
Research Progress in Leaf Related Molecular Breeding of Cucurbitaceae
1
Collaborative Innovation Center for Efficient and Green Production of Agriculture in Mountainous Areas of Zhejiang Province, College of Horticulture Science, Zhejiang Agriculture and Forestry University, Hangzhou 311300, China
2
College of Horticulture, Qingdao Agricultural University, Qingdao 266109, China
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Agronomy 2022, 12(11), 2908; https://doi.org/10.3390/agronomy12112908
Submission received: 1 November 2022
/
Revised: 16 November 2022
/
Accepted: 16 November 2022
/
Published: 21 November 2022
(This article belongs to the Special Issue Breeding Innovations in Crop for Resilient Cropping Systems)
Abstract
:Cucurbitaceae crops play an important role in agricultural production, and they are a primary source of vegetables and fruits for daily needs. The cultivation of cucurbit varieties with excellent agronomic characters has attracted much attention in recent years. As the main source energy organ of Cucurbitaceae, the development of leaves is closely related to yield and environmental response. In this paper, the main research achievements on leaf development of Cucurbitaceae plants were summarized, and the origin, genetic research, gene mapping, and effects on the agronomic and economic characters of the leaf shape and leaf color mutants of Cucurbitaceae were introduced. In addition, the effects of some hormones and environmental factors were reviewed. This work provides a reference point for molecular breeding of leaf development of Cucurbitaceae.
1. Introduction
Cucurbitaceae, as one of the most important edible plants in the world, has more than 800 species, among which the representative plants are cucumber, pumpkin, melon, and watermelon. As important economic crops, they have not only edible value but also medicinal value [1]. Leaves are the main organ of Cucurbitaceae for photosynthesis. The shape and color of leaves affect the efficiency of photosynthesis and respiration. For example, the larger the leaf area, the more light energy it absorbs, and the more nutrients can be produced. The thinner the leaves, the more favorable the plant is for respiration. However, if the leaves are too wide and thin, it is not conducive to retaining water, that is, the leaf area and leaf thickness are limited by water supply [2]. The change in leaf color is caused by the change in chlorophyll and anthocyanin in the leaf, and it further affects photosynthesis [3]. Therefore, leaf development plays a crucial role in the process of gas exchange, nutrient accumulation, and water transport in plants. A leaf begins to differentiate from the shoot apical meristem first, and then the leaf primordium develops along the proximal–distal, abaxial–adaxial, and medial–lateral to establish the three-dimensional polarity of the leaf [4]. After the three-dimensional polarity of the leaf is established, the leaves begin to expand and grow to maturity [5]. The leaf shape of plants shows diversity under the influence of genes, hormones, and environment. Different species of Cucurbitaceae have different leaf shapes. Even in the same plant, the leaf shape changes because of different developmental stages and environments. At present, few comprehensive summaries on Cucurbitaceae leaf development can be found. Thus, in the present paper, the genes, hormones, and environmental factors that affect the development of Cucurbitaceae leaves and the molecular regulation mechanism of Cucurbitaceae leaf development were reviewed.
2. Genes Affect Leaf Development
2.1. Leaf Shape Mutant
Research on key genes regulating the leaf development of Cucurbitaceae plants is of great importance to reveal the mechanism of leaf development and breeding of Cucurbitaceae plants. There seems to be a close relationship between leaves and the development of reproductive organs in Cucurbitaceae. For example, male and female sterile mango fruit (mf) mutants in cucumber also have abnormal development of cotyledons and true leaves, and cotyledons appear narrower, wrinkled, downward cupped, and darker than the wild-type. The leaf expansion of true leaves was significantly reduced [6]. Leaf shape is controlled by complex interactions between genetic networks. The genetic mechanism of the leaf shape characteristics of Cucurbitaceae plants is not completely clear, but some representative leaf-shaped mutants of Cucurbitaceae were found. The identification of key genes from leaf shape mutants is critically important to decipher the functions of some genes regulating leaf traits.
2.1.1. Lobed Leaf
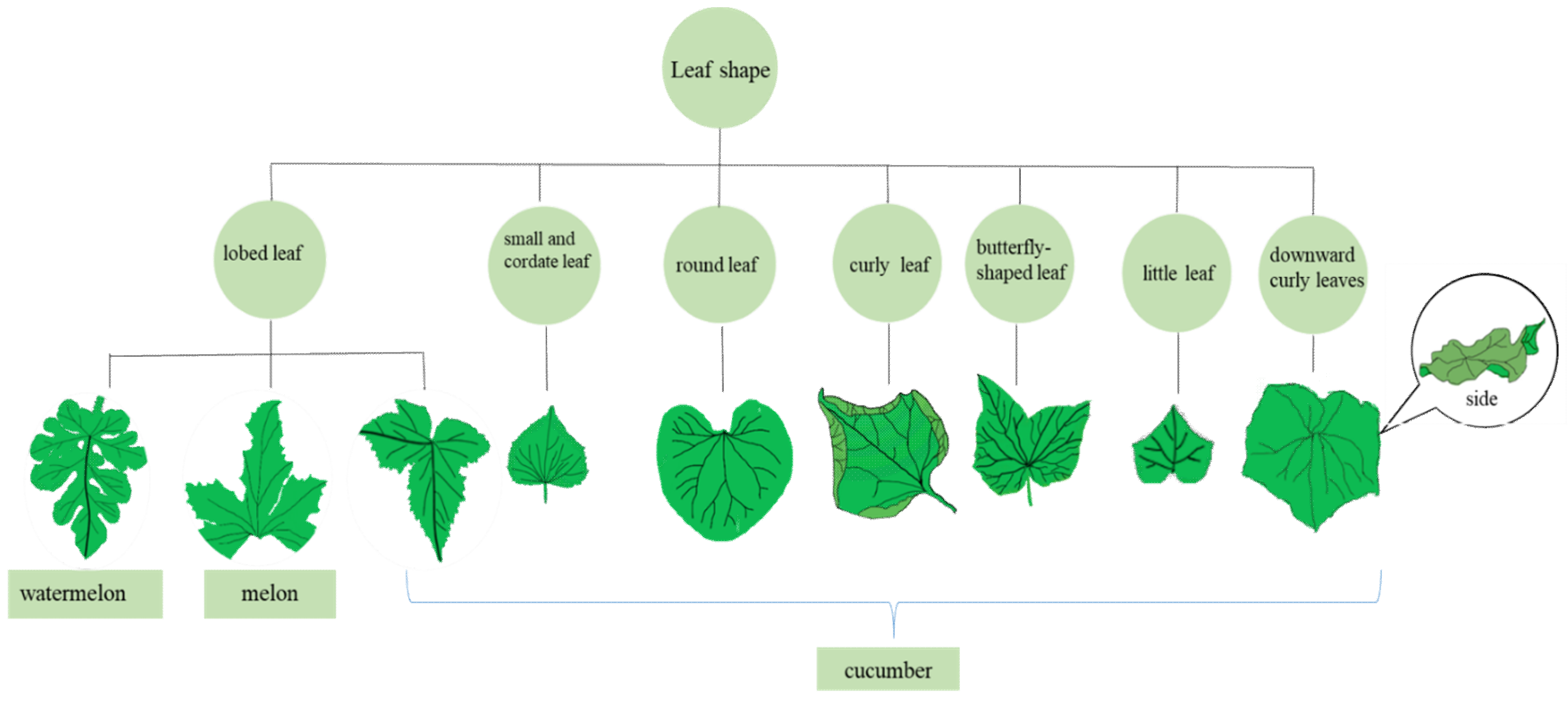
In cucumber, a high proportion of the leaves in CsHAN1-OE and CsHAN1-RNAi lines were highly lobed [7] (Figure 1). After ectopic expression of CsHAN1 in Arabidopsis, CsHAN1 could restore the smooth leaves of the han-2 mutant in the Col background to the jagged shape of Col. The rosette leaves and cauline leaves of transgenic lines overexpressing CsHAN1 in Arabidopsis wild-type (WT) Col were serrated leaves. These findings indicate that the elaborate expression of HAN is vital for the development of leaf shape, and the increase or decrease in this expression could cause changes in leaf margin. A natural mutant, named BM7, with palmately lobed leaves, was identified in the melon line Mhy with normal round leaves [8]. Genetic analysis demonstrated that the palmately lobed leaf trait was controlled by a pair of recessive genes pll. The BM7 plants possessed lower vitality and occupied less space than the melon lines with normal round leaves, so planting in greenhouses is more suitable. Furthermore, pll is located on the melon linkage group (LG) III, between the SSR marker G69 and 784RS, and MELO3C010784 was the only gene predicted in this region (Table 1).The lobed leaf character of watermelon was controlled by a single dominant gene, and the candidate gene was identified as Cla018360 [9].
2.1.2. Round Leaf
Two round-leaf mutants C356 and C949 appeared after EMS mutagenesis of the cucumber inbred line CCMC [10]. The true leaf margins of the two mutants were smooth and round (Figure 1), and some mutant plants had abnormal cotyledon development, characterized by polycotyledon or asymmetric development and female infertility. According to genetic analysis, the C356 and C949 mutants are controlled by single recessive nuclear genes Csrl-1 and Csrl-2, respectively. Remarkably, CSrl-1 and CSrl-2 are different mutation sites of the same candidate gene CsPID (Csa1G537400). The CsPID mutants were obtained by simultaneous mutagenesis in two other laboratories. Liu et al. [11] identified a stable genetic round-leaf mutant rl of offspring by EMS mutagenesis of cucumber “649”. The rl mutant leaves were round, and they lacked leaf formation in some nodes. In addition, defects were observed in the development of the tendrils and floral organs of rl mutants. The first 20 nodes did not produce tendrils, pollen viability decreased, and the females were sterile. Song et al. [12]. found that the number of lateral roots increased, and that the growth of primary roots decreased in the round-leaf mutant (rl). The mutant gene, Csa1M537400, encodes a PINOID kinase protein involved in auxin transport. The expression of rl mutant Csa1M537400 was significantly reduced compared with that of the WT, and IAA was expressed at higher levels in several tissues.
2.1.3. Curly Leaf
Two curly leaf mutants cul-1 and cul-2 in the M2 population were induced by EMS of the cucumber inbred line CCMC (Figure 1). Through map-based cloning, cul-1 and cul-2 were found to be allelic mutations, and the candidate gene was CsPHB (Csa6G525430). The mutation site was the binding site of cs-miRNA165/166. The expression of CsPHB in the root, stem, true leaf, and flower of the mutant was higher than that of the WT, but the expression of cs-miR165/166 in the mutant was lower. This finding indicated a negative correlation between the expression of the two genes, so the destruction of miRNA165/166 binding is likely to lead to the functional acquisition mutation of HD-ZIP III. Whether cs-miR165/166 regulates cucumber leaf development by targeting CsPHB remains to be further researched [13].
2.1.4. Small and Cordate Leaf
The Csa7G062760 gene is a candidate gene from a cucumber small and cordate leaf 1 (scl1) mutant [14]. The mutant is heart-shaped with reduced leaf area without the auriculate leaf base and protruded middle margin (Figure 1). The scl1 mutant is speculated to encode a nucleoside bisphosphate phosphatase that is involved in the regulation of leaf development and may not involve known hormonal pathways.
2.1.5. Little Leaf
The cucumber littleleaf (ll) mutant had smaller leaves, flowers, fruits, and seeds, and added lateral branches when compared to the WT cucumber (Figure 1). The candidate gene ORF4 had pleiotropic effects, and its single base mutation resulted in the conversion of amino acid 264 from tryptophan to glycine, leading to the heteromorphosis of multiple organs in the mutant. The overexpression of ll in WT cucumber can significantly increase the size of flowers, fruits, seeds, and epidermal cells, and the expression of ll in overexpression lines was much higher than that in the control. These results indicated that ll can affect the size of cucumber organs [15].
2.1.6. Downward Curly Leaves
Interference with cucumber from the CsIVP-RNA interference (RNAi) mutant can cause defective development of leaf veins, with a reduced angle between leaf midvein and vertical axis, increased angle between leaf primary veins, and increased secondary veins and bending of the leaf midvein, which all lead to the downward curl of the leaf (Figure 1). In addition, the leaf margins overlapped without clearance. Some defects were also found in the floral organ development of the mutant plants, including the shortening of mature fruit length and a decrease in the number of seeds [16].
2.1.7. Butterfly-Shaped Leaves
In cucumber, CsWOX1 was expressed in the early stage of leaf primordium and in the middle mesophyll and leaf margin. The leaves of the mango fruit (mf) mutant from the CsWOX1 gene mutation were narrower than those of WT (AM218) leaves, and the ratio of leaf length to width was significantly larger than that of AM218 leaves. Among the CsWOX1-OE line, it is obvious that the leaf size was significantly smaller than that of WT (CU2), and that a small number of butterfly-shaped leaves with abnormal veins were produced (Figure 1). The analysis of the cucumber mf rl (round leaf) double mutant showed that CsWOX1 played a role in leaf vein development by controlling auxin transport through PINOID (CsPID1). In addition, CsWOX1 regulates leaf size through CIN-TCP transcription factors [17].
2.2. Leaf Color Mutants
Leaf color mutations are ubiquitous among Cucurbitaceae plants. Leaf color is mainly affected by various pigments inside the leaves, including chlorophyll, carotenoids, and anthocyanins. Changes in the type and ratio of pigments inside the leaves lead to the formation of leaf color mutants [18,19]. Leaf color mutations can be divided into natural mutations and artificial mutations. In accordance with the degree of leaf color mutations and their effects on plants, they can be divided into albino death, yellow-green, post-green, mottles, and spots [20]. With cucumber as an example, the leaf color mutants found so far were chlorophyll deficiency (cd), golden (g), golden cotyledons (gc), light green cotyledons-1 (lg-1), light green cotyledons-2 (lg-2), variegated virescence (vvi), yellow cotyledons-1 (yc-1), yellow cotyledons-2 (yc-2), virescent-2 (v-2), virescent1 (v-1), virescent (v), pale lethal (pl) and albino cotyledon (al), light sensitive (ls) [21,22,23,24,25]. The reciprocal interspecific hybrids of wild cucumber and cultivated cucumber all had a new phenotype of yellow leaves, but the causes were different. Chlorophyll biosynthesis of orthogonal hybrids is limited by the process of transformation from Pchlide to Chl a, and the chlorophyll biosynthesis of anti-cross hybrids was blocked at the step of converting PBG to urogen-III, which significantly decreased the chlorophyll content and showed a novel phenotype of yellow leaves [26]. Song [27] identified a cucumber mutant vyl (virescent-yellow leaf) that exhibited reduced pigment content and delayed chloroplast development, controlled by a single recessive gene, Csa4G637110.
The yellow cotyledons of the EMS-induced mutant yl2.1 did not turn green as the leaves grew [28]. Niu et al. located al on cucumber chromosome 7 by finely mapping the mutant al of cucumber albino leaf mutants [29]. The content of various pigments in cucumber yellow-green leaf mutant C777 was significantly lower than that in the WT, and the net photosynthetic rate of the mutant was significantly lower [30]. The leaves of cucumber yellow-green leaf mutant C528 were yellow from the cotyledon stage, and this color did not change with development. The photosynthetic pigment mass fraction and net photosynthetic efficiency were significantly lower than those of the WT [31]. In addition, some other Cucurbitaceae plants have leaf color mutations. For example, watermelon plants carry an albinism lethal gene [32]. In pumpkin silver leaf mutant 48a, the mature leaves were silver-gray, the chlorophyll content was significantly reduced, the epidermal cells and palisade cells on the leaves were obviously peeled off, and obvious gaps were found [33]. In yellow-to-green mutant (MT) of thin-skinned melon, the leaf color changed from yellow to yellow-green during the process of growth and development. The veins were green, and the chlorophyll content was significantly lower than that of the non-mutant. The main cause of leaf yellowing is speculated to be the reduction in chlorophyll content caused by the imperfect development of chloroplast structure, and the yellowing-to-green mutation is controlled by a stealth gene [34]. In spontaneous cucumber leaf virescent mutant SC311Y, whose cotyledons and real leaves are yellow and gradually turn green, light and temperature can affect the process of turning green. The mutant is male and sterile, and the real leaves have ruffled leaves with white stripes. The mutant gene Csa3G042730 regulates chloroplast development and reduces the content of chlorophyll and carotenoid in the mutant [35]. A cucumber yellow young leaf mutant C777 was produced by EMS mutagenesis. The mutant cotyledons and newborn true leaves are yellow, and there are green spots on the real leaves, and the leaves gradually turn green with the growth. Here, CsHD is its mutant gene, and this nonsense mutation introduced a stop codon and, thus, a premature protein. The expression of CsHD in the mutant was significantly decreased [36].
2.3. Plant Hormones Affect Leaf Development
As important regulatory factors affecting leaf shape, plant hormones are critical in plant life activities [37]. Plant hormones include auxin, gibberellin, cytokinin, and other hormones. Auxin is indispensable in almost all plant developments, such as leaf development, fruitification, and blossoming. It regulates leaf initiation, leaf formation, and compound leaf configuration [38]. Some research reported that the symmetry of the front and back of leaves and the symmetry of both sides are affected by auxin [39,40]. The round leaf-mutant caused by a single recessive gene rl is similar to the leaves of WT plants treated with auxin transport inhibitors, indicating that rl plays a key role in the biosynthesis, transport, and response of auxin in cucumber [12]. The mutant gene of the cucumber rl round-leaf mutant is CsPID, which regulates the auxin signaling pathway by phosphorylating PIN. Decreased auxin (IAA) levels were detected in specific tissues of rl round-leaf mutants [11]. Vascular bundles or veins in leaves were arranged in the form of branches, which determine the leaf shape to a great extent. Auxin can not only differentiate vascular cells but also induces vascular bundle formation. The deletion of CsIVP affected the development of vascular bundles, increased the accumulation of auxin leaf veins, and led to the variation in leaf shape. The process of leaf senescence was also regulated by plant hormones [41]. Ethylene, jasmonic acid, auxin, and cytokinin were proved to be related to leaf senescence, among which cytokinin plays an important role in initiating and preventing leaf senescence. Bangerth, K.-F. [42] found that exogenous spraying of synthetic cytokinin, growth polar transport inhibitor, or gibberellin biosynthesis inhibitor could delay or prevent leaf senescence, whereas exogenous spraying gibberellin could accelerate it. The induction of auxin can instantly increase the expression of Aux/IAA. The expression of CsaIAAs can be induced by cytokinin 6-benzylaminopurine (6-BA) in most organs, especially in roots, stems, leaves, and tendrils. Therefore, cytokinin can also induce the expression of auxin, also indicating that auxin and cytokinin play a coordinated role in the development and senescence of leaves [43].

{kind=link}
{kind=link}
Table 1.
Genes involved in leaf shape regulation of cucurbitaceae.
| Mocular Marker | Gene Name | Species | Traits | Reference |
|---|---|---|---|---|
| —— | CsHAN1 | Cucumber | Lobed leaf | Ding et al. (2015) [7] |
| G69 and 784RS | Pll (MELO3C010784) | Melon | Lobed leaf | Gao et al. (2014) [8] |
| W07164 and W07061 | Cl LL1 (Cla018360) | Watermelon | Lobed leaf | Chen (2018) [9] |
| Chr1: 16 164 573–21 583 426 | CsPID1 (Csa1G537400) | Cucumber | Round leaf | Zhang et al. (2018) [10] Liu et al. (2019) [11] |
| dCAPS-01 and NWSSR0024 | CsPHB (Csa6G525430) | Cucumber | Curly leaf | Rong et al. (2018) [13] |
| SNP7G3596452 | scl1 (Csa7G062760) | Cucumber | Small and cordate leaf | Gao et al. (2017) [14] |
| SSR21758 and UW083795 | ORF4 | Cucumber | Littleleaf | Yang et al. (2018) [15] |
| —— | CsIVP | Cucumber | Downward curly leaves | Yan et al. (2020) [16] |
| UW026993 and UW027011 | CsWOX1 (Csa1G042780) | Cucumber | Butterfly-shaped leaves | Niu et al. (2018) [6] Wang et al. (2020) [17] |
3. Environmental Factors
As the main lateral organ of Cucurbitaceae, leaves are sensitive to the environment. Different environments may affect leaf development, leading to the change in leaf shape. For example, the palmately split-leaf melon BM7 showed split leaves only when planted in the field [8]. In this section, cucumber was taken as an example to illustrate the effects of various environmental factors on leaves, including light, temperature, and CO2 concentration.
3.1. Temperature Affects Leaf Development
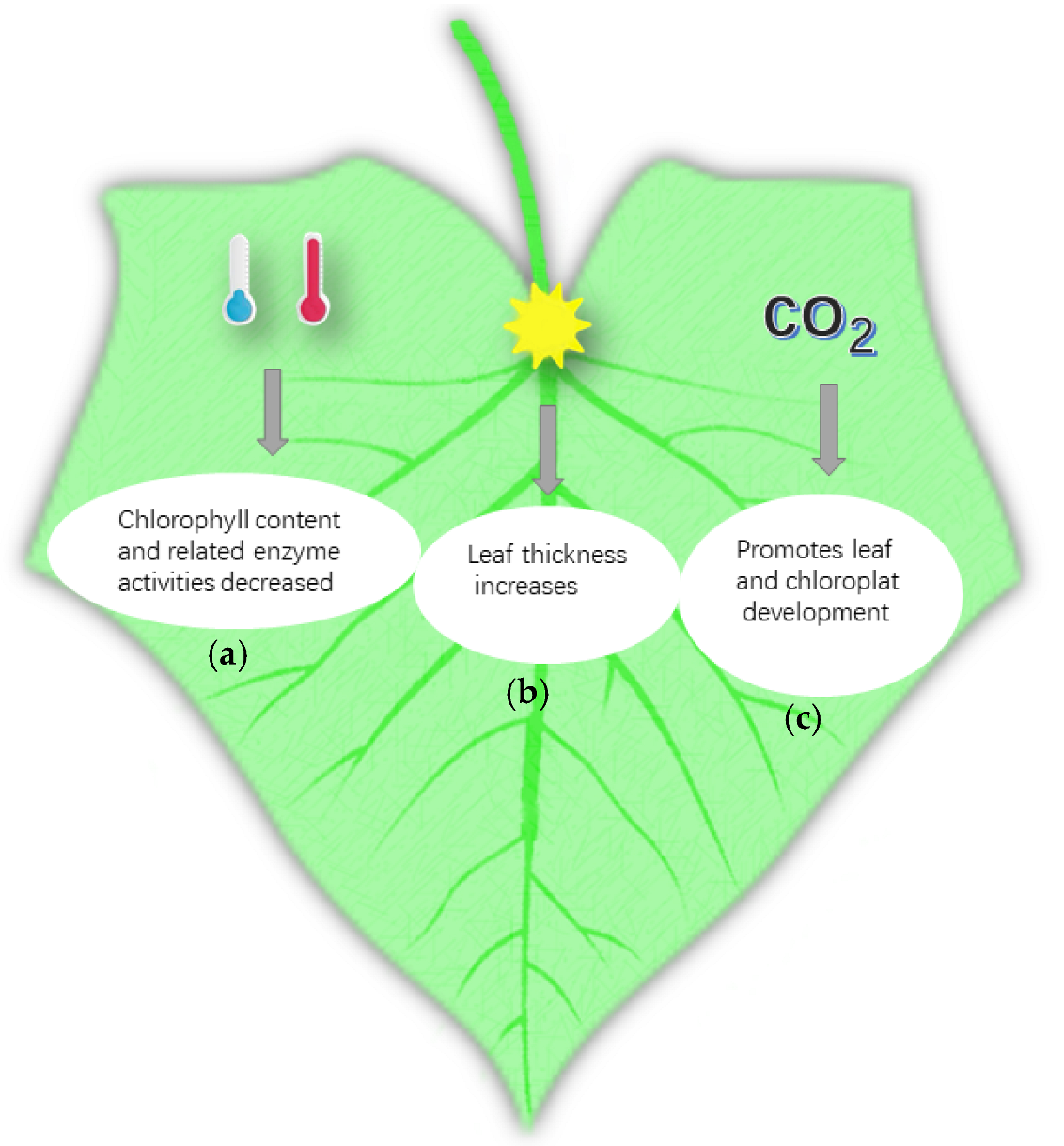
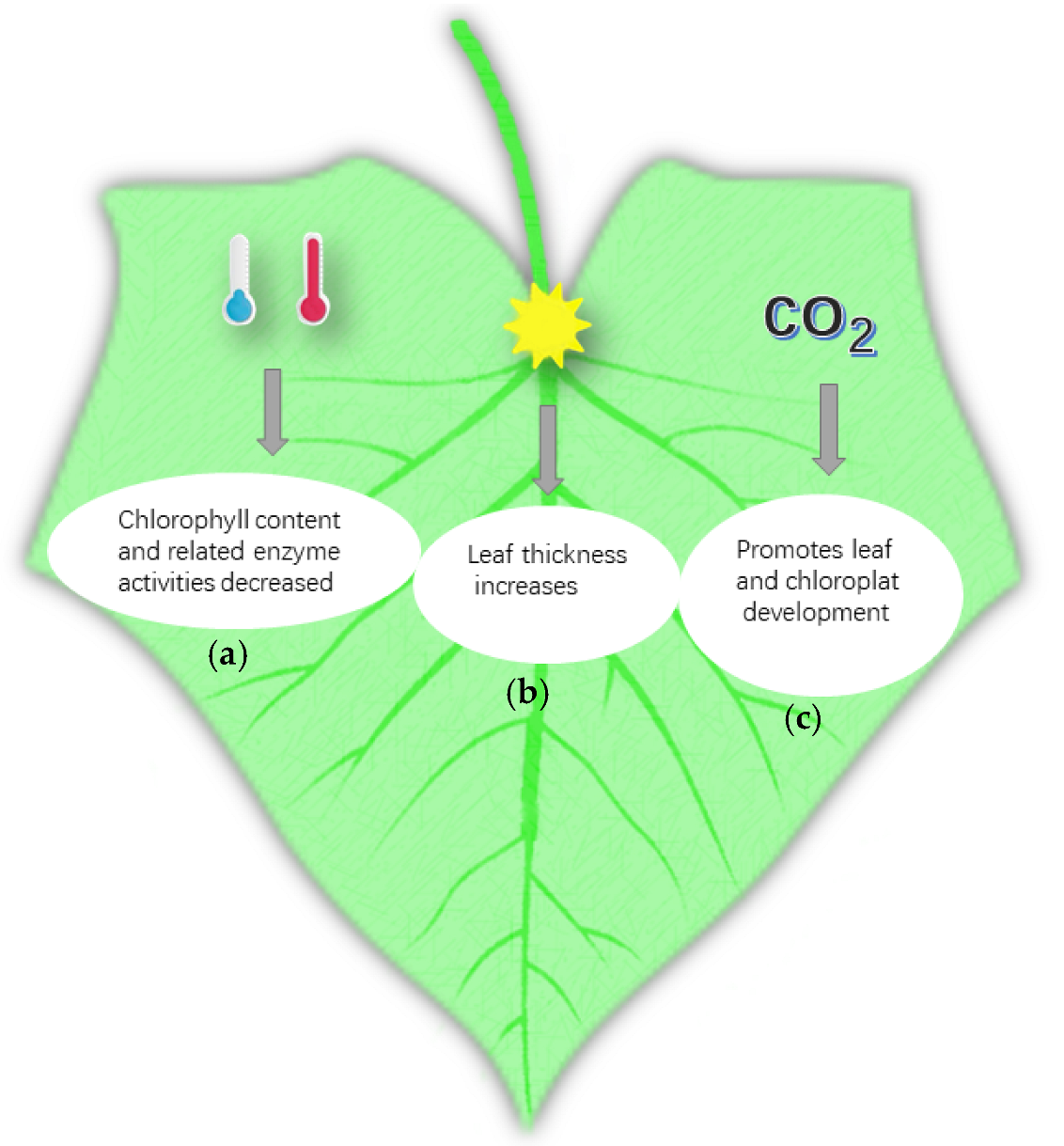
Cucumber is sensitive to low temperatures and, hence, it is mainly distributed in subtropical regions. Therefore, low-temperature chilling injury is a major obstacle in cucumber cultivation. Cucumber plants need to have various organ-sensitive and physiological responses to cope with the environment, and the structure of their leaves can change due to the influence of external temperature [44,45]. Under low-temperature conditions, the chlorophyll content in cucumber leaves, the activities of various reactive enzymes, photosynthesis, and net photosynthetic rate decreased. The activities of the chloroplast and the mitochondria in cucumber cells also decreased, and the degree of decrease in these indicators varies in different development stages [46]. The relative indices of cucumber leaves at different developmental stages under low-temperature stress include leaf electrolyte permeability, chlorophyll content, photosynthesis, stomatal resistance, transpiration rate, intercellular CO2 concentration, MDA content, soluble sugar content, and proline content. The low-temperature resistance of leaves at the seedling stage was stronger, and plants at this stage were found to have a higher physiological metabolism and to easily recover under normal temperature by comparing the cold resistance indices of cucumber at different developmental stages [47]. In fact, high-temperature stress is just oxidative stress. Plants accumulate reactive oxygen species under such stress, which causes membrane lipid peroxidation and destroys their membrane system [48]. The effects of high-temperature stress on leaf senescence characteristics and endogenous hormones of cucumber at the flowering stage indicated that short-term heat stress increased the cucumber leaf chlorophyll content and leaf antioxidant enzyme activity [49]. With the increase in temperature and stress time, the content of chlorophyll and the activity of antioxidant enzymes in leaves can decrease gradually. With the increase in high-temperature stress, the cucumber seedling’s leaf net photosynthetic rate, chlorophyll content, and soluble sugar content showed a trend of gradual decline, and the intercellular CO2 concentration and the content of malondialdehyde (MDA) in leaves showed a trend of slow decline after the first rise. Transpiration showed an increasing trend which decreased after the first rise and later under high-temperature stress. Meanwhile, the decrease in net photosynthetic rate and chlorophyll content slowed down, indicating that the cucumber seedlings had certain resistance. Various studies have shown that the chlorophyll content and related enzyme activities in cucumber leaves decreased in low- and high-temperature environments, and that cucumber has self-regulation ability to adapt to environmental temperature under high- or low-temperature stress [50] (Figure 2a).
3.2. Effect of Light on Leaf Development
Photosynthesis is one of the most important physiological activities of green plants. Light is one of the necessary conditions for photosynthesis, so it has an important influence on the growth and development of plants. The leaf is a major sensory organ of plants. Light intensity can directly affect leaf development and morphogenesis. Supplementation of different light qualities affects the development of cucumber. In one study, cucumber under low-light condition were treated with red light and blue light. Red light could promote the expansion of leaf area, increase leaf chlorophyll content and photosynthetic rate, promote the accumulation of assimilates, and promote the distribution of assimilates to stems, roots, and other organs. Adding blue light could increase blade thickness and improve blade pore conductance [51]. At present, the effect of light on the respiration of cucumber leaves has been studied. Li et al. found that under the condition of LED supplementary light, the key enzymes of the glycolysis pathway decreased, whereas those of the tricarboxylic acid cycle and pentose phosphate pathway increased under the condition of 3 h/D LED supplementary light, which ensured the normal respiration of plants [52]. Continuous low light induced the SPAD (relative chlorophyll) value of cucumber leaves to increase and the net photosynthetic rate to decrease. The longer the duration of low light, the smaller the net photosynthetic rate, and the worse the photosynthetic recovery ability of cucumber leaves. Under low-light stress, the activities of superoxide dismutase and peroxidase in cucumber leaves increased, the activity of catalase increased first and then decreased, and the content of MDA increased continuously. The metabolic function of cucumber leaves could not recover to the normal level after more than 7 days of low-light stress [53]. The effect of light time on the leaf structure and photosynthetic characteristics of cucumber is noteworthy. With the increase in light time, the leaf thickness increased, that is, the content of accumulated organic matter, the thickness of palisade and sponge tissues, the number of stomata, and the content of chlorophyll in leaves increased [54] (Figure 2b).
3.3. Influence of CO2
The stomata of leaves are the main channel for the exchange of external gases between plants and the outside. The number and stomatal opening of leaves are determined by the concentration of external CO2 [55]. Plants can reduce the absorption of CO2 by closing stomata when the CO2 concentration increases in the environment, and the stomatal conductance of leaves treated with high concentration of CO2 was obviously lower than that of leaves treated with CO2 concentration under natural conditions [56]. Li revealed that the increase in CO2 concentration made the sugar content and sucrose synthase activity in cucumber leaves significantly increase. Under the same CO2 concentration, except for the leaves treated with high nitrogen under drought stress, the total nitrogen content of leaves under normal irrigation and high nitrogen treatment was significantly higher than under other treatments. Therefore, CO2 also increased the activities of key enzymes in the nitrogen metabolism in cucumber leaves. When CO2 enrichment increased the crop photosynthetic rate, the structure of leaves can be adjusted to adapt to the high concentration of CO2 [57]. Moreover, CO2 enrichment affected the microstructure and sub-microstructure of cucumber leaves. The results illustrated that the thickness of palisade tissue, epidermis, and cucumber leaves increased. The palisade cells became longer and were arranged more neatly and tightly after CO2 enrichment. The amount and size of chloroplasts, the number and size of starch grains, grana, and its lamellae, and the starch content per unit of chloroplast increased. The CO2 enrichment promoted the development of cucumber leaves and chloroplasts, increased the photosynthetic rate of cucumber leaves, and produced and accumulated nutrients needed for plant growth and development [58] (Figure 2c).
4. Prospect
Cucurbitaceae plants play an important role in vegetable production, and they are commonly used in daily life. As the main photosynthetic organ, the development of leaves is also directly related to the yield of Cucurbitaceae crops. Some leaf mutations, such as round leaves, leaflets, and downward curled leaves of cucumber, not only change the leaf shape but also affect other agronomic or economic characteristics compared with the WT. Whether due to multiple effects or closely linked traits, the relevant reasons depend on the in-depth research of functional genomics. Leaf development is an extremely complex process. Leaves of different species have their own peculiarities. Although cucumbers, melons, watermelons, and pumpkins belong to the Cucurbitaceae family, the shape of their leaves differs. Because the leaves are regulated by genes, hormones, and the external environment, the leaves of the same species have a certain variability. We have learned that cucurbits are susceptible to biotic stresses, of which pest and disease stresses are more serious. Common diseases of Cucurbitaceae include powdery mildew, downy mildew, wilt, etc. These pathogens mainly attack the leaves, which might affect the photosynthesis of the leaves or even make the leaves die. Insect pests mainly include aphid, whitefly, yellow melon lea beetle, etc. The pests that live on the leaves seriously affect the development of the leaves, mainly including aphids, whitefly, and Aulcophora indica, etc. For example, aphids can cause deformation in the form of crumpled leaves. Since leaves are the main object of disease and pest infection, can changes in leaf morphology and structure lead to improvements in plant resistance? Interestingly, some research results have verified this conjecture. The resistance of cucumber leaf roll mutants to powdery mildew was enhanced. The resistance traits of the mutants to leaf roll and powdery mildew were controlled by the same dominant mutant gene PMrl. The sponge tissue of the mutant leaf thickened and became tightly arranged, and the palisade tissue became narrower. The upper and lower epidermis cells thickened, and the leaf wax content was significantly higher than that of the wild type [59]. Coincidentally, the resistance of downward curly leaf mutants to downy mildew was also increased. The increase in salicylic acid (SA) in the mutant may be the reason for its enhanced resistance to downy mildew. Digging resistance genes and analyzing their resistance mechanism are the basis and premise of cultivating new disease-resistant and insect-resistant varieties, which not only helps to accelerate the progress of disease-resistant and pest-resistant variety breeding, but also provides new ideas for pest control [15]. Leaf shape and leaf color mutants could be used as marker traits to quickly identify variety purity and save much time. Furthermore, research on its molecular mechanism could provide a reference for cultivating an ideal plant type and increasing yield. At present, the germplasm resources of leaf mutants of Cucurbitaceae are relatively few, and they have not been widely utilized. Although some leaf mutants have been located and cloned, the complex regulatory network behind them is not clear, and more research work is needed. Excavating the functional genes of important agronomic traits of Cucurbitaceae and analyzing their regulatory network, establishing a stable genetic transformation system, and promoting the close combination of molecular breeding and conventional breeding techniques will be the focus of the next work. With the development of molecular biology and bioinformatics, the research and application of leaf molecular breeding of Cucurbit plants will make new progress and breakthroughs.
Author Contributions
Y.L., H.W. and S.F. jointly conceived the review, conducted the literature review and wrote the manuscript. Y.L., H.X. and S.F. collected the literature. H.W. provided critical comments on the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Natural Science Foundation of Zhejiang province (LY21C150002), the National Natural Science Foundation of China (grant no. 32172595, 31801862, 31872105, 31972221, 32002048), the National College Students Innovation and EntrepreneurshipTraining Program in 2021 (202110341043).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rolnik, A.; Olas, B. Vegetables from Cucurbitaceae family and their products; positive effect on human health. Nutrition 2020, 78, 110788. [Google Scholar] [CrossRef] [PubMed]
- Tsukaya, H. Leaf shape: Genetic controls and environmental factors. Int. J. Dev. Biol. 2005, 49, 547–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mingyue, F.U.; Cheng, S.; Feng, X.U.; Chen, Z.; Liu, Z.; Zhang, W.; Zheng, J.; Wang, L. Advance in mechanism of plant leaf colour mutation. Not. Bot. Horti Agrobot. Cluj-Napoca 2021, 49, 12071. [Google Scholar]
- Moon, J.; Hake, S. How a leaf gets its shape. Curr. Opin. Plant Biol. 2011, 14, 24–30. [Google Scholar] [CrossRef]
- Dkhar, J.; Pareek, A. What determines a leaf’s shape? EvoDevo 2014, 5, 47. [Google Scholar] [CrossRef] [Green Version]
- Niu, H.; Liu, X.; Tong, C.; Wang, H.; Li, S.; Lu, L.; Pan, Y.; Zhang, X.; Weng, Y.; Li, Z. The WUSCHEL-related homeobox1 gene of cucumber regulates reproductive organ development. J. Exp. Bot. 2018, 69, 5373–5387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, L.; Yan, S.; Jiang, L.; Liu, M.; Zhang, J.; Zhao, J.; Zhao, W.; Han, Y.; Wang, Q.; Zhang, X. HANABA TARANU regulates the shoot apical meristem and leaf development in cucumber (Cucumis sativus L.). J. Exp. Bot. 2015, 66, 7075–7087. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Ning, X.; Wang, Y.; Wang, X.; Yan, W.; Zhang, Z.; Li, G. Fine mapping of a gene that confers palmately lobed leaf (pll) in melon (Cucumis melo L.). Euphytica 2014, 200, 337–347. [Google Scholar] [CrossRef]
- Chen, X. Fine Mapping and Cloning of Watermelon Split Leaf Gene ClLL1. Master’s Thesis, Northwest University of Agriculture and Forestry Science and Technology, Shaanxi, China, 2018. (In Chinese). [Google Scholar]
- Zhang, C.; Chen, F.; Zhao, Z.; Hu, L.; Liu, H.; Cheng, Z.; Weng, Y.; Chen, P.; Li, Y. Mutations in CsPID encoding a Ser/Thr protein kinase are responsible for round leaf shape in cucumber (Cucumis sativus L.). Tag.Theor. Appl. Genetics.Theor. Und Angew. Genet. 2018, 131, 1379–1389. [Google Scholar] [CrossRef]
- Liu, X.; Hao, N.; Li, H.; Ge, D.; Du, Y.; Liu, R.; Wen, C.; Li, Y.; Zhang, X.; Wu, T. PINOID is required for lateral organ morphogenesis and ovule development in cucumber. J. Exp. Bot. 2019, 70, 5715–5730. [Google Scholar] [CrossRef]
- Song, M.; Cheng, F.; Wang, J.; Wei, Q.; Fu, W.; Yu, X.; Li, J.; Chen, J.; Lou, Q. A leaf shape mutant provides insight into PINOID Serine/Threonine Kinase function in cucumber (Cucumis sativus L.). J Integr. Plant Biol. 2019, 61, 1000–1014. [Google Scholar] [CrossRef] [PubMed]
- Rong, F.; Chen, F.; Huang, L.; Zhang, J.; Zhang, C.; Hou, D.; Cheng, Z.; Weng, Y.; Chen, P.; Li, Y. A mutation in class III homeodomain-leucine zipper (HD-ZIP III) transcription factor results in curly leaf (cul) in cucumber (Cucumis sativus L.). Theor. Appl. Genet. 2018, 132, 113–123. [Google Scholar] [CrossRef]
- Gao, D.; Zhang, C.; Zhang, S.; Hu, B.; Wang, S.; Zhang, Z.; Huang, S. Mutation in a novel gene SMALL AND CORDATELEAF 1 affects leaf morphology in cucumber. J. Bot. Engl. Ed. 2017, 59, 6. [Google Scholar]
- Yang, L.; Liu, H.; Zhao, J.; Pan, Y.; Cheng, S.; Lietzow, C.D.; Wen, C.; Zhang, X.; Weng, Y. LITTLELEAF (LL) encodes a WD40 repeat domain-containing protein associated with organ size variation in cucumber. Plant J. 2018, 95, 834–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, S.; Ning, K.; Wang, Z.; Liu, X.; Zhong, Y.; Ding, L.; Zi, H.; Cheng, Z.; Li, X.; Shan, H.; et al. CsIVP functions in vasculature development and downy mildew resistance in cucumber. PLoS Biol. 2020, 18, e3000671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Niu, H.; Li, C.; Shen, G.; Liu, X.; Weng, Y.; Wu, T.; Li, Z. WUSCHEL-related homeobox1 (WOX1) regulates vein patterning and leaf size in Cucumis sativus. Hortic. Res. 2020, 7, 182. [Google Scholar] [CrossRef]
- Shang, C.; Liu, Q.; Wang, M.; Hou, L.; Zhao, C.; Wang, X.; Xia, H. Research Progress on Molecular Mechanisms of Leaf Color Variation in Plants. Shandong Agric. Sci. 2021, 53, 127–134. (In Chinese) [Google Scholar] [CrossRef]
- Miao, H.; Gu, X.; Zhang, S.; Wang, X. Research Progress of vegetable Leaf Color Mutants. Chin. Veg. 2007, 6, 39–42. (In Chinese) [Google Scholar]
- Wang, J.; Ma, S.; Shang, J.; Cheng, S. Leaf Colour Mutants in Cucurbits and Their Research Progress. Anhui Agric. Sci. 2011, 39, 16039–16040+16116. (In Chinese) [Google Scholar] [CrossRef]
- Hang, K.; Li, Y.; Zhu, W.; Wei, Y.; Njogu, M.K.; Lou, Q.; Li, J.; Chen, J. Fine Mapping and Transcriptome Analysis of Virescent Leaf Gene v-2 in Cucumber (Cucumis sativus L.). Front. Plant Sci. 2020, 11, 1458. [Google Scholar]
- Miao, H.; Zhang, S.; Wang, M.; Wang, Y.; Weng, Y.; Gu, X. Molecular sciences fine mapping of virescent leaf gene v-1 in cucumber (Cucumis sativus L.). Mol. Sci. 2016, 17, 1602. [Google Scholar]
- Pierce, L.K.; Wehner, T.C. Review of Genes and Linkage Groups in Cucumber. HortScience: A publication of the American Society for Horticultural Science. HortScience 1990, 25, 605–615. [Google Scholar] [CrossRef]
- Guo, Y.M.; Gu, X.F.; Zhang, C.Z.; Fang, X.J.; Zhang, S.P.; Xu, C.Q. Genetic mechanism of the cucumber leaf mutant. Acta Hortic. Sin. 2003, 30, 409–412. [Google Scholar]
- Miao, H.; Gu, X.; Zhang, S.; Wang, X.; Fang, Z.; Zhang, Z. Changes of the Photosynthetic Pigment and Differential Expression of the Correlated Genes in a Chlorophyll-Deficient Cucumber Mutant (Cucumis sativus L.). Chin. Agric. Sci. 2010, 43, 4027–4035. (In Chinese) [Google Scholar]
- Chen, Y.; Cui, L.; Cheng, J. Chlorophyll Biosynthesis and Metabolism Analyses of Reciprocal Interspecific Hybrid between Cucumis hystrix and C. sativus. Plant Physiol. J. 2013, 49, 452–456. (In Chinese) [Google Scholar] [CrossRef]
- Song, M.; Wei, Q.; Wang, J.; Fu, W.; Qin, X.; Lu, X.; Cheng, F.; Yang, K.; Zhang, L.; Yu, X.; et al. Fine Mapping of CsVYL, Conferring Virescent Leaf Through the Regulation of Chloroplast Development in Cucumber. Front. Plant Ence 2018, 9, 432. [Google Scholar] [CrossRef]
- Xiong, L.; Du, H.; Zhang, K.; Lv, D.; He, H.; Pan, J.; Cai, R.; Wang, G. A Mutation in CsYL2.1 Encoding a Plastid Isoform of Triose Phosphate Isomerase Leads to Yellow Leaf 2.1 (yl2.1) in Cucumber (Cucumis sativus L.). Int. J. Mol. Sci. 2020, 22, 322. [Google Scholar] [CrossRef]
- Niu, Y.; Li, Z. Characteristic and fine-mapping of albino cotyledons(al) mutant in cucumber(Cucumis sativus L.). J. Northwest AF Univ. (Nat. Sci. Ed.) 2021, 49, 88–95+122. (In Chinese) [Google Scholar] [CrossRef]
- Hu, L.; Zhao, Z.; Zhang, H.; Chen, F.; Zhang, C.; Rong, F.; Chen, P.; Li, Y. Photosynthetic characteristics of analysis of new leaf color mutant in cucumber. Acta Agric. Boreali-Occident. Sin. 2018, 27, 1622–1628. (In Chinese) [Google Scholar]
- Li, W.; Gao, B.; Yang, J.; Chen, P.; Li, Y. Physiological characteristics of analysis a new leaf color yellow mutant in cucumber. Acta Agric. Boreali-Occident. Sin. 2015, 24, 98–103. (In Chinese) [Google Scholar]
- Ma, S.; Zhang, L. Discovery of mutant strains carrying watermelon albino lethal gene. Chin. Watermelon Muskmelon 1990, 4, 22. (In Chinese) [Google Scholar] [CrossRef]
- Wang, D.; Yang, J.; Li, Q.; Wu, S.; He, C.; Ma, Y. Phenotypic characteristics and genetic analysis of pumpkin (Cucurbita moschata Duch.) silver leaf mutant 48a. Chin. Cucurbita 2020, 33, 12–16. (In Chinese) [Google Scholar] [CrossRef]
- Lai, Y.; Fu, Q.; Lv, J.; Zhou, M.; He, M.; Xu, W.; Wang, H.; Huang, Z. Analysis of Physiological Characteristics and Chloroplast Ultrastructure of A New Leaf Color Mutant in Melon. J. Sichuan Agric. Univ. 2018, 36, 372–379. (In Chinese) [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, J.; Xing, G.; Li, M.; Li, S. Integrating physiology, genetics, and transcriptome to decipher a new thermo-sensitive and light-sensitive virescent leaf gene mutant in cucumber. Front. Plant Sci. 2022, 13, 972620. [Google Scholar] [CrossRef]
- Hu, L.; Zhang, H.; Xie, C.; Wang, J.; Zhang, J.; Wang, H.; Weng, Y.; Chen, P.; Li, Y. A mutation in CsHD encoding a histidine and aspartic acid domain-containing protein leads to yellow young leaf-1 (yyl-1) in cucumber (Cucumis sativus L.). Plant Sci. 2020, 293, 110407. [Google Scholar] [CrossRef]
- Shwartz, I.; Levy, M.; Ori, N.; Bar, M. Hormones in tomato leaf development. Dev. Biol. 2016, 419, 132–142. [Google Scholar] [CrossRef]
- Xiong, Y.; Jiao, Y. The Diverse Roles of Auxin in Regulating Leaf Development. Plants 2019, 8, 243. [Google Scholar] [CrossRef] [Green Version]
- Pekker, I.; Alvarez, J.P.; Eshed, Y. Auxin Response Factors Mediate Arabidopsis Organ Asymmetry via Modulation of KANADI Activity. Plant Cell 2005, 17, 2899–2910. [Google Scholar] [CrossRef] [Green Version]
- Chitwood, D.H.; Headland, L.R.; Ranjan, A.; Martinez, C.C.; Braybrook, S.A.; Koenig, D.P.; Kuhlemeier, C.; Smith, R.S.; Sinha, N.R. leaf asymmetry as a developmental constraint imposed by auxin-dependent phyllotactic patterning oa. Plant Cell 2017, 24, 2318–2327. [Google Scholar] [CrossRef]
- Donner, T.J.; Scarpella, E. Auxin-transport-dependent leaf vein formation. Botany 2009, 87, 678–684. [Google Scholar] [CrossRef]
- Bangerth, K.-F. Basipetal auxin versus acropetal cytokinin transport, and their interaction with NO3 fertilisation in cotyledon senescence and sink:source relationships in cucumber (Cucumis sativus L.). Plant Biol. 2015, 17, 1135–1145. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, D.; Ding, F.; Wu, J.; Gan, D. Expression of CsaIAAs gene in cucumber induced by 6-BA treatment. J. Anhui Agric. Univ. 2014, 41, 260–264. (In Chinese) [Google Scholar] [CrossRef]
- Dong, S.; Wang, W.; Bo, K.; Miao, H.; Song, Z.; Wei, S.; Zhang, S.; Gu, X. Quantitative Trait Loci Mapping and Candidate Gene Analysis of Low Temperature Tolerance in Cucumber Seedlings. Front. Plant Sci. 2019, 10, 1620. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Wang, M.; Xie, J.; Xiong, X.; Hu, X.; Qin, Y. Advances in response and research methods of plant low temperature stress. Jiangsu Agric. Sci. 2019, 47, 31–36. (In Chinese) [Google Scholar] [CrossRef]
- Zhang, Z.; Xu, H.; Wang, S.; Liang, Y.; Zhang, D. Effects of low temperature stress on physiological indexes of cucumber seedlings. Jiangsu Agric. Sci. 2013, 41, 126–127. (In Chinese) [Google Scholar] [CrossRef]
- Guo, F.; Lu, Y.; Dai, Z. Cold Tolerance of the Cucumber in Differ ent Development Period. Hubei Agric. Sci. 2008, 544, 548. (In Chinese) [Google Scholar]
- Zhou, Q. Research Advances on Physiological Responses and Tolerant Mechanism to High Temperature Stress in Rice. Chin. Agric. Sci. Bull. 2015, 31, 249–258. (In Chinese) [Google Scholar]
- Xue, S.; Yang, Z.; Zhu, L.; Li, J. Effects of high temperature stress on senescence and endogenous hormone of cucumber during flowering period. Chin. J. Ecol. 2018, 37, 409–416. (In Chinese) [Google Scholar] [CrossRef]
- Cui, Q.; Wu, L.; Chen, F.; Feng, X. Effect of High Temperature Stress on Physiological and Biochemical Characteristics and Photosynthesis of Cucumissativus L. Seedlings. J. Yanan Univ. (Nat. Sci. Ed.) 2021, 40, 23–26+31. (In Chinese) [Google Scholar] [CrossRef]
- Ni, J.; Chen, X.; Chen, C.; Xu, Q. Effects of Supplemental Different Light Qualities on Growth, Photosynthesis, Biomass Partition and Early Yield of Greenhouse Cucumber. Sci. Agric. Sin. 2009, 42, 2615–2623. (In Chinese) [Google Scholar]
- Li, X.; Ban, T.; Ma, C.; Wang, Q.; Yang, H.; Lu, J. Effects of light supplement on respiratory enzymes in leaves of cucumber seedling. Guizhou Agric. Sci. 2021, 49, 111–115. (In Chinese) [Google Scholar]
- Xiong, Y.; Diao, J.; Xue, X.; Lu, X.; Zhang, J. Effects of Continuous Overcast Weather on Cucumber Growth and Antioxidant Enzyme Activities in Glasshouse. A Chin. J. Agrometeorol. 2017, 38, 537–547. (In Chinese) [Google Scholar]
- Qin, L. Effects of Light time on Leaf structure and photosynthetic characteristics of Cucumber. J. Chifeng Univ. (Nat. Sci. Ed.) 2016, 32, 25–27. (In Chinese) [Google Scholar] [CrossRef]
- Chen, H.; Dickinson, R.E.; Dai, Y.; Zhou, L. Sensitivity of simulated terrestrial carbon assimilation and canopy transpiration to different stomatal conductance and carbon assimilation schemes. Clim. Dyn. 2011, 36, 1037–1054. [Google Scholar] [CrossRef]
- Ge, T.; Dabuxi, T. Effects of CO2 enrichment and different potassium levels on stomatal state of cucumber. Jiangsu Agric. Sci. 2018, 46, 110–113. (In Chinese) [Google Scholar] [CrossRef]
- Li, M.; Dong, Y.; Cui, Q.; Zhang, W.; Ai, X.; Liu, B.; Li, Q. Effects of water-nitrogen coupling on the metabolites and key enzyme activities of carbon and nitrogen metabolism in cucumber leaves under doubled CO2 concentration. Plant Physiol. J. 2017, 53, 1717–1727. (In Chinese) [Google Scholar] [CrossRef]
- Wei, M.; Xing, Y.; Wang, X.; Ma, H. Effects of CO2 Enrichment on the Microstructure and Ultrastructure of Leaves in Cucumber. Acta Hortic. Sin. 2002, 29, 30–34. (In Chinese) [Google Scholar]
- Rong, F.; Qin, Y.; Chen, F.; Zhang, C.; Zhao, Z.; Hu, L.; Li, Y. The leaf structure in cucumber slightly curly leaf mutant of resistance to powdery mildew. China Cucurbits Veg. 2017, 30, 11–14. (In Chinese) [Google Scholar] [CrossRef]
Figure 1.
Leaves of several leaf-shaped mutants of Cucurbitaceae.

Figure 2.
Environmental factors affecting leaf development as follows: (a) effect of temperature (blue, low temperature; red, high temperature); (b) effect of light—with the increase in leaf thickness, the thickness of palisade tissue and sponge tissue increased, the number of stomata increased, and the content of chlorophyll also increased; (c) influence of CO2—after CO2 enrichment, the thickness of the palisade tissue, epidermis, and cucumber leaves increased, and the number and size of chloroplasts, starch grains, grana, and their lamellae increased.
Figure 2.
Environmental factors affecting leaf development as follows: (a) effect of temperature (blue, low temperature; red, high temperature); (b) effect of light—with the increase in leaf thickness, the thickness of palisade tissue and sponge tissue increased, the number of stomata increased, and the content of chlorophyll also increased; (c) influence of CO2—after CO2 enrichment, the thickness of the palisade tissue, epidermis, and cucumber leaves increased, and the number and size of chloroplasts, starch grains, grana, and their lamellae increased.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, Y.; Xu, H.; Wang, H.; Feng, S. Research Progress in Leaf Related Molecular Breeding of Cucurbitaceae. Agronomy 2022, 12, 2908. https://doi.org/10.3390/agronomy12112908
AMA Style
Liu Y, Xu H, Wang H, Feng S. Research Progress in Leaf Related Molecular Breeding of Cucurbitaceae. Agronomy. 2022; 12(11):2908. https://doi.org/10.3390/agronomy12112908
Chicago/Turabian StyleLiu, Yafei, Huinan Xu, Huasen Wang, and Shengjun Feng. 2022. "Research Progress in Leaf Related Molecular Breeding of Cucurbitaceae" Agronomy 12, no. 11: 2908. https://doi.org/10.3390/agronomy12112908
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.