
| PnSOS | Pnh | Pnf | Pns | PnEOS | DM | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| df | F | p | F | p | F | p | F | P | F | p | F | p | |
| Y | 2 | 6.82 | <0.01 | 13.15 | <0.001 | 1.21 | 0.31 | 17.54 | <0.001 | 7.01 | <0.01 | 7.70 | <0.01 |
| T | 1 | 1.05 | 0.31 | 1.86 | 0.19 | 1.18 | 0.29 | 0.92 | 0.35 | 0.05 | 0.83 | 1.07 | 0.31 |
| Pre | 1 | 0.23 | 0.64 | 10.41 | <0.01 | 0.13 | 0.72 | 0.24 | 0.63 | 0.64 | 0.43 | 1.18 | 0.29 |
| Y × T | 2 | 0.64 | 0.53 | 0.66 | 0.52 | 5.57 | <0.01 | 2.79 | 0.08 | 0.02 | 0.98 | 0.08 | 0.93 |
| Y × Pre | 2 | 0.65 | 0.53 | 1.71 | 0.20 | 3.83 | <0.05 | 3.23 | 0.06 | 0.02 | 0.98 | 0.19 | 0.83 |
| T × Pre | 1 | 2.01 | 0.17 | 0.13 | 0.73 | 18.67 | <0.001 | 0.58 | 0.45 | 0.97 | 0.34 | 1.73 | 0.20 |
| Y × T × Pre | 2 | 0.23 | 0.80 | 0.33 | 0.72 | 10.41 | <0.001 | 5.63 | <0.05 | 1.32 | 0.27 | 1.90 | 0.17 |
Note: PnSOS represents Pn at the start of the growing season; Pnh represents Pn at the heading stage; Pnf represents Pn at the flowering stage; Pns represents Pn at the seed formation stage; PnEOS represents Pn at the end of the growing season; DM represents dry mass. The values of p < 0.05 are bolded.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yu, H.; Zhou, G.; Lv, X.; He, Q.; Zhou, M. Effects of Phenological Changes on Plant Production—From the View of Stipa krylovii. Agronomy 2022, 12, 3208. https://doi.org/10.3390/agronomy12123208
AMA Style
Yu H, Zhou G, Lv X, He Q, Zhou M. Effects of Phenological Changes on Plant Production—From the View of Stipa krylovii. Agronomy. 2022; 12(12):3208. https://doi.org/10.3390/agronomy12123208
Chicago/Turabian StyleYu, Hongying, Guangsheng Zhou, Xiaomin Lv, Qijin He, and Mengzi Zhou. 2022. "Effects of Phenological Changes on Plant Production—From the View of Stipa krylovii" Agronomy 12, no. 12: 3208. https://doi.org/10.3390/agronomy12123208
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.