
Impacts of Drought Stress on Water Use Efficiency and Grain Productivity of Rice and Utilization of Genotypic Variability to Combat Climate Change



 ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
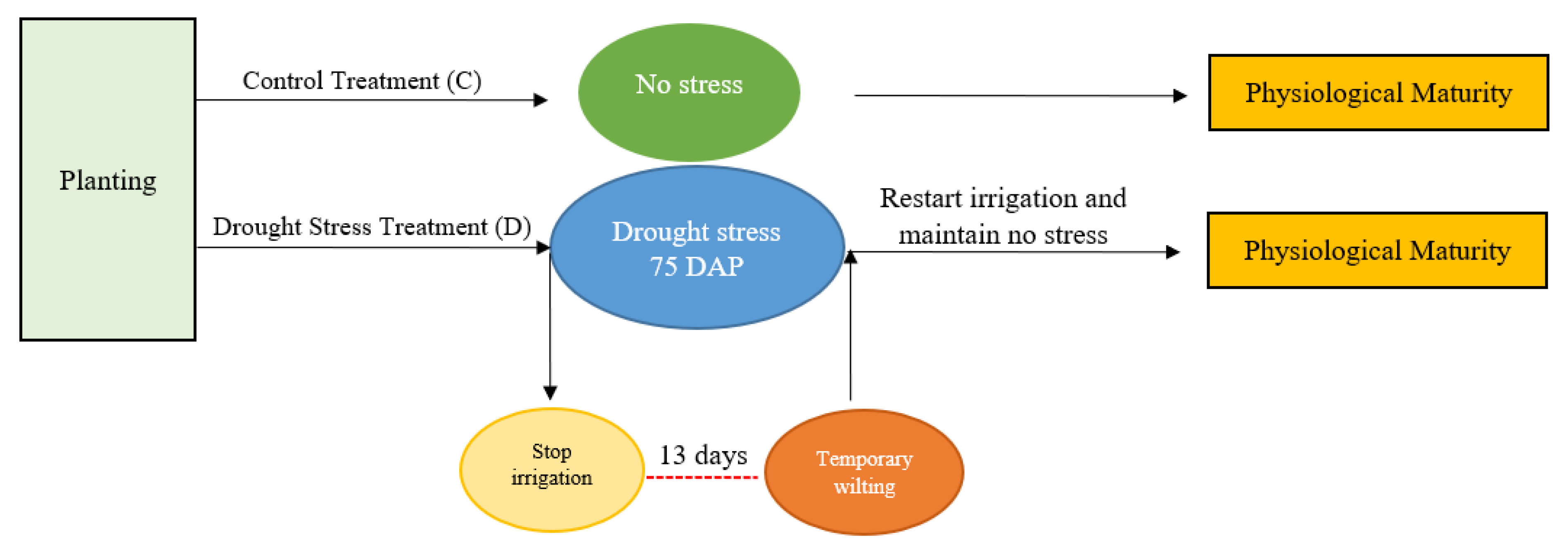
2.1. Experimental Setup and Crop Management
2.2. Crop Harvest and Data Collection
2.3. Determination of the Water Use Efficiency
2.4. Statistical Analysis
3. Results
3.1. Irrigation Water Consumption and Crop Duration
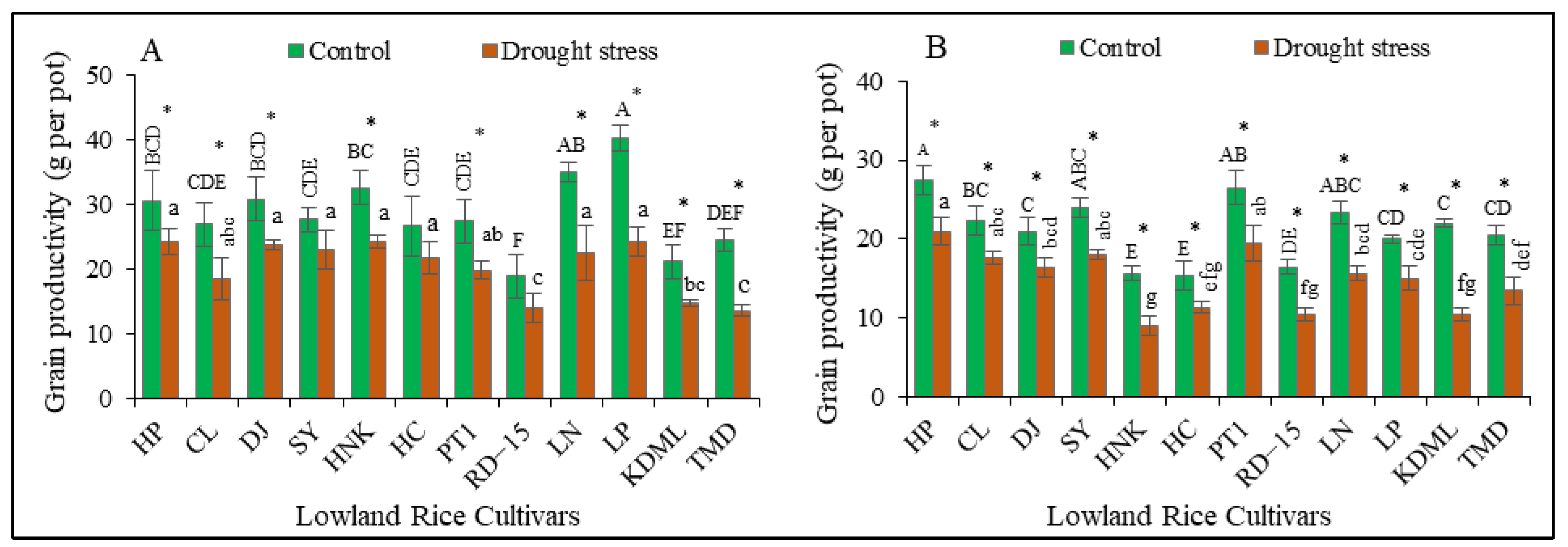
3.2. Impact of Drought Stress on Grain Productivity
3.3. Impact of Drought Stress on Water Use Efficiency
3.4. Genotypic Variability
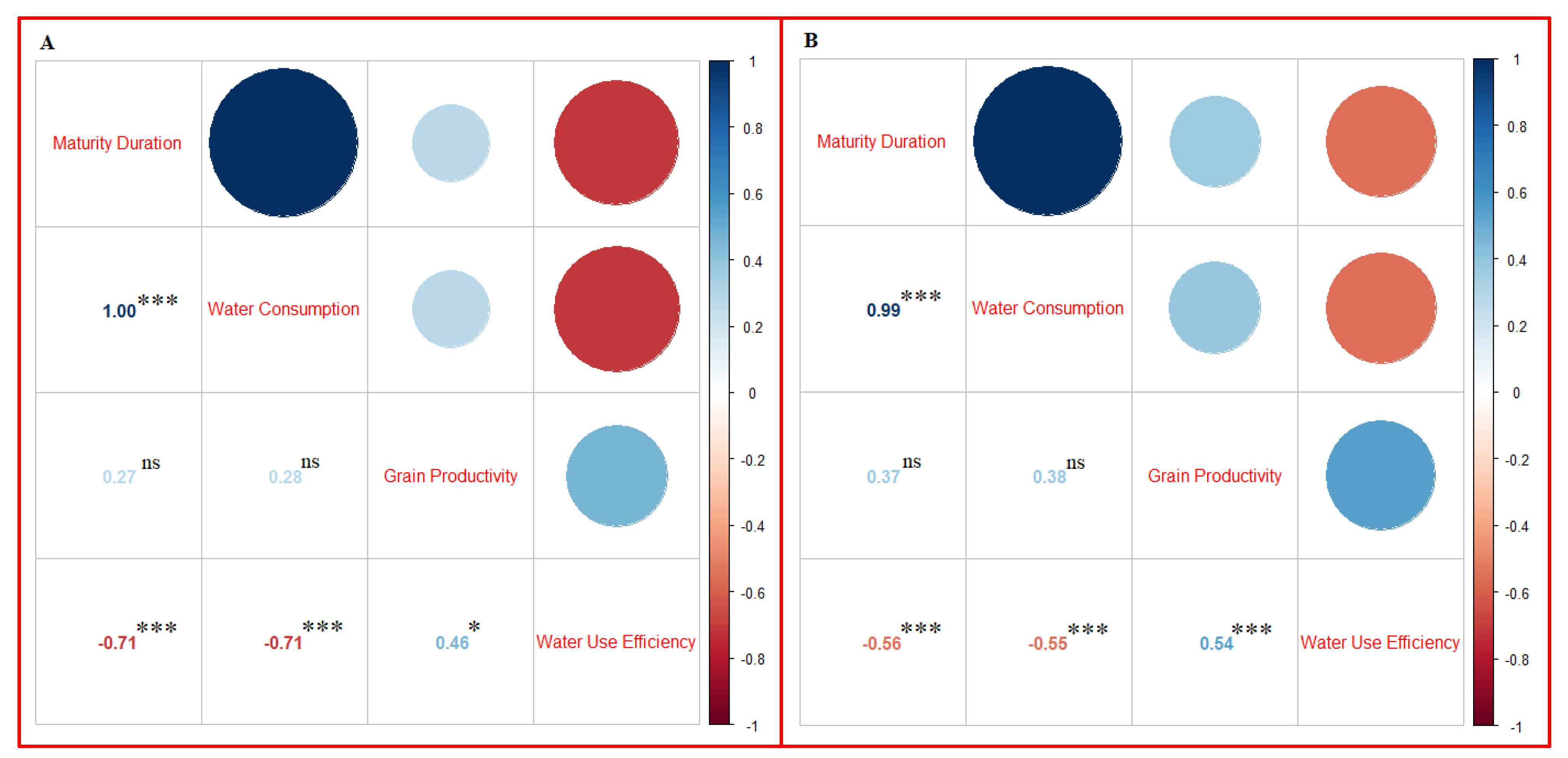
3.5. Correlation Study
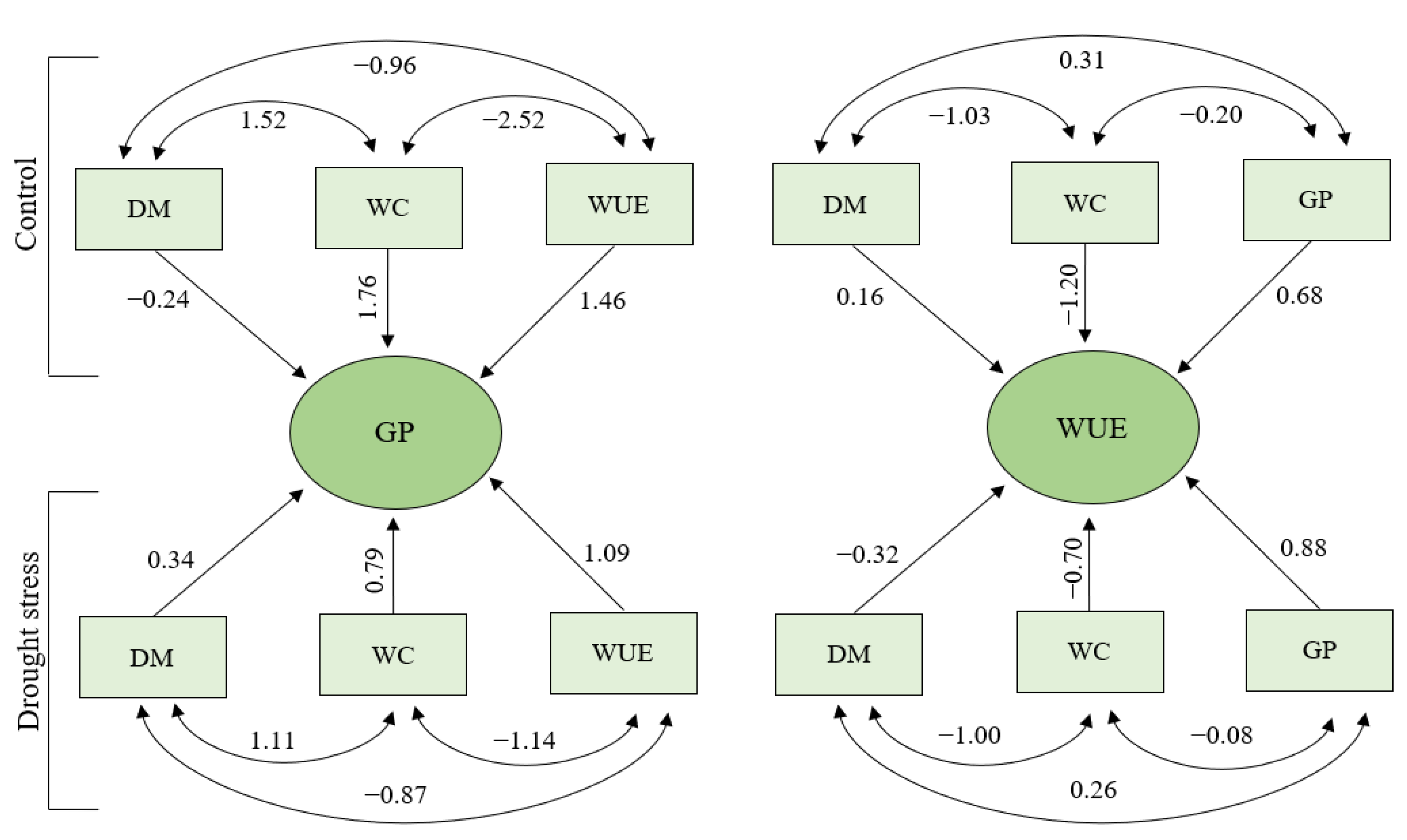
3.6. Path Analysis
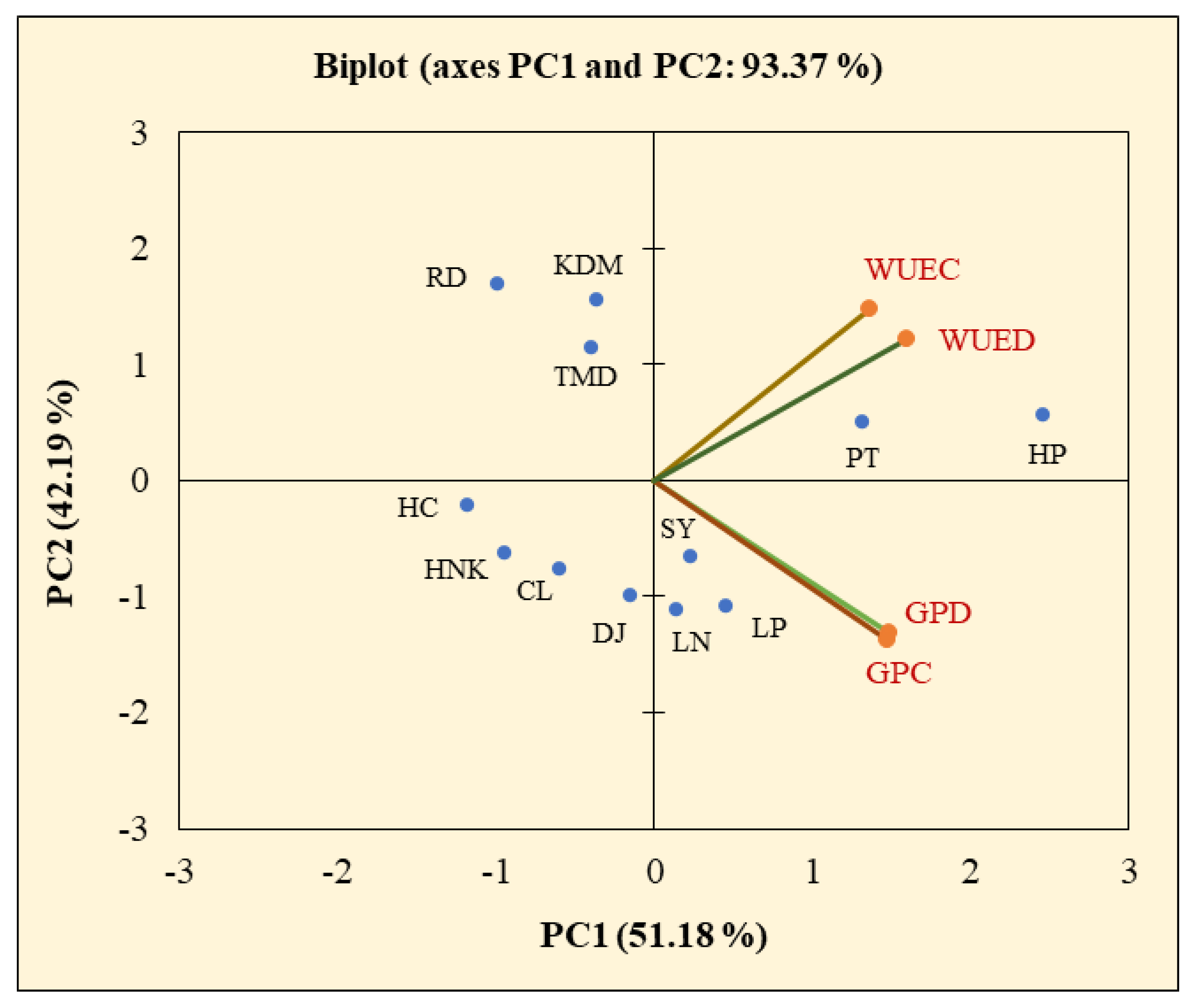
3.7. Classification of Cultivars Using Hierarchical Clustering and Principal Component Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Latif, M.; Bukhari, S.A.H.; Alrajhi, A.A.; Alotaibi, F.S.; Ahmad, M.; Shahzad, A.N.; Dewidar, A.Z.; Mattar, M.A. Inducing Drought Tolerance in Wheat through Exopolysaccharide-Producing Rhizobacteria. Agronomy 2022, 12, 1140. [Google Scholar] [CrossRef]
- USDA Thailand: Irrigation Shortage Reduces 2015/16 Rice Production. 2015. Available online: https://ipad.fas.usda.gov/highlights/2015/10/th/index.htm (accessed on 25 June 2020).
- Hussain, T.; Gollany, H.T.; Hussain, N.; Ahmed, M.; Tahir, M.; Duangpan, S. Synchronizing Nitrogen Fertilization and Planting Date to Improve Resource Use Efficiency, Productivity, and Profitability of Upland Rice. Front. Plant Sci. 2022, 13, 895811. [Google Scholar] [CrossRef] [PubMed]
- FAO. FAOSTAT. 2020. Available online: http://www.fao.org/faostat/en/#home (accessed on 18 August 2021).
- Hussain, T.; Hussain, N.; Ahmed, M.; Nualsri, C.; Duangpan, S. Responses of lowland rice genotypes under terminal water stress and identification of drought tolerance to stabilize rice productivity in southern Thailand. Plants 2021, 10, 2565. [Google Scholar] [CrossRef] [PubMed]
- Ullah, H.; Mohammadi, A.; Datta, A. Growth, yield and water productivity of selected lowland Thai rice varieties under different cultivation methods and alternate wetting and drying irrigation. Ann. Appl. Biol. 2018, 173, 302–312. [Google Scholar] [CrossRef]
- El Sherbiny, H.A.; El-Hashash, E.F.; Abou El-Enin, M.M.; Nofal, R.S.; Abd El-Mageed, T.A.; Bleih, E.M.; El-Saadony, M.T.; El-Tarabily, K.A.; Shaaban, A. Exogenously Applied Salicylic Acid Boosts Morpho-Physiological Traits, Yield, and Water Productivity of Lowland Rice under Normal and Deficit Irrigation. Agronomy 2022, 12, 1860. [Google Scholar] [CrossRef]
- Hussain, T.; Anothai, J.; Nualsri, C.; Soonsuwon, W. Evaluating performance of sixteen upland rice genotypes under field conditions for further breeding process. J. Agric. Sci. 2018, 10, 144. [Google Scholar] [CrossRef][Green Version]
- Ray, D.K.; West, P.C.; Clark, M.; Gerber, J.S.; Prishchepov, A.V.; Chatterjee, S. Climate change has likely already affected global food production. PLoS ONE 2019, 14, e0217148. [Google Scholar] [CrossRef] [PubMed]
- Ullah, H.; Santiago-Arenas, R.; Ferdous, Z.; Attia, A.; Datta, A. Improving Water Use Efficiency, Nitrogen Use Efficiency, and Radiation Use Efficiency in Field Crops under Drought Stress: A Review, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2019; Volume 156, ISBN 9780128175989. [Google Scholar]
- Watcharin, S.; Kanrob, P.; Charassri, N.; Nattapon, J. Improving photoperiod insensitivity of a Thai upland rice variety by marker-assisted foreground selection. Int. J. Agric. Technol. 2020, 16, 199–206. [Google Scholar]
- Ahmadikhah, A.; Marufinia, A. Effect of reduced plant height on drought tolerance in rice. 3 Biotech 2016, 6, 221. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wang, X.; Jiao, Y.; Qin, Y.; Liu, X.; Zhang, Q.; Fan, L.; Wang, X. Global genome expression analysis of rice in response to drought and high-salinity stresses in shoot, flag leaf, and panicle. Plant Mol. Biol. 2007, 63, 591–608. [Google Scholar] [CrossRef] [PubMed]
- Lafitte, H.R.; Yongsheng, G.; Yan, S.; Li, Z.K. Whole plant responses, key processes, and adaptation to drought stress: The case of rice. J. Exp. Bot. 2007, 58, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, P.; Parida, S.K.; Raghuvanshi, S.; Kapoor, S.; Khurana, P.; Khurana, J.P.; Tyagi, A.K. Rice Improvement Through Genome-Based Functional Analysis and Molecular Breeding in India. Rice 2016, 9, 1. [Google Scholar] [CrossRef] [PubMed]
- Pantuwan, G.; Fukai, S.; Cooper, M.; Rajatasereekul, S.; O’Toole, J.C. Yield response of rice (Oryza sativa L.) genotypes to drought under rainfed lowland 3. Plant factors contributing to drought resistance. Field Crops Res. 2002, 73, 181–200. [Google Scholar] [CrossRef]
- Ovenden, B.; Milgate, A.; Lisle, C.; Wade, L.J.; Rebetzke, G.J.; Holland, J.B. Selection for water-soluble carbohydrate accumulation and investigation of genetic × environment interactions in an elite wheat breeding population. Theor. Appl. Genet. 2017, 130, 2445–2461. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Kobayashi, N.; Ito, O.; Wahid, A.; Serraj, R. Broader leaves result in better performance of indica rice under drought stress. J. Plant Physiol. 2010, 167, 1066–1075. [Google Scholar] [CrossRef] [PubMed]
- Chutia, J.; Borah, S.P. Water Stress Effects on Leaf Growth and Chlorophyll Content but Not the Grain Yield in Traditional Rice (Oryza sativa Linn.) Genotypes of Assam, India II. Protein and Proline Status in Seedlings under PEG Induced Water Stress. Am. J. Plant Sci. 2012, 3, 971–980. [Google Scholar] [CrossRef]
- Kandel, B.P.; Joshi, L.P.; Sharma, S.; Adhikari, P.; Koirala, B.; Shrestha, K. Drought tolerance screening of rice genotypes in mid-hills of Nepal using various drought indices. Acta Agric. Scand. Sect. B Soil Plant Sci. 2022, 72, 744–750. [Google Scholar] [CrossRef]
- Blum, A. Drought adaptation in cereal crops. In Drought Adaptation in Cereals; Ribaut, J.-M., Ed.; Haworth Food Products Press: New York, NY, USA, 2006; pp. 301–333. [Google Scholar]
- Haefele, S.M.; Kato, Y.; Singh, S. Climate ready rice: Augmenting drought tolerance with best management practices. Field Crops Res. 2016, 190, 60–69. [Google Scholar] [CrossRef]
- Lal, R.; Delgado, J.A.; Gulliford, J.; Nielsen, D.; Rice, C.W.; Van Pelt, R.S. Adapting agriculture to drought and extreme events. J. Soil Water Conserv. 2012, 67, 162–166. [Google Scholar] [CrossRef]
- Hu, Y.C.; Shao, H.B.; Chu, L.Y.; Gang, W. Relationship between water use efficiency (WUE) and production of different wheat genotypes at soil water deficit. Colloids Surf. B Biointerfaces 2006, 53, 271–277. [Google Scholar] [CrossRef]
- Blum, A. Drought resistance, water-use efficiency, and yield potential—Are they compatible, dissonant, or mutually exclusive? Aust. J. Agric. Res. 2005, 56, 1159–1168. [Google Scholar] [CrossRef]
- Monclus, R.; Dreyer, E.; Villar, M.; Delmotte, F.M.; Delay, D.; Petit, J.M.; Barbaroux, C.; Le Thiec, D.; Bréchet, C.; Brignolas, F. Impact of drought on productivity and water use efficiency in 29 genotypes of Populus deltoides x Populus nigra. New Phytol. 2006, 169, 765–777. [Google Scholar] [CrossRef]
- Chuchert, S.; Nualsri, C.; Junsawang, N.; Soonsuwon, W. Genetic diversity, genetic variability, and path analysis for yield and its components in indigenous upland rice (Oryza sativa L. var. glutinosa). Songklanakarin J. Sci. Technol. 2018, 40, 609–616. [Google Scholar] [CrossRef]
- Khomphet, T.; Intana, W.; Promwee, A.; Islam, S.S. Genetic Variability, Correlation, and Path Analysis of Thai Commercial Melon Varieties. Int. J. Agron. 2022, 2022, 7877239. [Google Scholar] [CrossRef]
- Ichsan, C.N.; Basyah, B.; Zakaria, S.; Efendi, E. Differences of water status and relationship with roots growth and yield of rice under water stress. Syst. Rev. Pharm. 2020, 11, 611–618. [Google Scholar]
- Schober, P.; Schwarte, L.A. Correlation coefficients: Appropriate use and interpretation. Anesth. Analg. 2018, 126, 1763–1768. [Google Scholar] [CrossRef] [PubMed]
- Lleras, C. Path Analysis. In Encyclopedia of Social Measurement; Kempf-Leonard, K., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2005; pp. 25–30. [Google Scholar]
- Sari, W.K.; Nualsri, C.; Junsawang, N.; Soonsuwon, W. Path analysis for yield and its components in F1 upland rice hybrids and their parental lines. Songklanakarin J. Sci. Technol. 2019, 41, 1419–1425. [Google Scholar]
- Islam, S.; Anothai, J.; Nualsri, C.; Soonsuwon, W. Correlation and path analysis of phenological traits of Thai upland rice genotypes. Songklanakarin J. Plant Sci. 2020, 7, 133–142. [Google Scholar]
- Raina, A.; Laskar, R.A.; Wani, M.R.; Jan, B.L.; Ali, S.; Khan, S. Gamma Rays and Sodium Azide Induced Genetic Variability in High-Yielding and Biofortified Mutant Lines in Cowpea [Vigna unguiculata (L.) Walp.]. Front. Plant Sci. 2022, 13, 911049. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.; Yousaf, Z.; Din, A.; Munawar, M.; Aftab, A.; Riaz, N.; Younas, A.; Alaraidh, I.A.; Okla, M.K.; AbdElgawad, H. Assessment of Mineral Nutrient Efficiency in Genetically Diverse Spinach Accessions by Biochemical and Functional Marker Strategies. Front. Plant Sci. 2022, 13, 889604. [Google Scholar] [CrossRef] [PubMed]
- Shamuyarira, K.W.; Shimelis, H.A.; Mathew, I.; Tsilo, T.J. Correlation and path coefficient analyses of yield and yield components in drought-tolerant bread wheat populations. South Afr. J. Plant Soil 2019, 36, 367–374. [Google Scholar] [CrossRef]
- Arpali, D.; Yagmur, M. The Determination of Selection Criteria Using Path Analysis in Two Rowed Barley (Hordeum vulgare L. Conv. Distichon). Turkish J. Agrci. Nat. Res. 2015, 2, 248–255. [Google Scholar]
- Duangpan, S.; Buapet, P.; Sujitto, S.; Eksomtramage, T. Early assessment of drought tolerance in oil palm D × P progenies using growth and physiological characters in seedling stage. Plant Genet. Resour. 2018, 16, 544–554. [Google Scholar] [CrossRef]
- Zhou, Q.; Ju, C.X.; Wang, Z.Q.; Zhang, H.; Liu, L.J.; Yang, J.C.; Zhang, J.H. Grain yield and water use efficiency of super rice under soil water deficit and alternate wetting and drying irrigation. J. Integr. Agric. 2017, 16, 1028–1043. [Google Scholar] [CrossRef]
- Duangpan, S.; Tongchu, Y.; Hussain, T.; Eksomtramage, T.; Onthong, J. Beneficial Effects of Silicon Fertilizer on Growth and Physiological Responses in Oil Palm. Agronomy 2022, 12, 413. [Google Scholar] [CrossRef]
- Al-Tabbal, J.A.; Al-Fraihat, A.H. Genetic Variation, Heritability, Phenotypic and Genotypic Correlation Studies for Yield and Yield Components in Promising Barley Genotypes. J. Agric. Sci. 2011, 4, 193–210. [Google Scholar] [CrossRef]
- Covarrubias-Pazaran, G.E. Heritability: Meaning and Computation. Optimizing Breeding Schemes Manual. 22 June 2021. [Google Scholar]
- Teklu, D.H.; Alamerew, K.S.; Gebremichael, D.E. Assessment of genetic variability, genetic advance, correlation and path analysis for morphological traits in sesame genotypes. Asian J. Agric. Res. 2014, 8, 181–194. [Google Scholar] [CrossRef]
- Oladosu, Y.; Rafii, M.Y.; Magaji, U.; Abdullah, N.; Miah, G.; Chukwu, S.C.; Hussin, G.; Ramli, A.; Kareem, I. Genotypic and phenotypic relationship among yield components in rice under tropical conditions. BioMed Res. Int. 2018, 2018, 8936767. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Krualee, S.; Sdoodee, S.; Eksomtramage, T.; Sereeprasert, V. Correlation and path analysis of palm oil yield components in oil palm (Elaeis guineensis Jacq.). Kasetsart J. 2013, 47, 528–533. [Google Scholar]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using principal component analysis and heatmap. Nucleic Acid Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef] [PubMed]
- Campozano, L.; Ballari, D.; Montenegro, M.; Avilés, A. Future Meteorological Droughts in Ecuador: Decreasing Trends and Associated Spatio-Temporal Features Derived from CMIP5 Models. Front. Earth Sci. 2020, 8, 17. [Google Scholar] [CrossRef]
- Spinoni, J.; Vogt, J.V.; Naumann, G.; Barbosa, P.; Dosio, A. Will drought events become more frequent and severe in Europe? Int. J. Climatol. 2018, 1736, 1718–1736. [Google Scholar] [CrossRef]
- Chowdhury, M.K.; Hasan, M.A.; Bahadur, M.M.; Islam, M.R.; Hakim, M.A.; Iqbal, M.A.; Javed, T.; Raza, A.; Shabbir, R.; Sorour, S.; et al. Evaluation of drought tolerance of some wheat (Triticum aestivum L.) genotypes through phenology, growth, and physiological indices. Agron. 2021, 11, 1792. [Google Scholar] [CrossRef]
- Ahmad, A.; Aslam, Z.; Naz, M.; Hussain, S.; Javed, T.; Aslam, S.; Raza, A.; Ali, H.M.; Siddiqui, M.H.; Salem, M.Z.; et al. Exogenous salicylic acid-induced drought stress tolerance in wheat (Triticum aestivum L.) grown under hydroponic culture. PloS ONE 2021, 16, e0260556. [Google Scholar] [CrossRef]
- Islam, M.R.; Sarker, B.C.; Alam, M.A.; Javed, T.; Alam, M.J.; Zaman, M.S.U.; Azam, M.G.; Shabbir, R.; Raza, A.; Habib-ur-Rahman, M.; et al. Yield stability and genotype environment interaction of water deficit stress tolerant Mung Bean (Vigna radiata L. Wilczak) genotypes of Bangladesh. Agron. 2021, 11, 2136. [Google Scholar] [CrossRef]
- Shabbir, R.; Javed, T.; Hussain, S.; Ahmar, S.; Naz, M.; Zafar, H.; Pandey, S.; Chauhan, J.; Siddiqui, M.H.; Pinghua, C. Calcium homeostasis and potential roles to combat environmental stresses in plants. South Afr. J. Bot. 2022, 148, 683–693. [Google Scholar] [CrossRef]
- Singhal, R.K.; Fahad, S.; Kumar, P.; Choyal, P.; Javed, T.; Jinger, D.; Singh, P.; Saha, D.; Bose, B.; Akash, H.; et al. Beneficial elements: New Players in improving nutrient use efficiency and abiotic stress tolerance. Plant Growth Regul. 2022, 1–29. [Google Scholar] [CrossRef]
- Bukhari, M.A.; Yousaf, M.; Ahmad, Z.; Rafay, M.; Shah, A.N.; Abbas, A.; Shah, A.A.; Javed, T.; Afzal, M.; Ali, S.; et al. Enhancing drought stress tolerance in Camelina (Camelina sativa L.) through exogenous application of potassium. Physiol. Plant. 2022, 10, e13779. [Google Scholar] [CrossRef]
- Balbaa, M.G.; Osman, H.T.; Kandil, E.E.; Javed, T.; Lamlom, S.F.; Ali, H.M.; Kalaji, H.M.; Wróbel, J.; Telesiñski, A.; Brysiewicz, A.; et al. Determination of morpho-physiological and yield traits of maize inbred lines (Zea mays L.) under optimal and drought stress conditions. Front. Plant Sci. 2022, 13, 959203. [Google Scholar] [CrossRef]
- Ahmad, A.; Aslam, Z.; Hussain, D.; Bellitürk, K.; Javed, T.; Hussain, S.; Bashir, S.; Raza, A.; Alotaibi, S.; Kalaji, H.M.; et al. Rice straw vermicompost enriched with cellulolytic microbes ameliorate the negative effect of drought in wheat through modulating the morpho-physiological attributes. Front. Environ. Sci. 2022, 10, 902999. [Google Scholar] [CrossRef]
- Ali, A.; Altaf, M.T.; Nadeem, M.A.; Shah, A.N.; Azeem, H.; Baloch, F.S.; Karaköy, T.; Hussain, T.; Duangpan, S.; Aasim, M. Recent advancement in OMICS approaches to enhance abiotic stress tolerance in Legumes. Front. Plant Sci. 2022, 13, 952759. [Google Scholar] [CrossRef]
- Singh, B.; Reddy, K.R.; Redoña, E.D.; Walker, T. Screening of Rice Cultivars for Morpho-Physiological Responses to Early-Season Soil Moisture Stress. Rice Sci. 2017, 24, 322–335. [Google Scholar] [CrossRef]
- Haldhar, S.M.; Kumar, R.; Corrado, G.; Berwal, M.K.; Gora, J.S.; Thaochan, N.; Samadia, D.K.; Hussain, T.; Rouphael, Y.; Kumar, P.; et al. A field screening of a pomegranate (Punica granatum) ex-situ germplasm collection for resistance against the false spider mite (Tenuipalpus punicae). Agriculture 2022, 12, 1686. [Google Scholar] [CrossRef]
- Ali, M.F.; Ali, U.; Bilal, S.; Zulfiqar, U.; Sohail, S.; Hussain, T. Response of sorghum and millet to poultry and farmyard 782 manure–based biochar treatments. Arabian J. Geosci. 2022.15, 1–12. [CrossRef]
- Wu, B.; Lin, X.; Ali, M.F.; Wang, D. Development of an irrigation regime for winter wheat to save water resources by avoiding irrigation at anthesis stage. J. Agron. Crop Sci. 2022, 00, 1–16. [Google Scholar] [CrossRef]
- Aslam, M.A.; Ahmed, M.; Hassan, F.-U.; Afzal, O.; Mehmood, M.Z.; Qadir, G.; Asif, M.; Komal, S.; Hussain, T. Impact of temperature fluctuations on plant morphological and physiological traits. In Building Climate Resilience in Agriculture: Theory, Practice and Future Perspective; Jatoi, W.N., Mubeen, M., Ahmad, A., Cheema, M.A., Lin, Z., Hashmi, M.Z., Eds.; Springer International Publishing: Cham, Switzerland, 2022; pp. 25–52. [Google Scholar]
- Hussain, T.; Anothai, J.; Nualsri, C.; Soonsuwon, W. Application of CSM-CERES-Rice in scheduling irrigation and simulating effect of drought stress on upland rice yield. Indian J. Agric. Res. 2018, 52, 140–145. [Google Scholar] [CrossRef]
- Hussain, N.; Ahmed, M.; Duangpan, S.; Hussain, T.; Taweekun, J. Potential impacts of water stress on rice biomass composition and feedstock availability for bioenergy production. Sustainability 2021, 13, 10449. [Google Scholar] [CrossRef]
- Davatgar, N.; Neishabouri, M.R.; Sepaskhah, A.R.; Soltani, A. Physiological and morphological responses of rice (Oryza sativa L.) to varying water stress management strategies. Int. J. Plant Prod. 2009, 3, 19–32. [Google Scholar] [CrossRef]
- Saikumar, S.; Varma, C.M.K.; Saiharini, A.; Kalmeshwer, G.P.; Nagendra, K.; Lavanya, K.; Ayyappa, D. Grain yield responses to varied level of moisture stress at reproductive stage in an interspecific population derived from Swarna/O. glaberrima introgression line. NJAS Wagening. J. Life Sci. 2016, 78, 111–122. [Google Scholar] [CrossRef]
- Verulkar, S.B.; Mandal, N.P.; Dwivedi, J.L.; Singh, B.N.; Sinha, P.K.; Mahato, R.N.; Dongre, P.; Singh, O.N.; Bose, L.K.; Swain, P.; et al. Breeding resilient and productive genotypes adapted to drought-prone rainfed ecosystem of India. Field Crops Res. 2010, 117, 197–208. [Google Scholar] [CrossRef]
- Pantuwan, G.; Fukai, S.; Cooper, M.; Rajatasereekul, S. Yield response of rice (Oryza sativa L.) genotypes to drought under rainfed lowlands 4. Vegetative stage screening in the dry season. Field Crops Res. 2004, 89, 281–297. [Google Scholar] [CrossRef]
- Torres, R.O.; McNally, K.L.; Cruz, C.V.; Serraj, R.; Henry, A. Screening of rice Genebank germplasm for yield and selection of new drought tolerance donors. Field Crops Res. 2013, 147, 12–22. [Google Scholar] [CrossRef]
- Kumar, A.; Verulkar, S.; Dixit, S.; Chauhan, B.; Bernier, J.; Venuprasad, R.; Zhao, D.; Shrivastava, M.N. Yield and yield-attributing traits of rice (Oryza sativa L.) under lowland drought and suitability of early vigor as a selection criterion. Field Crops Res. 2009, 114, 99–107. [Google Scholar] [CrossRef]
- Poehlman, J. Breeding Field Crops; The Avi Publishing Company, Inc.: Westport, CT, USA, 1979. [Google Scholar]
- Sari, W.K.; Nualsri, C.; Junsawang, N.; Soonsuwon, W. Combining ability and heritability for yield and its related traits in Thai rice. Agric. Nat. Resour. 2020, 54, 229–236. [Google Scholar]
- Sumanth, V.; Suresh, B.G.; Ram, B.J.; Srujana, G. Estimation of genetic variability, heritability and genetic advance for grain yield components in rice (Oryza sativa L.). J. Pharmacogn. Phytochem. 2017, 6, 1437–1439. [Google Scholar]
- Vange, T. Biometrical studies on genetic diversity of some upland rice (Oryza Sativa L.) accessions. Nat. Sci. 2009, 7, 21–27. [Google Scholar]
- Santiago-Arenas, R.; Dhakal, S.; Ullah, H.; Agarwal, A.; Datta, A. Seeding, nitrogen and irrigation management optimize rice water and nitrogen use efficiency. Nutr. Cycl. Agroecosystems 2021, 120, 325–341. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Water Consumption | Days to Maturity | Grain Productivity | Water Use Efficiency | |
|---|---|---|---|---|
| Cultivar (C) | 49929.70 *** | 63879.70 *** | 1522.47 *** | 0.59 *** |
| Treatment (T) | 162.56 *** | 564.10 *** | 1901.32 *** | 0.44 *** |
| Season (S) | 297.56 *** | 217.60 ** | 1490.02 *** | 0.11 *** |
| C × T | 702.19 *** | 942.20 *** | 124.46 ns | 0.07 ** |
| C × S | 1089.19 *** | 1530.40 *** | 927.80 *** | 0.19 *** |
| T × S | 33.06 *** | 52.60 ns | 27.78 ns | 0.00 ns |
| C × T × S | 375.69 *** | 651.00 ** | 129.17 ns | 0.06 ns |
| First Season (A) | Second Season (B) | |||
|---|---|---|---|---|
| Cultivars | Control | Drought Stress | Control | Drought Stress |
| Hom Pathum | 128 ± 3 EF | 132 ± 2 D | 122 ± 0 G | 126 ± 0 F |
| Chor Lung | 170 ± 2 A | 151 ± 1 C | 171 ± 1 A | 175 ± 1 A |
| Dum Ja | 168 ± 3 AB | 173 ± 7 A | 165 ± 0 C | 173 ± 2 AB |
| Sang Yod | 147 ± 1 D | 161 ± 1 BC | 147 ± 1 F | 155 ± 1 E |
| Hom Nang Kaew | 158 ± 5 C | 165 ± 6 AB | 163 ± 0 C | 167 ± 0 CD |
| Hom Chan | 144 ± 2 D | 154 ± 4 C | 150 ± 1 E | 156 ± 0 E |
| Pathum Thani−1 | 131 ± 3 E | 139 ± 7 D | 122 ± 0 G | 126 ± 1 F |
| RD−15 | 124 ± 1 EF | 113 ± 4 E | 108 ± 1 I | 111 ± 0 H |
| Lep Nok | 168 ± 4 AB | 172 ± 2 AB | 167 ± 2 B | 170 ± 3 BC |
| Look Pla | 161 ± 1 BC | 166 ± 3 AB | 159 ± 1 D | 166 ± 0 D |
| Khao Dawk Mali−105 | 124 ± 3 EF | 119 ± 1 E | 109 ± 0 I | 113 ± 1 H |
| Tia Malay Dang | 120 ± 2 F | 131 ± 2 D | 116 ± 1 H | 122 ± 0 G |
| Grain Productivity | Days to Maturity | Water Consumption | Water Use Efficiency | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Control Treatment | GCV | 13.72 | M | 15.18 | M | 25.68 | H | 23.48 | H |
| PCV | 17.17 | M | 15.29 | M | 25.68 | H | 25.34 | H | |
| H2 | 63.93 | H | 98.51 | H | 100.00 | H | 85.88 | H | |
| GA | 5.62 | L | 44.50 | H | 40.31 | H | 0.15 | L | |
| GAM | 22.60 | H | 31.05 | H | 52.89 | H | 44.83 | H | |
| Drought Stress Treatment | GCV | 17.09 | M | 15.19 | M | 25.35 | H | 21.65 | H |
| PCV | 22.14 | H | 15.42 | M | 25.35 | H | 25.62 | H | |
| H2 | 59.60 | M | 97.13 | H | 100.00 | H | 71.40 | H | |
| GA | 4.79 | L | 45.45 | H | 40.80 | H | 0.09 | L | |
| GAM | 27.17 | H | 30.84 | H | 52.23 | H | 37.68 | H | |
| Control | Drought Stress Treatment | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Traits | DM | WC | WUE | CC.GP | Traits | DM | WC | WUE | CC.GP |
| DM | −0.24 | 1.75 | −1.14 | 0.37 | DM | 0.34 | 0.78 | −0.67 | 0.45 |
| WC | −0.23 | 1.76 | −1.14 | 0.39 | WC | 0.33 | 0.79 | −0.66 | 0.47 |
| WUE | 0.18 | −1.38 | 1.46 | 0.27 | WUE | −0.20 | −0.48 | 1.09 | 0.41 |
| Residual effect = 0.005 | Residual effect = 0.026 | ||||||||
| Control | Drought Stress Treatment | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Traits | DM | WC | GP | CC.WUE | Traits | DM | WC | GP | CC.WUE |
| DM | 0.16 | −1.19 | 0.25 | −0.78 | DM | −0.32 | −0.69 | 0.40 | −0.61 |
| WC | 0.16 | −1.20 | 0.27 | −0.78 | WC | −0.31 | −0.70 | 0.41 | −0.60 |
| GP | 0.06 | −0.47 | 0.68 | 0.27 | GP | −0.14 | −0.33 | 0.88 | 0.41 |
| Residual effect = 0.002 | Residual effect = 0.021 | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussain, T.; Hussain, N.; Tahir, M.; Raina, A.; Ikram, S.; Maqbool, S.; Fraz Ali, M.; Duangpan, S. Impacts of Drought Stress on Water Use Efficiency and Grain Productivity of Rice and Utilization of Genotypic Variability to Combat Climate Change. Agronomy 2022, 12, 2518. https://doi.org/10.3390/agronomy12102518
Hussain T, Hussain N, Tahir M, Raina A, Ikram S, Maqbool S, Fraz Ali M, Duangpan S. Impacts of Drought Stress on Water Use Efficiency and Grain Productivity of Rice and Utilization of Genotypic Variability to Combat Climate Change. Agronomy. 2022; 12(10):2518. https://doi.org/10.3390/agronomy12102518
Chicago/Turabian StyleHussain, Tajamul, Nurda Hussain, Muhammad Tahir, Aamir Raina, Sobia Ikram, Saliha Maqbool, Muhammad Fraz Ali, and Saowapa Duangpan. 2022. "Impacts of Drought Stress on Water Use Efficiency and Grain Productivity of Rice and Utilization of Genotypic Variability to Combat Climate Change" Agronomy 12, no. 10: 2518. https://doi.org/10.3390/agronomy12102518
APA StyleHussain, T., Hussain, N., Tahir, M., Raina, A., Ikram, S., Maqbool, S., Fraz Ali, M., & Duangpan, S. (2022). Impacts of Drought Stress on Water Use Efficiency and Grain Productivity of Rice and Utilization of Genotypic Variability to Combat Climate Change. Agronomy, 12(10), 2518. https://doi.org/10.3390/agronomy12102518