
Current Advances in the Action Mechanisms of Safeners


Abstract
1. Introduction
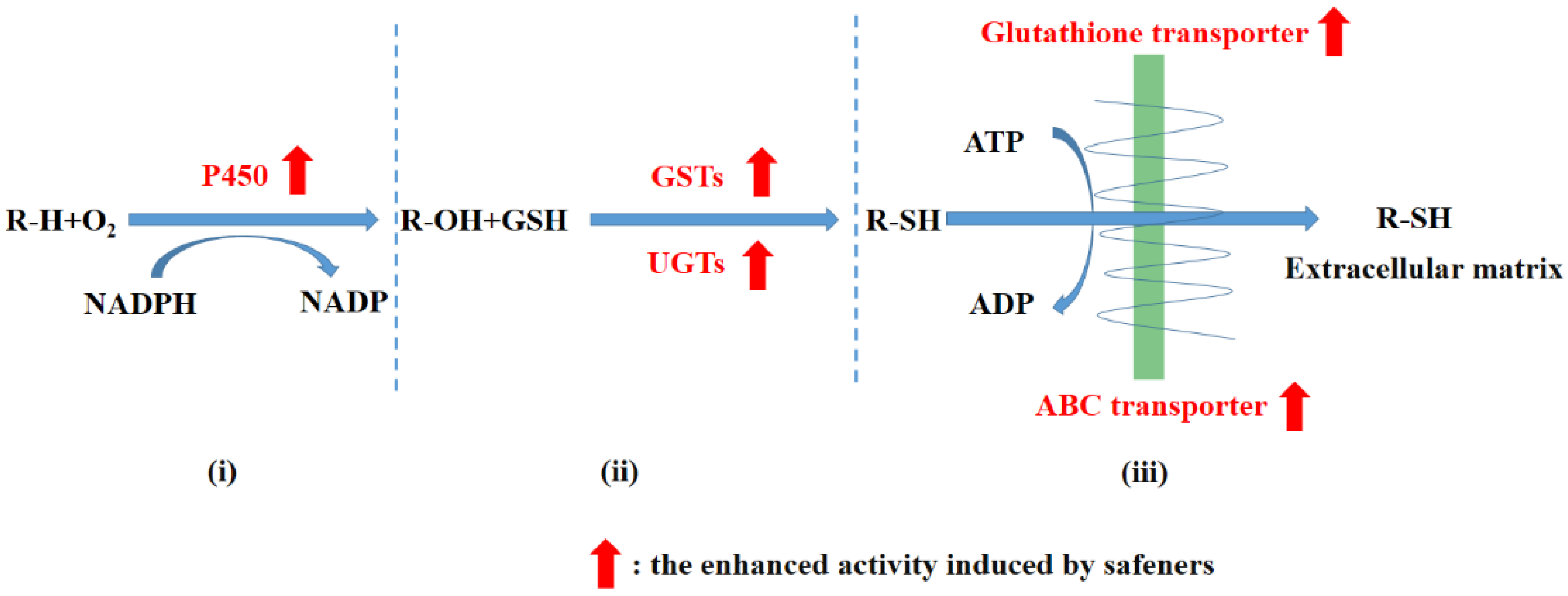
2. Enhancing the Herbicide Metabolism in Crop Plants
2.1. Induction of GSTs
2.2. Induction of UGTs
2.3. Induction of P450s
2.4. Induction of ABC Transporters and Glutathione Transporters
3. Effect of Safeners on Herbicide Absorption and Transport
4. Target Site Competition (Structure–Activity Theory)
5. Effecting the Target Enzyme Activity
6. Induction of Signaling Pathway
7. Effects of Safeners on Weeds
8. Conclusions and Perspectives
Funding
Data Availability Statement
Conflicts of Interest
References
- Deng, X.L.; Zhao, P.Y.; Zhou, X.M.; Bai, L.Y. Excellent sustained-release efficacy of herbicide quinclorac with cationic covalent organic frameworks. Chem. Eng. J. 2021, 405, 126979. [Google Scholar] [CrossRef]
- Guo, W.L.; Yu, C.J.; Zhang, C.; Zhang, T.J.; Tian, X.S. Multiple resistance detection to glyphosate and other herbicides in Conyza sumatrensis and the evaluation of chemical control herbicides. Chin. J. Pestic. Sci. 2022, 24, 789–797. [Google Scholar]
- Hossen, K.; Das, K.R.; Asato, Y.; Teruya, T.; Kato-Noguchi, H. Characterization of allelopathic substances from Elaeocarpus floribundus blume leaves for the development of bioherbicides. Agronomy 2022, 12, 57. [Google Scholar] [CrossRef]
- Deng, X.L.; Zhou, Y.; Zheng, W.N.; Bai, L.Y.; Zhou, X.M. Dissipation dynamic and final residues of oxadiargyl in paddy fields using high-performance liquid chromatography-tandem mass spectrometry coupled with modified QuEChERS Method. Int. J. Environ. Res. Public Health 2018, 15, 1680. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.Q.; Liu, X.D.; Napier, R.; Dong, L.Y.; Li, J. Mode of action of a novel synthetic auxin herbicide halauxifen-methyl. Agronomy 2022, 12, 1659. [Google Scholar] [CrossRef]
- Deng, X.L.; Zheng, W.N.; Zhou, X.M.; Bai, L.Y. The effect of salicylic acid and 20 substituted molecules on alleviating metolachlor herbicide injury in rice (Oryza sativa). Agronomy 2020, 10, 317. [Google Scholar] [CrossRef]
- Zheng, W.N.; Zhu, Z.Y.; Deng, Y.N.; Wu, Z.C.; Zhou, Y.; Zhou, X.M.; Bai, L.Y.; Deng, X.L. Synthesis, crystal structure, herbicide safening, and antifungal activity of N-(4,6-dichloropyrimidine-2-yl)benzamide. Crystals 2018, 8, 75. [Google Scholar] [CrossRef]
- Deng, X.L.; Zheng, W.N.; Jin, C.; Bai, L.Y. Synthesis of novel 6-aryloxy-4-chloro-2-phenylpyrimidines as fungicides and herbicide safeners. ACS Omega 2020, 5, 23996–24004. [Google Scholar] [CrossRef]
- Jia, L.; Jin, X.Y.; Zhao, L.X.; Fu, Y.; Ye, F. Research progress in the design and synthesis of herbicide safeners: A review. J. Agric. Food Chem. 2022, 70, 5499–5515. [Google Scholar] [CrossRef]
- Lin, C.E.; Wang, W.; Gao, X.L. Effects of the application of herbicides mixed with safeners on weeds control efficacy and the safety to proso mill. Chin. J. Pestic. Sci. 2022, 24, 352–360. [Google Scholar]
- Rosinger, C. Herbicide safeners: An overview. Julius Kühn Arch 2014, 443, 516–525. [Google Scholar]
- Marrs, K.A. The functions and regulation of glutathione s-transferases in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 127–158. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.; Choi, J.; Kim, J.; Cho, K.; Chun, J. Safening mode of action of 1, 8-naphthalic anhydride on corn and soybean against herbicide bensulfuron and imazaquin. Korean J. Weed Sci. 1994, 14, 8–15. [Google Scholar]
- Afelumo, O. The Effects of Herbicide Safener Naphthalic Anhydride on Nickel Toxicity in Corn. Master’s Thesis, State University of New York, New York, NY, USA, 2015. ProQuest. Available online: https://www.proquest.com/openview/b31d4667339ea16de2c2051dcf122a73/1?pq-origsite=gscholar&cbl=18750 (accessed on 22 September 2022).
- Davies, J.; Caseley, J.C. Herbicide safeners: A review. Pestic. Sci. 1999, 55, 1043–1058. [Google Scholar] [CrossRef]
- Hatzios, K.K.; Burgos, N. Metabolism-based herbicide resistance: Regulation by safeners. Weed Sci. 2017, 52, 454–467. [Google Scholar] [CrossRef]
- Zhao, L.X.; Qu, H.T.; Fu, Y.; Gao, S.; Ye, F. Alleviation of injury from chlorimuron-ethyl in maize treated with safener 3-dichloroacetyl oxazolidine. Can. J. Plant Sci. 2015, 95, 897–903. [Google Scholar] [CrossRef]
- Zhao, L.X.; Wang, Q.R.; Ying, F.; Li, C.Y.; Shuang, G.; Fei, Y. Induction of herbicide detoxifying enzyme in maize by chiral 3-dichloroacetyl oxazolidine. J. Agric. Sci. 2018, 24, 359–365. [Google Scholar] [CrossRef]
- Holt, D.C.; Lay, V.J.; Clarke, E.D.; Dinsmore, A.; Jepson, I.; Bright, S.W.; Greenland, A.J. Characterization of the safener-induced glutathione S-transferase isoform II from maize. Planta 1995, 196, 295–302. [Google Scholar] [CrossRef]
- Zhao, L.X.; Yin, M.L.; Wang, Q.R.; Zou, Y.L.; Ren, T.; Gao, S.; Fu, Y.; Ye, F. Novel thiazole phenoxypyridine derivatives protect maize from residual pesticide injury caused by PPO-inhibitor fomesafen. Biomolecules 2019, 9, 514. [Google Scholar] [CrossRef]
- Zhao, L.X.; Fu, Y.; Zhi-Yong, S.G.; Wei, X.L.N.; Ye, F. Protective responses induced by 3-dichloroacetyl oxazolidine safeners in maize (Zea mays). Int. J. Agric. Biol. 2014, 16, 1204–1208. [Google Scholar]
- Fuerst, E.P.; Irzyk, G.P.; Miller, K.D. Partial characterization of glutathione S-transferase isozymes induced by the herbicide safener benoxacor in maize. Plant Physiol. 1993, 102, 795–802. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.H.; Deng, X.L.; Bai, L.Y. Developmental toxicity and transcriptome analysis of zebrafish (Danio rerio) embryos following exposure to chiral herbicide safener benoxacor. Sci. Total Environ. 2021, 761, 143273. [Google Scholar] [CrossRef] [PubMed]
- Dutka, F.; Komives, T. Pesticide Science and Biotechnology; Blackwell Scientific Publications: Ottawa, ON, Canada, 1986; pp. 201–204. [Google Scholar]
- Islam, S.; Rahman, I.A.; Islam, T.; Ghosh, A. Genome-wide identification and expression analysis of glutathione S-transferase gene family in tomato: Gaining an insight to their physiological and stress-specific roles. PLoS ONE 2017, 12, e0187504. [Google Scholar] [CrossRef] [PubMed]
- DeRidder, B.P.; Dixon, D.P.; Beussman, D.J.; Edwards, R.; Goldsbrough, P.B. Induction of glutathione S-transferases in Arabidopsis by herbicide safeners. Plant Physiol. 2002, 130, 1497–1505. [Google Scholar] [CrossRef]
- Riechers, D.E.; Yang, K.; Irzyk, G.P.; Jones, S.S.; Fuerst, E.P. Variability of glutathioneS-transferase levels and dimethenamid tolerance in safener-treated wheat and wheat relatives. Pestic. Biochem. Physiol. 1996, 56, 88–101. [Google Scholar] [CrossRef]
- Ekler, Z.; Dutka, F.; Stephenson, G.R. Safener effects on acetochlor toxicity, uptake, metabolism and glutathione S-transferase activity in maize. Weed Res. 1993, 33, 311–318. [Google Scholar] [CrossRef]
- Deng, F.; Hatzios, K.K. Purification and characterization of two glutathione S-transferase isozymes from indica-type rice involved in herbicide detoxification. Pestic. Biochem. Physiol. 2002, 72, 10–23. [Google Scholar] [CrossRef]
- Wu, J.; Omokawa, H.; Hatzios, K.K. Glutathiones-S-transferase activity in unsafened and fenclorim-safened rice (Oryza sativa). Pestic. Biochem. Physiol. 1996, 54, 220–229. [Google Scholar] [CrossRef]
- Cummins, I.; Cole, D.J.; Edwards, R. Purification of multiple glutathione transferases involved in herbicide detoxification from wheat (Triticum aestivum L.) treated with the safener fenchlorazole-ethyl. Pestic. Biochem. Physiol. 1997, 59, 35–49. [Google Scholar] [CrossRef]
- Sun, L.; Wu, R.; Su, W.; Gao, Z.; Lu, C. Physiological basis for isoxadifen-ethyl induction of nicosulfuron detoxification in maize hybrids. PLoS ONE 2017, 12, e0173502. [Google Scholar] [CrossRef][Green Version]
- Bianchi, L.; Perissato, S.M.; Anunciato, V.M.; Dias, R.C.; Gomes, D.M.; Carbonari, C.A.; Velini, E.D. Stimulation action of mefenpyr-diethyl on soybean, wheat, and signal grass plants. J. Environ. Sci. Health Part B 2021, 56, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Brazier, M.; Cole, D.J.; Edwards, R. O-glucosyltransferase activities toward phenolic natural products and xenobiotics in wheat and herbicide-resistant and herbicide-susceptible black-grass (Alopecurus myosuroides). Phytochemistry 2002, 59, 149–156. [Google Scholar] [CrossRef]
- Giannakopoulos, G.; Dittgen, J.; Schulte, W.; Zoellner, P.; Helmke, H.; Lagojda, A.; Edwards, R. Safening activity and metabolism of the safener cyprosulfamide in maize and wheat. Pestic. Manag. Sci. 2020, 76, 3413–3422. [Google Scholar] [CrossRef] [PubMed]
- Giraldo Acosta, M.; Cano, A.; Hernández-Ruiz, J.; Arnao, M.B. Melatonin as a possible natural safener in crops. Plants 2022, 11, 890. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Deng, X.L.; Zhou, S.F.; Wang, C.; Yang, L.; Tang, Y.; Liu, S.H.; Yang, H.N.; Wu, L.M.; Wang, L.F.; et al. The effect and mechanism of gibberellin alleviate the phytotoxicity of S-metolachlor on rice seedlings. Agrochem 2019, 58, 519–522. [Google Scholar]
- Tang, X.; Zhou, X.; Wu, J.; Li, J.; Bai, L. A novel function of sanshools: The alleviation of injury from metolachlor in rice seedlings. Pestic. Biochem. Physiol. 2014, 110, 44–49. [Google Scholar] [CrossRef]
- Hu, L.F.; Wang, L.F.; Zhou, X.M.; Luo, K.; Bai, L.Y. Two coumarins with safener activity from Rhizoma et Radix Notopterygii. Weed Technol. 2017, 29, 161–167. [Google Scholar] [CrossRef]
- Li, J.B.; Zheng, W.; Wang, Y.H.; Cai, H.L.; Jin, C.Z.; Liu, X.B.; Bai, L.Y. Identification and S-metolachlor-safening effects of compounds extracted from Ligusticum chuanxiong on rice. Int. J. Agric. Biol. 2016, 18, 698–702. [Google Scholar] [CrossRef]
- Zhou, X.M.; Bai, L.Y.; Liu, X.Y.; Ren, X.G.; Li, S.Q.; Bu, X.L.; Huang, K.C. Studies on the mechanism of activity of brassinolide for reducing the phytotoxicity of ethametsulfuron on rice. Chin. J. Pestic. Sci. 2003, 5, 61–67. [Google Scholar]
- Deng, X.L.; Zheng, W.N.; Jin, C.; Zhan, Q.C.; Bai, L.Y. Novel phenylpyrimidine derivatives containing a hydrazone moiety protect rice seedlings from injury by metolachlor. Bioorgan. Chem. 2021, 108, 104645. [Google Scholar] [CrossRef]
- Deng, X.L.; Zheng, W.N.; Zhan, Q.C.; Deng, Y.N.; Zhou, Y.; Bai, L.Y. New Lead discovery of herbicide safener for metolachlor based on a scaffold-hopping strategy. Molecules 2020, 25, 4986. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Y.; Gao, S.; Liu, Y.X.; Wang, C.; Jiang, W.; Zhao, L.X.; Fu, Y.; Ye, F. Design, synthesis, and biological activity of novel diazabicyclo derivatives as safeners. J. Agric. Food Chem. 2020, 68, 3403–3414. [Google Scholar] [CrossRef] [PubMed]
- Kang, T.; Gao, S.; Zhao, L.X.; Zhai, Y.; Ye, F.; Fu, Y. Design, synthesis, and SAR of novel 1,3-disubstituted imidazolidine or hexahydropyrimidine derivatives as herbicide safeners. J. Agric. Food Chem. 2021, 69, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Guo, K.L.; Zhao, L.X.; Wang, Z.W.; Gao, Y.C.; Li, J.J.; Gao, S.; Fu, Y.; Ye, F. Design, synthesis, and bioevaluation of substituted phenyl isoxazole analogues as herbicide safeners. J. Agric. Food Chem. 2020, 68, 10550–10559. [Google Scholar] [CrossRef]
- Jia, L.; Zhao, L.X.; Sun, F.; Peng, J.; Wang, J.Y.; Leng, X.Y.; Gao, S.; Fu, Y.; Ye, F. Diazabicyclo derivatives as safeners protect cotton from injury caused by flumioxazin. Pestic. Biochem. Physiol. 2022, 187, 105185. [Google Scholar] [CrossRef]
- Jia, L.; Gao, S.; Zhang, Y.Y.; Zhao, L.X.; Fu, Y.; Ye, F. Fragmenlt recombination design, synthesis, and safener activity of novel ester-substituted pyrazole derivatives. J. Agric Food Chem 2021, 69, 8366–8379. [Google Scholar] [CrossRef]
- Fu, Y.; Zhang, D.; Kang, T.; Guo, Y.Y.; Chen, W.G.; Gao, S.; Ye, F. Fragment splicing-based design, synthesis and safener activity of novel substituted phenyl oxazole derivatives. Bioorgan. Med. Chem. Lett. 2019, 29, 570–576. [Google Scholar] [CrossRef]
- Wang, Z.W.; Zhao, L.X.; Ma, P.; Ye, T.; Fu, Y.; Ye, F. Fragments recombination, design, synthesis, safener activity and CoMFA model of novel substituted dichloroacetylphenyl sulfonamide derivatives. Pest. Manag. Sci. 2021, 77, 1724–1738. [Google Scholar] [CrossRef]
- Wang, Z.W.; Zhao, L.X.; Gao, S.; Leng, X.Y.; Yu, Y.; Fu, Y.; Ye, F. Quinoxaline derivatives as herbicide safeners by improving Zea mays tolerance. Pestic. Biochem. Physiol. 2021, 179, 104958. [Google Scholar] [CrossRef]
- Ye, F.; Zhai, Y.; Kang, T.; Wu, S.L.; Li, J.J.; Gao, S.; Zhao, L.X.; Fu, Y. Rational design, synthesis and structure-activity relationship of novel substituted oxazole isoxazole carboxamides as herbicide safener. Pestic. Biochem. Physiol. 2019, 157, 60–68. [Google Scholar] [CrossRef]
- Scarponi, L.; Quagliarini, E.; Del Buono, D. Induction of wheat and maize glutathione S-transferase by some herbicide safeners and their effect on enzyme activity against butachlor and terbuthylazine. Pest. Manag. Sci. 2006, 62, 927–932. [Google Scholar] [CrossRef] [PubMed]
- Deng, F.; Hatzios, K.K. Characterization and safener induction of multiple glutathione S-transferases in three genetic lines of rice. Pestic. Biochem. Physiol. 2002, 72, 24–39. [Google Scholar] [CrossRef]
- Ye, F.; Zhai, Y.; Guo, K.L.; Liu, Y.X.; Li, N.; Gao, S.; Zhao, L.X.; Fu, Y. Safeners improve maize tolerance under herbicide toxicity stress by increasing the activity of enzymes in vivo. J. Agric. Food. Chem. 2019, 67, 11568–11576. [Google Scholar] [CrossRef] [PubMed]
- Ye, F.; Cao, H.F.; Chen, X.S.; Zhang, M.; FU, Y.; Li, C.Y.; Gao, S. Effects of chiral 3-dichloroacetyl oxazolidine on glutathione S-transferase and antioxidant enzymes activity in maize treated with acetochlor. Tarım Bilim. Derg. J. Agric. Sci. 2018, 24, 422–429. [Google Scholar] [CrossRef]
- Kreuz, K.; Gaudin, J.; Stingelin, J.; Ebert, E. Metabolism of the aryloxyphenoxypropanoate herbicide, cga 184927, in wheat, barley and maize: Differential effects of the safener, CGA 185072. Pestic. Biochem. Physiol. 1991, 46, 901–905. [Google Scholar] [CrossRef]
- Edwards, R.; Buono, D.D.; Fordham, M.; Skipsey, M.; Brazier, M.; Dixon, D.P.; Cummins, I. Differential induction of glutathione transferases and glucosyltransferases in wheat, maize and Arabidopsis thaliana by herbicide safeners. Z. Naturforsch. C 2005, 60, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Riechers, D.E.; Kreuz, K.; Zhang, Q. Detoxification without intoxication: Herbicide safeners activate plant defense gene expression. Plant Physiol. 2010, 153, 3–13. [Google Scholar] [CrossRef]
- Kreuz, K.; Tommasini, R.; Martinoia, E. Old enzymes for a new job (Herbicide detoxification in plants). Plant Physiol. 1996, 111, 349–353. [Google Scholar] [CrossRef]
- Siminszky, B. Plant cytochrome P450-mediated herbicide metabolism. Phytochem. Rev. 2006, 5, 445–458. [Google Scholar] [CrossRef]
- Frear, D.S.; Swanson, H.R.; Tanaka, F.S. N-demethylation of substituted 3-(phenyl)-1-methylureas: Isolation and characterization of a microsomal mixed function oxidase from cotton. Phytochemistry 1969, 8, 2157–2169. [Google Scholar] [CrossRef]
- Leavitt, J.R.C.; Penner, D. In vitro conjugation of glutathione and other thiols with acetanilide herbicides and EPTC sulfoxide and the action of the herbicide antidote R-25788. J. Agric. Food. Chem. 1979, 27, 533–536. [Google Scholar] [CrossRef]
- Persans, M.W.; Schuler, M.A. Differential induction of cytochrome P450-mediated triasulfuron metabolism by naphthalic anhydride and triasulfuron. Plant Physiol. 1995, 109, 1483–1490. [Google Scholar] [CrossRef] [PubMed]
- Deng, F.; Hatzios, K.K. Characterization of cytochrome P450-mediated bensulfuron-methyl O-demethylation in rice. Pestic. Biochem. Physiol. 2002, 74, 102–115. [Google Scholar] [CrossRef]
- Liu, H. Spectral characterization and chiral interactions of plant microsomal cytochrome P450 with metolachlor and herbicide safeners. J. Environ. Sci. Health Part B 2009, 45, 33–39. [Google Scholar] [CrossRef]
- Yamada, T.; Kambara, Y.; Imaishi, H.; Ohkawa, H. Molecular cloning of novel cytochrome P450 species induced by chemical treatments in cultured tobacco cells. Pestic. Biochem. Physiol. 2000, 68, 11–25. [Google Scholar] [CrossRef]
- Forthoffer, N.; Helvig, C.; Dillon, N.; Benveniste, I.; Zimmerlin, A.; Tardif, F.; Salaün, J.P. Induction and inactivation of a cytochrome P450 confering herbicide resistance in wheat seedlings. Eur. J. Drug Metab. Pharmacokinet. 2001, 26, 9–16. [Google Scholar] [CrossRef]
- LIi, Y.J.; Ye, F. Effects of herbicide safeners on levels and activity of cytochrome P450 and other enzymes of crop. Chin. J. Pestic. Sci. 2003, 5, 9–15. [Google Scholar]
- Simarmata, M.; Penner, D. Protection from primisulfuron injury to corn (Zea mays) and sorghum (Sorghum bicolor) with herbicide safeners. Weed Technol. 1993, 5, 174–179. [Google Scholar] [CrossRef]
- Cseresnyes, I.; Fekete, G.; Vegh, K.R.; Szekacs, A.; Mortl, M.; Rajkai, K. Monitoring of herbicide effect in maize based on electrical measurements. Int. Agrophys. 2012, 26, 243–247. [Google Scholar] [CrossRef]
- Kreuz, K.; Fonné-Pfister, R. Herbicide-insecticide interaction in maize: Malathion inhibits cytochrome P450-dependent primisulfuron metabolism. Pestic. Biochem. Physiol. 1992, 43, 232–240. [Google Scholar] [CrossRef]
- Jablonkai, I.; Hatzios, K.K. Microsomal oxidation of the herbicides EPTC and acetochlor and of the safener MG-191 in maize. Pestic. Biochem. Physiol. 1992, 43, 98–240. [Google Scholar] [CrossRef]
- Polge, N.D.; Barrett, M. Characterization of cytochrome P450-mediated chlorimuron ethyl hydroxylation in maize microsomes. Pestic. Biochem. Physiol. 1992, 95, 193–204. [Google Scholar] [CrossRef]
- Moreland, D.E.; Corbin, F.T.; Mcfarland, J.E. Effects of safeners on the oxidation of multiple substrates by grain sorghum microsomes. Pestic. Biochem. Physiol. 1993, 45, 43–53. [Google Scholar] [CrossRef]
- Romano, M.L.; Stephenson, G.R.; Tal, A.; Hall, J.C. The effect of monooxygenase and glutathione S-transferase inhibitors on the metabolism of diclofop-methyl and fenoxaprop-ethyl in barley and wheat. Pestic. Biochem. Physiol. 1993, 46, 181–189. [Google Scholar] [CrossRef]
- Gao, Z.J.; Jiang, J.Z.; Li, X.F.; Qiu, L.H.; Wang, C.J.; Zhang, W.J.; Cheng, Y.C. Studies on the induction of wheat cytochrome P450 by mefenpyr--diethyl and the spectrum characteristics. Plant Prot. 2005, 31, 40–44. [Google Scholar]
- Ishikawa, T. The ATP-dependent glutathione S-conjugate export pump. Trends Biochem. Sci. 1992, 17, 463–468. [Google Scholar] [CrossRef]
- Klein, M.; Burla, B.; Martinoia, E. The multidrug resistance-associated protein (MRP/ABCC) subfamily of ATP-binding cassette transporters in plants. FEBS Lett. 2006, 580, 1112–1122. [Google Scholar] [CrossRef]
- Gaillard, C.; Dufaud, A.; Tommasini, R.; Kreuz, K.; Amrhein, N.; Martinoia, E. A herbicide antidote (safener) induces the activity of both the herbicide detoxifying enzyme and of a vacuolar transporter for the detoxified herbicide. FEBS Lett. 1994, 352, 219–221. [Google Scholar] [CrossRef]
- Martinoia, E.; Grill, E.; Tommasini, R.; Kreuz, K.; Amrhein, N. ATP-dependent glutathione S-conjugate ‘export’ pump in the vacuolar membrane of plants. Nature 1993, 364, 247–249. [Google Scholar] [CrossRef]
- Pang, S.; Ran, Z.; Liu, Z.; Song, X.; Duan, L.; Li, X.; Wang, C. Enantioselective induction of a glutathione-S-transferase, a glutathione transporter and an ABC transporter in maize by Metolachlor and its (S)-isomer. PLoS ONE 2012, 7, e48085. [Google Scholar] [CrossRef]
- Bartholomew, D.M.; Van Dyk, D.E.; Lau, S.M.C.; O’Keefe, D.P.; Rea, P.A.; Viitanen, P.V. Alternate energy-dependent pathways for the vacuolar uptake of glucose and glutathione conjugates. Plant Physiol. 2002, 130, 1562–1572. [Google Scholar] [CrossRef] [PubMed]
- Klein, M.; Weissenböck, G.; Dufaud, A.; Gaillard, C.; Kreuz, K.; Martinoia, E. Different energization mechanisms drive the vacuolar uptake of a flavonoid glucoside and a herbicide glucoside. J. Biol. Chem. 1996, 271, 29666–29671. [Google Scholar] [CrossRef] [PubMed]
- Theodoulou, F.L.; Clark, I.M.; He, X.L.; Pallett, K.E.; Cole, D.J.; Hallahan, D.L. Co-induction of glutathione-S-transferases and multidrug resistance associated protein by xenobiotics in wheat. Pest. Manag. Sci. 2003, 59, 202–214. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Xu, F.; Lambert, K.N.; Riechers, D.E. Safeners coordinately induce the expression of multiple proteins and MRP transcripts involved in herbicide metabolism and detoxification in Triticum tauschii seedling tissues. Proteomics 2007, 7, 1261–1278. [Google Scholar] [CrossRef]
- Pang, S.; Duan, L.; Liu, Z.; Song, X.; Li, X.; Wang, C. Co-induction of a glutathione-S-transferase, a glutathione transporter and an ABC transporter in maize by xenobiotics. PLoS ONE 2012, 7, e40712. [Google Scholar] [CrossRef]
- Pang, S.; Duan, L.; Liu, Z.; Song, X.; Li, X.; Wang, C. Metolachlor-induced ZmGT1 expression in maize cultivars is correlated with their tolerance to the herbicide. J. Food Agric. Environ. 2012, 10, 621–623. [Google Scholar]
- Han, S.; Hatzios, K.K. Uptake, translocation, and metabolism of [14C]pretilachlor in fenclorim-safened and unsafened rice seedlings. Pestic. Manag. Sci. 1991, 39, 281–290. [Google Scholar] [CrossRef]
- Scarponi, L.; Del Buono, D.; Vischetti, C. Effect of pretilachlor and fenclorim on carbohydrate and protein formation in relation to their persistence in rice. Pestic. Manag. Sci. 2005, 61, 371–376. [Google Scholar] [CrossRef]
- Bunting, J.A.; Sprague, C.L.; Riechers, D.E. Physiological basis for tolerance of corn hybrids to foramsulfuron. Weed Sci. 2017, 52, 711–717. [Google Scholar] [CrossRef]
- Yenne, S.P.; Hatzios, K.K. Molecular comparisons of selected herbicides and their safeners by computer-aided molecular modeling. J. Agric. Food. Chem. 1990, 38, 1950–1956. [Google Scholar] [CrossRef]
- Ezra, G.; Krochmal, E.; Gressel, J. Competition between a thiocarbamate herbicide and herbicide protectants at the level of uptake into maize cells in culture. Pestic. Biochem. Physiol. 1982, 18, 107–112. [Google Scholar] [CrossRef]
- Walton, J.D.; Casida, J.E. Specific binding of a dichloroacetamide herbicide safener in maize at a site that also binds thiocarbamate and chloroacetanilide herbicides. Plant Physiol. 1995, 109, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.J.; Chen, J.P.; Hu, J.Y.; Qian, C.F.; Li, Z.M. Activity of acetolactate synthase from maize (Zea mays L.) as influenced by chlorsulfuron and tribenuron-methyl. Agric. Sci. China 2003, 2, 176–182. [Google Scholar]
- Rubin, B.; Casida, J.E. R-25788 effects on chlorsulfuron injury and acetohydroxyacid synthase activity. Weed Sci. 2017, 33, 462–468. [Google Scholar] [CrossRef]
- Zhao, L.X.; Wu, H.; Fu, Y.; Zou, Y.L.; Ye, F. 3-dichloroacetyl oxazolidine protect maize from imazethapyr herbicide injury. Chil. J. Agric. Res. 2016, 76, 158–162. [Google Scholar] [CrossRef]
- Barrett, M. Reduction of imazaquin injury to corn (Zea mays) and sorghum (Sorghum bicolor) by antidotes. Weed Sci. 2017, 37, 34–41. [Google Scholar] [CrossRef]
- Yun, M.S.; Shim, I.S.; Usui, K. Differential activities of Odealkylation by cytochrome P-450 in rice and Cyperus serotinus. J. Weed Sci. Technol. 2000, 45, 39–42. [Google Scholar] [CrossRef][Green Version]
- Hu, L.F.; Huang, Y.J.; Ding, B.A.; Cai, R.W.; Bai, L.Y. Selective action mechanism of fenclorim on rice and echinochloa crusgalli is associated with the inducibility of detoxifying enzyme activities and antioxidative defense. J. Agric. Food. Chem. 2021, 69, 5830–5839. [Google Scholar] [CrossRef]
- Rosenhauer, M.; Rosinger, C.; Petersen, J. Impact of the safener mefenpyr-diethyl on herbicide resistance evolution in Alopecurus myosuroides (Huds.) biotypes. Julius-Kühn-Archiv. 2016, 452, 50–56. [Google Scholar]
- Cummins, I.; Bryant, D.N.; Edwards, R. Safener responsiveness and multiple herbicide resistance in the weed black-grass (Alopecurus myosuroides). Plant. Biotechnol. J. 2009, 7, 807–820. [Google Scholar] [CrossRef]


{kind=link}
| Commercial Safeners | References | Commercial Safeners | References |
|---|---|---|---|
| 1,8-naphthalic anhydride (NA) | [13,14] | dichlormid | [15,16] |
| R-28725 | [17,18] | R-29148 | [19] |
| furilazole | [20] | AD-67 | [21] |
| benoxacor | [22,23] | MG-191 | [24] |
| cyometrinil | [25] | oxabetrinil | [26] |
| fluoxfenim | [27] | acetamate | [28] |
| fenclorim | [29,30] | fenchlorazole-ethyl | [31] |
| isoxadifen-ethyl | [32] | mefenpyr-diethyl | [33] |
| cluquintocet-mexyl | [34] | cyprosulfamide | [35] |
| Natural safeners | References | Natural safeners | References |
| melatonin | [36] | gibberellin | [37] |
| sanshools | [38] | isopimpinellin | [39] |
| 5-methoxypsoralen | [39] | Z-ligustilide | [40] |
| brassinolide | [41] | ||
| Safener candidates | References | Safener candidates | References |
| (E)-4-(2-substituted hydrazinyl)-6-chloro-2-phenypyrimidines | [42] | N-alkyl amides | [43] |
| diazabicyclo derivatives | [44] | 1,3-disubstituted imidazolidine or hexahydropyrimidine derivatives | [45] |
| phenyl isoxazole analogues | [46] | diazabicyclo derivatives | [47] |
| ester-substituted pyrazole derivatives | [48] | substituted phenyl oxazole derivatives | [49] |
| substituted dichloroacetylphenyl sulfonamide derivatives | [50] | quinoxaline derivatives | [51] |
| substituted oxazole isoxazole carboxamides | [52] | N-tosyloxazolidine-3-carboxamide | [53] |
| Commercial Safeners | References | Commercial Safeners | References |
|---|---|---|---|
| dichlormid | [61] | NA | [62,63,64,65,66,67] |
| R-28725 | [68] | R-29148 | [69] |
| furilazole | [20] | AD-67 | [70] |
| benoxacor | [71] | MG-191 | [72] |
| cyometrinil | [73] | fluoxfenim | [74] |
| fenclorim | [69] | fenchlorazole-ethyl | [75] |
| isoxadifen-ethy | [76] | mefenpyr-diethyl | [32] |
| Natural safener | References | Safener candidates | References |
| melatonin | [39] | quinoxaline derivatives | [51] |
| diazabicyclo derivatives | [47] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, X. Current Advances in the Action Mechanisms of Safeners. Agronomy 2022, 12, 2824. https://doi.org/10.3390/agronomy12112824
Deng X. Current Advances in the Action Mechanisms of Safeners. Agronomy. 2022; 12(11):2824. https://doi.org/10.3390/agronomy12112824
Chicago/Turabian StyleDeng, Xile. 2022. "Current Advances in the Action Mechanisms of Safeners" Agronomy 12, no. 11: 2824. https://doi.org/10.3390/agronomy12112824
APA StyleDeng, X. (2022). Current Advances in the Action Mechanisms of Safeners. Agronomy, 12(11), 2824. https://doi.org/10.3390/agronomy12112824