

Identification of SNP Markers Associated with Grain Quality Traits in a Barley Collection (Hordeum vulgare L.) Harvested in Kazakhstan
 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
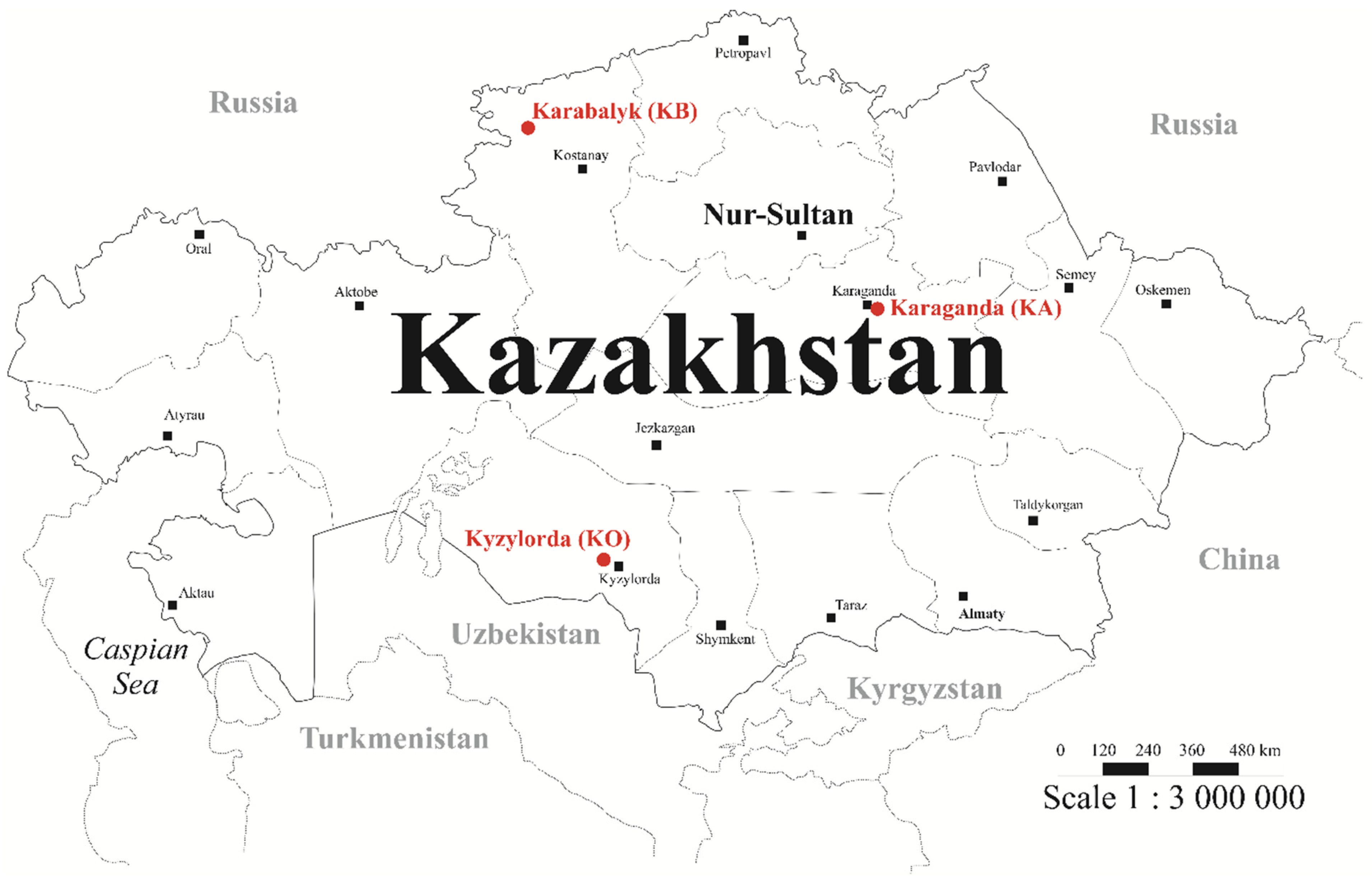
2.1. Plant Material and Grain Quality Traits Assessment
2.2. Genotyping, Population Structure, and GWAS Analysis
2.3. Development of KASP Assays and Their Assessment
3. Results
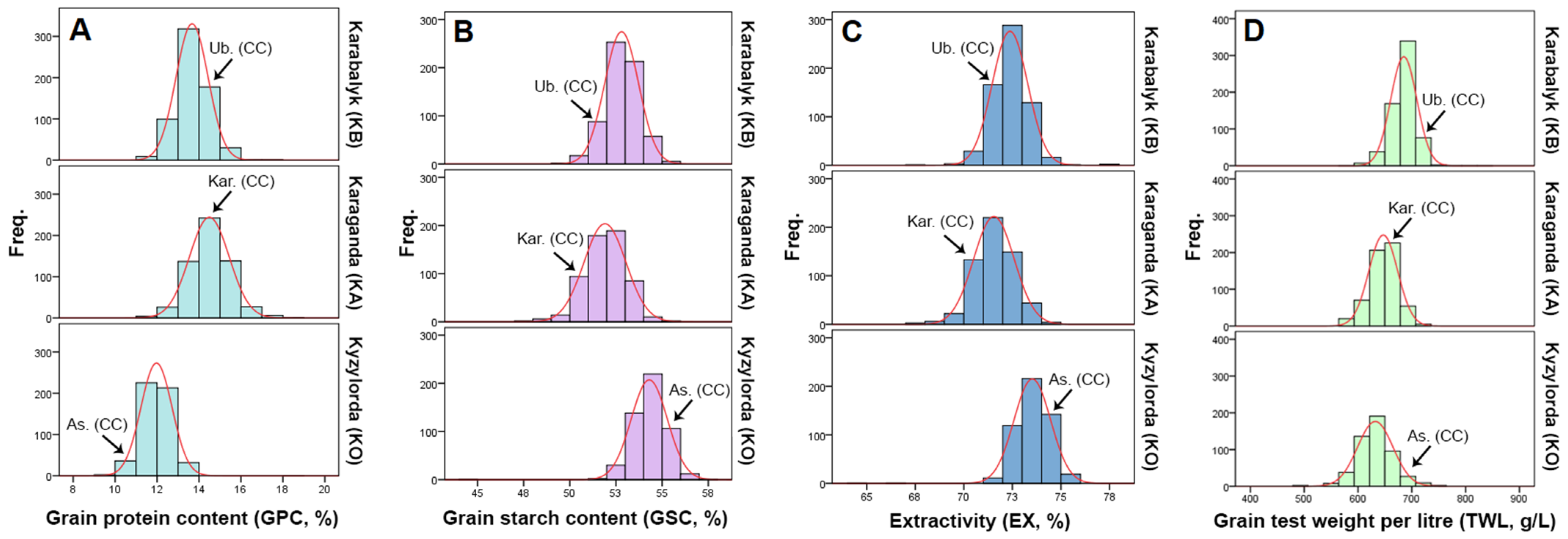
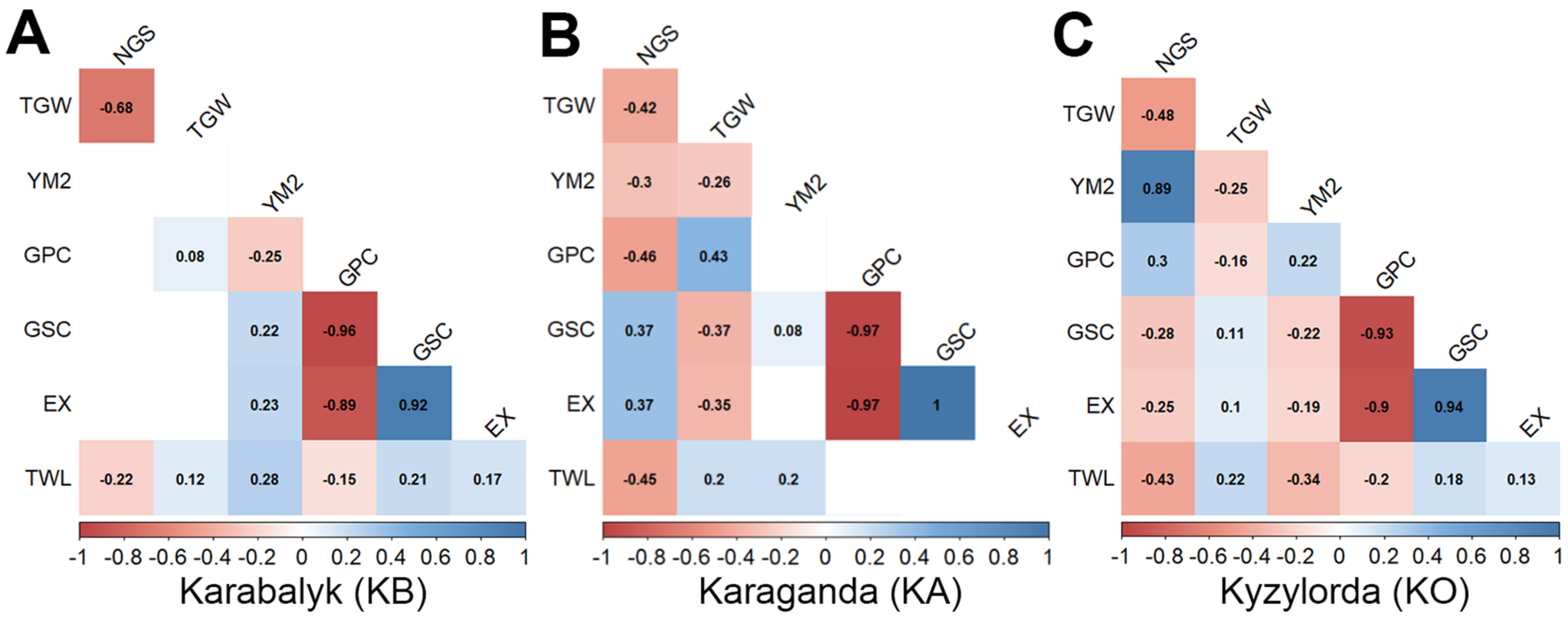
3.1. Variation in Grain Quality Traits in the Collection
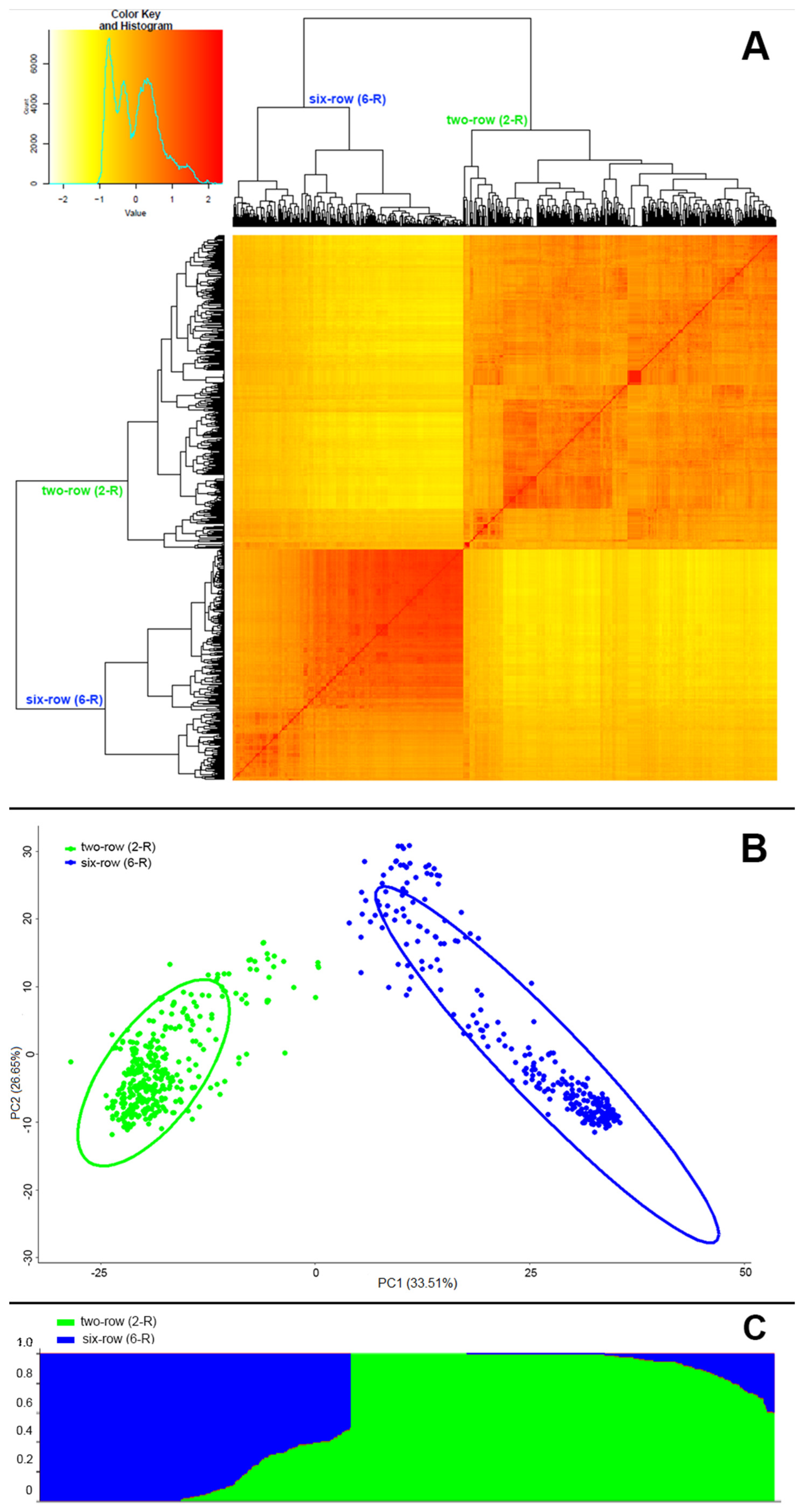
3.2. Population Structure in the Collection of Two- and Six-Rowed Barley Accessions
3.3. Identification of Marker-Trait Associations for Grain Quality Traits Using GWAS
3.4. The significance of KASP Assays for SNP Markers in identified MTAs for Grain Quality Traits
4. Discussion
4.1. The Variability Ranges in the Quality Traits of the Barley Collection Harvested in Three Regions of Kazakhstan
4.2. Identification of QTLs Associated with Quality Traits Based on GWAS
4.3. The Significance of KASP Assays for Evaluation of Grain Quality Traits
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Langridge, P. Economic and Academic Importance of Barley. In The Barley Genome. Compendium of Plant Genomes; Stein, N., Muehlbauer, G.J., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 1–10. [Google Scholar]
- Briggs, D.E. Malts and Malting; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1998. [Google Scholar]
- Emebiri, L.C.; Moody, D.B.; Horsley, R.; Panozzo, J.; Read, B.J. The genetic control of grain protein content variation in a doubled haploid population derived from a cross between Australian and North American two-rowed barley lines. J. Cereal Sci. 2015, 41, 107–114. [Google Scholar] [CrossRef]
- Jamar, C.; Loffet, F.; Frettinger, P.; Ramsay, L.; Fauconnier, M.L.; Du Jardin, P. NAM-1 gene polymorphism and grain protein content in Hordeum. J. Plant Physiol. 2010, 167, 497–501. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.; Yu, G.; Chen, X.; Huang, Y.; Jiang, X.; Zhang, G.; Jin, X. Grain protein content variation and its association analysis in barley. BMC Plant Biol. 2013, 13, 35. [Google Scholar] [CrossRef]
- Hagenblad, J.; Vanhala, T.; Madhavan, S.; Leino, M.W. Protein content and HvNAM alleles in Nordic barley (Hordeum vulgare) during a century of breeding. Hereditas 2022, 159, 12. [Google Scholar] [CrossRef]
- Holtekjølen, A.K.; Uhlen, A.K.; Bråthen, E.; Sahlstrøm, S.; Knutsen, S.H. Contents of starch and non-starch polysaccharides in barley varieties of different origin. Food Chem. 2006, 94, 348–358. [Google Scholar] [CrossRef]
- Langenaeken, N.A.; De Schepper, C.F.; De Schutter, D.P.; Courtin, C.M. Carbohydrate content and structure during malting and brewing: A mass balance study. J. Inst. Brew. 2020, 126, 253–262. [Google Scholar] [CrossRef]
- Collins, H.M.; Betts, N.S.; Dockter, C.; Berkowitz, O.; Braumann, I.; Cuesta-Seijo, J.A.; Skadhauge, B.; Whelan, J.; Bulone, V.; Fincher, G.B. Genes that mediate starch metabolism in developing and germinated barley grain. Front. Plant Sci. 2021, 12, 208. [Google Scholar] [CrossRef]
- Li, M.; Du, J.; Zhang, K. Profiling of carbohydrates in commercial beers and their influence on beer quality. J. Sci. Food Agric. 2020, 100, 3062–3070. [Google Scholar] [CrossRef] [PubMed]
- Titova, Y.M.; Vnukova, M.A. Productivity and quality of malting barley varieties. Bull. Oryol State Agrar. Univ. 2008, 3, 5–8. (In Russian) [Google Scholar]
- Hunt, C.W. Factors affecting the feeding quality of barley for ruminants. Anim. Feed Sci. Technol. 1996, 62, 37–48. [Google Scholar] [CrossRef]
- Campbell, L.D.; Boila, R.J.; Stothers, S.C. Variation in the chemical composition and test weight of barley and wheat grain grown at selected locations throughout Manitoba. Can. J. Anim. Sci. 1995, 75, 239–246. [Google Scholar] [CrossRef]
- Romagosa, I.; Fox, P.N. Genotype×environment interaction and adaptation. In Plant Breeding: Principles and Prospects; Hayward, M.D., Bosemark, N.O., Romagosa, I., Eds.; Chapman & Hall: London, UK, 1993; pp. 373–390. [Google Scholar]
- Molina-Cano, J.L.; Francesch, M.; Perez-Vendrell, A.M.; Ramo, T.; Voltas, J.; Brufau, J. Genetic and environmental variation in malting and feed quality of barley. J. Cereal Sci. 1997, 25, 37–47. [Google Scholar] [CrossRef]
- Kaczmarek, Z.; Adamski, T.; Surma, M.; Jezowsk, S.; Leśniewska-Frątczak, M. Genotype-environment interaction of barley doubled haploids with regard to malting quality. Plant Breed. 1999, 118, 243–247. [Google Scholar] [CrossRef]
- Emebiri, L.C.; Moody, D.B.; Panozzo, J.F.; Chalmers, K.J.; Kretschmer, J.M.; Ablett, G.A. Identification of QTLs associated with variations in grain protein concentration in two-row barley. Aust. J. Agric. Res. 2003, 54, 1211–1221. [Google Scholar] [CrossRef]
- Fan, C.; Zhai, H.; Wang, H.; Yue, Y.; Zhang, M.; Li, J.; Wen, S.; Guo, G.; Zeng, Y.; Ni, Z.; et al. Identification of QTLs controlling grain protein concentration using a high-density SNP and SSR linkage map in barley (Hordeum vulgare L.). BMC Plant Biol. 2017, 17, 122. [Google Scholar] [CrossRef]
- Abdel-Haleem, H.; Bowman, J.; Giroux, M.; Kanazin, V.; Talbert, H.; Surber, L.; Blake, T. Quantitative trait loci of acid detergent fiber and grain chemical composition in hulled×hull-less barley population. Euphytica 2010, 172, 405–418. [Google Scholar] [CrossRef]
- Tinker, N.A.; Mather, D.E.; Rossnagel, B.G.; Kasha, K.J.; Kleinhofs, A.; Hayes, P.M.; Falk, D.E.; Ferguson, T.; Shugar, L.P.; Legge, W.G.; et al. Regions of the genome that affect agronomic performance in two-row barley. Crop Sci. 1996, 36, 1053–1062. [Google Scholar] [CrossRef]
- Marquez-Cedillo, L.A.; Hayes, P.M.; Kleinhofs, A.; Legge, W.G.; Rossnagel, B.G.; Sato, K.; Ullrich, S.E.; Wesenberg, D.M. QTL analysis of agronomic traits in barley based on the doubled haploid progeny of two elite North American varieties representing different germplasm groups. Theor. Appl. Genet. 2001, 103, 625–637. [Google Scholar] [CrossRef]
- Szucs, P.; Blake, V.C.; Bhat, P.R.; Chao, S.; Close, T.J.; Cuesta-Marcos, A.; Muehlbauer, G.J.; Ramsay, L.; Waugh, R.; Hayes, P.M. An integrated resource for barley linkage map and malting quality QTL alignment. Plant Genome 2009, 2, 134–140. [Google Scholar] [CrossRef]
- Peñalba, J.V.; Wolf, J.B. From molecules to populations: Appreciating and estimating recombination rate variation. Nat. Rev. Genet. 2020, 21, 476–492. [Google Scholar] [CrossRef]
- Stumpf, M.P.; McVean, G.A. Estimating recombination rates from population-genetic data. Nat. Rev. Genet. 2003, 4, 959–968. [Google Scholar] [CrossRef] [PubMed]
- Close, T.J.; Bhat, P.R.; Lonardi, S.; Wu, Y.H.; Rostoks, N.; Ramsay, L.; Druka, A.; Stein, N.; Svensson, J.T.; Wanamaker, S.; et al. Development and implementation of high-throughput SNP genotyping in barley. BMC Genom. 2009, 10, 582. [Google Scholar] [CrossRef] [PubMed]
- Close, T.J.; Wanamaker, S.I.; Caldo, R.A.; Turner, S.M.; Ashlock, D.A.; Dickerson, J.A.; Wing, R.A.; Muehlbauer, G.J.; Kleinhofs, A.; Wise, R.P. A new resource for cereal genomics: 22K barley genechip comes of age. Plant Physiol. 2004, 134, 960–968. [Google Scholar] [CrossRef]
- Korte, A.; Farlow, A. The advantages and limitations of trait analysis with GWAS: A review. Plant Methods 2013, 9, 29. [Google Scholar] [CrossRef] [PubMed]
- Pauli, D.; Muehlbauer, G.J.; Smith, K.P.; Cooper, B.; Hole, D.; Obert, D.E.; Ullrich, S.E.; Blake, T.K. Association mapping of agronomic QTLs in US spring barley breeding germplasm. Plant Genome 2014, 7, 11. [Google Scholar] [CrossRef]
- Tsai, H.Y.; Janss, L.L.; Andersen, J.R.; Orabi, J.; Jensen, J.D.; Jahoor, A.; Jensen, J. Genomic prediction and GWAS of yield, quality and disease-related traits in spring barley and winter wheat. Sci. Rep. 2020, 10, 3347. [Google Scholar] [CrossRef]
- Pasam, R.K.; Sharma, R.; Malosetti, M.; van Eeuwijk, F.A.; Haseneyer, G.; Kilian, B.; Graner, A. Genome-wide association studies for agronomical traits in a world wide spring barley collection. BMC Plant Biol. 2012, 12, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Geng, L.; Xie, S.; Wu, D.; Ye, L.; Zhang, G. Genome-wide association study on total starch, amylose and amylopectin in barley grain reveals novel putative alleles. Int. J. Mol. Sci. 2021, 22, 553. [Google Scholar] [CrossRef]
- Zhang, Y.; Luan, H.; Zang, H.; Yang, H.; Xu, X.; Chen, H.; Qiao, H.; Shen, H. Identification of molecular markers for starch content in barley (Hordeum vulgare L.) by genome-wide association studies based on bulked samples. Plant Genet. Resour. 2020, 18, 111–119. [Google Scholar] [CrossRef]
- Houston, K.; Russell, J.; Schreiber, M.; Halpin, C.; Oakey, H.; Washington, J.M.; Booth, A.; Shirley, N.; Burton, R.A.; Fincher, G.B. A genome wide association scan for (1, 3; 1, 4)-β-glucan content in the grain of contemporary 2-row Spring and Winter barleys. BMC Genom. 2014, 15, 907. [Google Scholar] [CrossRef] [PubMed]
- Berger, G.L.; Liu, S.; Hall, M.D.; Brooks, W.S.; Chao, S.; Muehlbauer, G.J.; Baik, B.K.; Steffenson, B.; Griffey, C.A. Marker-trait associations in Virginia Tech winter barley identified using genome-wide mapping. Theor. Appl. Genet. 2013, 126, 693–710. [Google Scholar] [CrossRef]
- Alqudah, A.M.; Sharma, R.; Pasam, R.K.; Graner, A.; Kilian, B.; Schnurbusch, T. Genetic dissection of photoperiod response based on GWAS of pre-anthesis phase duration in spring barley. PLoS ONE 2014, 9, e113120. [Google Scholar] [CrossRef]
- Thabet, S.G.; Moursi, Y.S.; Sallam, A.; Karam, M.A.; Alqudah, A.M. Genetic associations uncover candidate SNP markers and genes associated with salt tolerance during seedling developmental phase in barley. Environ. Exp. Bot. 2021, 188, 104499. [Google Scholar] [CrossRef]
- Al-Sayaydeh, R.; Al-Bawalize, A.; Al-Ajlouni, Z.; Akash, M.W.; Abu-Elenein, J.; Al-Abdallat, A.M. Agronomic evaluation and yield performance of selected barley (Hordeum vulgare L.) landraces from Jordan. Int. J. Agron. 2019, 2019, 9575081. [Google Scholar] [CrossRef]
- Gyawali, S.; Mamidi, S.; Chao, S.; Bhardwaj, S.C.; Shekhawat, P.S.; Selvakumar, R.; Gangwar, O.P.; Singh Verma, R.P. Genome-wide association studies revealed novel stripe rust resistance QTL in barley at seedling and adult-plant stages. Euphytica 2021, 217, 3. [Google Scholar] [CrossRef]
- Mitterbauer, E.; Enders, M.; Bender, J.; Erbs, M.; Habekuß, A.; Kilian, B.; Ordon, F.; Weigel, H.-J. Growth response of 98 barley (Hordeum vulgare L.) genotypes to elevated CO 2 and identification of related quantitative trait loci using genome-wide association studies. Plant Breed. 2017, 136, 483–497. [Google Scholar] [CrossRef]
- Comadran, J.; Kilian, B.; Russell, J.; Ramsay, L.; Stein, N.; Ganal, M.; Shaw, P.; Bayer, M.; Thomas, W.; Marshall, D. Natural variation in a homolog of Antirrhinum CENTRORADIALIS contributed to spring growth habit and environmental adaptation in cultivated barley. Nat. Genet. 2012, 44, 1388–1392. [Google Scholar] [CrossRef]
- Bayer, M.M.; Rapazote-Flores, P.; Ganal, M.; Hedley, P.E.; Macaulay, M.; Plieske, J.; Ramsay, L.; Russell, J.; Shaw, P.D.; Thomas, W.; et al. Development and Evaluation of a Barley 50k iSelect SNP Array. Front. Plant Sci. 2017, 8, 1792. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.E.; Chao, S.; Singh, R.P.; Sorrells, M.E. Validation of single nucleotide polymorphic markers linked to Ug99 stem rust resistance in spring wheat. PLoS ONE 2017, 12, e0171963. [Google Scholar] [CrossRef]
- Tan, C.T.; Assanga, S.; Zhang, G.; Rudd, J.C.; Haley, S.D.; Xue, Q.; Ibrahim, A.; Bai, G.; Zhang, X.; Byrne, P.; et al. Development and validation of KASP markers for wheat streak mosaic virus resistance gene Wsm2. Crop Sci. 2017, 57, 340–349. [Google Scholar] [CrossRef]
- Khalid, M.; Afzal, F.; Gul, A.; Amir, R.; Subhani, A.; Ahmed, Z.; Mahmood, Z.; Xia, X.; Rasheed, A.; He, Z. Molecular characterization of 87 functional genes in wheat diversity panel and their association with phenotypes under well-watered and water-limited conditions. Front. Plant Sci. 2019, 10, 717. [Google Scholar] [CrossRef]
- Comastri, A.; Janni, M.; Simmonds, J.; Uauy, C.; Pignone, D.; Nguyen, H.T. Marmiroli, N. Heat in wheat: Exploit reverse genetic techniques to discover new alleles within the Triticum durum sHsp26 family. Front. Plant Sci. 2018, 9, 1337. [Google Scholar] [CrossRef]
- Rasheed, A.; Wen, W.; Gao, F.; Zhai, S.; Jin, H.; Liu, J.; Guo, Q.; Zhang, Y.; Dreisigacker, S.; Xia, X.; et al. Development and validation of KASP assays for genes underpinning key economic traits in bread wheat. Theor. Appl. Genet. 2016, 129, 1843–1860. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Zhang, H.; Li, S.; Zou, Y.; Li, T.; Liu, J.; Ding, P.; Mu, Y.; Tang, H.; Deng, M.; et al. Identification of quantitative trait loci for kernel traits in a wheat cultivar Chuannong16. BMC Genet. 2019, 20, 77. [Google Scholar] [CrossRef] [PubMed]
- Jost, M.; Singh, D.; Lagudah, E.; Park, R.F.; Dracatos, P. Fine mapping of leaf rust resistance gene Rph13 from wild barley. Theor. Appl. Genet. 2020, 133, 1887–1895. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Mornhinweg, D.; Bernardo, A.; Li, G.; Bian, R.; Bai, G. Characterization of Rsg2.a3: A new greenbug resistance allele at the Rsg2 locus from wild barley (Hordeum vulgare ssp. spontaneum). Crop J. 2022, in press. [Google Scholar] [CrossRef]
- Wang, Y.; Habekuß, A.; Snowdon, R.J.; Ordon, F.; Perovic, D. Delineating the elusive BaMMV resistance gene rym15 in barley by medium-resolution mapping. Mol. Breed. 2021, 41, 76. [Google Scholar] [CrossRef]
- Yan, L.; Jiang, C.; Cai, Y.; Pan, Y.; Xu, R.; Luan, H.; Shen, H.; Ahmar, S.; Zhang, J.; Yang, P.; et al. Evaluating the genetic effects of seed dormancy regulatory genes Qsd1 and Qsd2 in a global collection of cultivated barley (Hordeum vulgare ssp. vulgare) with functional kompetitive allele-specific PCR markers. Plant Breed. 2021, 140, 827–834. [Google Scholar]
- Looseley, M.E.; Bayer, M.; Bul, H.; Ramsay, L.; Thomas, W.; Booth, A.; De La Fuente Canto, C.; Morris, J.; Hedley, P.E.; Russell, J. Association mapping of diastatic power in UK winter and spring barley by exome sequencing of phenotypically contrasting variety sets. Front. Plant Sci. 2017, 8, 1566. [Google Scholar] [CrossRef]
- Cu, S.T.; March, T.J.; Degner, S.; Eglinton, J.K. Identification of novel alleles from wild barley for the improvement of alpha-amylase and related malt quality traits. Plant Breed. 2016, 135, 663–670. [Google Scholar] [CrossRef]
- Genievskaya, Y.; Almerekova, S.; Sariev, B.; Chudinov, V.; Tokhetova, L.; Sereda, G.; Ortaev, A.; Tsygankov, V.; Blake, T.; Chao, S.; et al. Marker–trait associations in two–rowed spring barley accessions from Kazakhstan and the USA. PLoS ONE 2018, 3, e0205421. [Google Scholar] [CrossRef]
- Almerekova, S.; Sariev, B.; Abugalieva, A.; Chudinov, V.; Sereda, G.; Tokhetova, L.; Ortaev, A.; Tsygankov, V.; Blake, T.; Chao, S.; et al. Association mapping for agronomic traits in six–rowed spring barley from the USA harvested in Kazakhstan. PLoS ONE 2019, 14, e0221064. [Google Scholar] [CrossRef]
- Turuspekov, Y.; Ormanbekova, D.; Rsaliev, A.; Abugalieva, S. Genome-wide association study on stem rust resistance in Kazakh spring barley lines. BMC Plant Biol. 2016, 16, 13–21. [Google Scholar] [CrossRef]
- Turuspekov, Y.; Sariev, B.; Chudinov, V.; Sereda, G.; Tokhetova, L.; Doszhanov, M.; Volis, S.; Abugalieva, S. Genotype × environment interaction patterns for grain yield of spring barley in different regions of Kazakhstan. Russ. J. Genet. 2013, 49, 196–205. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Yu, J.; Pressoir, G.; Briggs, W.H.; Vroh Bi, I.; Yamasaki, M.; Doebley, J.F.; McMullen, M.D.; Gaut, B.S.; Nielsen, D.M.; Holland, J.B.; et al. A unified mixed-model method for association mapping that accounts for multiple levels of relatedness. Nat. Genet. 2006, 38, 203–208. [Google Scholar] [CrossRef]
- Falush, D.; Wirth, T.; Linz, B.; Pritchard, J.K.; Stephens, M.; Kidd, M.; Blaser, M.J.; Graham, D.Y.; Vacher, S.; Perez-Perez, G.I.; et al. Traces of human migrations in Helicobacter pylori populations. Science 2003, 299, 1582–1585. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Earl, D.A.; von Holdt, B.M. Structure Harvester: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Bosnes, M.; Weideman, F.; Olsen, O.-A. Endosperm differentiation in barley wild-type and sex mutants. Plant J. 1992, 2, 661–674. [Google Scholar] [CrossRef]
- Maertens, K.; Reyns, P.; De Baerdemaeker, J. On-line measurement of grain quality with NIR technology. Trans. ASABE 2004, 47, 1135. [Google Scholar] [CrossRef]
- Kahriman, F.; Egesel, C.Ö. Development of a calibration model to estimate quality traits in wheat flour using NIR (Near Infrared Reflectance) spectroscopy. Res. J. Agric. Sci. 2011, 43, 392–400. [Google Scholar]
- Wang, H.; Smith, K.P.; Combs, E.; Blake, T.; Horsley, R.D.; Muehlbauer, G.J. Effect of population size and unbalanced data sets on QTL detection using genome-wide association mapping in barley breeding germplasm. Theor. Appl. Genet. 2012, 124, 111–124. [Google Scholar] [CrossRef]
- Liu, Y.J.; Papasian, C.J.; Liu, J.F.; Hamilton, J.; Deng, H.W. Is replication the gold standard for validating genome-wide association findings? PLoS ONE 2008, 3, e4037. [Google Scholar] [CrossRef]
- Stanley, D.; Rejzek, M.; Naested, H.; Smedley, M.; Otero, S.; Fahy, B.; Thorpe, F.; Nash, R.J.; Harwood, W.; Svensson, B.; et al. The role of α-glucosidase in germinating barley grains. Plant Physiol. 2011, 155, 932–943. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhao, H.; Tang, J.; Li, Z.; Li, Z.; Chen, D.; Lin, W. A proteomic study on molecular mechanism of poor grain-filling of rice (Oryza sativa L.) inferior spikelets. PLoS ONE 2014, 9, e89140. [Google Scholar] [CrossRef]
- Barrero-Sicilia, C.; Hernando-Amado, S.; González-Melendi, P.; Carbonero, P. Structure, expression profile and subcellular localisation of four different sucrose synthase genes from barley. Planta 2011, 234, 391–403. [Google Scholar] [CrossRef]
- See, D.; Kanazin, V.; Kephart, K.; Blake, T. Mapping genes controlling variation in barley grain protein concentration. Crop Sci. 2002, 42, 680–685. [Google Scholar] [CrossRef]
- Riaz, A.; Kanwal, F.; Börner, A.; Pillen, K.; Dai, F.; Alqudah, A.M. Advances in genomics-based breeding of barley: Molecular tools and genomic databases. Agronomy 2021, 11, 894. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GPC | |||||
| df | SS | MS | Var. | % of Total Var. | |
| Geno | 657 | 1336 | 2 | 0.3723 | 15.48 |
| Env | 5 | 4781 | 956.2 | 1.3325 | 55.44 |
| Geno x Env | 2913 | 2508 | 0.9 | 0.6990 | 29.08 |
| Total Var. | 2.4038 | 100.00 | |||
| GSC | |||||
| df | SS | MS | Var. | % of total Var. | |
| Geno | 657 | 1915 | 2.9 | 0.5337 | 18.02 |
| Env | 5 | 4333 | 866.5 | 1.2076 | 40.78 |
| Geno x Env | 2913 | 4378 | 1.5 | 1.2202 | 41.20 |
| Total Var. | 2.9615 | 100.00 | |||
| EX | |||||
| df | SS | MS | Var. | % of total Var. | |
| Geno | 657 | 1707 | 2.6 | 0.4758 | 19.09 |
| Env | 5 | 3115 | 623 | 0.8682 | 34.83 |
| Geno x Env | 2913 | 4121 | 1.4 | 1.1486 | 46.08 |
| Total Var. | 2.4925 | 100.00 | |||
| TWL | |||||
| df | SS | MS | Var. | % of total Var. | |
| Geno | 657 | 1,666,735 | 2510 | 464.5304 | 20.03 |
| Env | 5 | 4,097,249 | 819450 | 1141.931 | 49.23 |
| Geno x Env | 2913 | 2,558,541 | 877 | 713.0828 | 30.74 |
| Total Var. | 2319.544 | 100.00 | |||
| QTL | Trait | Marker | Chr. | Pos. (bp) | Pos. (cM) | Candidate Malting Quality Genes | GPC Candidate QTL | GSC Candidate QTL | TWL Candidate QTL |
|---|---|---|---|---|---|---|---|---|---|
| QTL_Q1 | GPC/GSC/EX | 12_30918 | 1H | 8,935,905 | 12.78 | ||||
| QTL_Q2 | GPC/GSC/EX/TWL | 11_11336 | 1H | 261,773,377 | 50 | QTl1_CPC (55.49 cM) [30] | QTL2_SC (51.23 cM) [30] | ||
| QTL_Q3 | GPC/GSC/EX | 11_10438 | 1H | 303,519,071 | 50 | QTl1_CPC (55.49 cM) [30] | QTL2_SC (51.23 cM) [30] | ||
| QTL_Q4 | TWL | 12_31381 | 1H | 325,808,056 | 50 | QTl1_CPC (55.49 cM) [30] | QTL2_SC (51.23 cM) [30] | ||
| QTL_Q5 | TWL | 12_30478 | 1H | 381,207,730 | 50.99 | QTl1_CPC (55.49 cM) [30] | QTL2_SC (51.23 cM) [30] | ||
| TWL | 12_30499 | 1H | 381,209,230 | 50.99 | |||||
| QTL_Q6 | TWL | 11_10176 | 1H | 420,656,686 | 59.01 | Aglu3 (12_30820, 419012101 bp) α-glucosidase [22] | QTl1_CPC (55.49 cM) [30] | ||
| QTL_Q7 | TWL | 11_20169 | 1H | 516,153,706 | 97.98 | QTL5_SC (126.01 cM) [30]; qTS-1.2 (534442471 bp) [31] | QTw1H.101 (98.56 cM) [28] | ||
| GPC/GSC/TWL | 12_30191 | 1H | 522,448,103 | 107.18 | |||||
| TWL | 11_10338 | 1H | 532,951,913 | 121.6 | |||||
| TWL | 12_31387 | 1H | 542,673,808 | 131.46 | |||||
| TWL | 11_20383 | 1H | 547,250,913 | 136.65 | |||||
| QTL_Q8 | GPC/GSC/EX | 11_10178 | 2H | 48,475,931 | 52.96 | ||||
| QTL_Q9 | GSC | 11_10909 | 2H | 545,242,939 | 69.55 | QTL5_CPC (74.37 cM) [30]; QGpc2H.54 (66.11 cM) [28] | QTL7_SC (64.24 cM) [30]; QTl8_SC (71.12 cM) [30] | ||
| QTL_Q10 | TWL | 12_31293 | 2H | 641,328,117 | 84.69 | QGpc.ZiSc-2H.1 (90.64 cM) [18]; QGpc2H.86 (90.99 cM) [28] | QTL9_SC (90.1 cM) [30] | QTwt-2H.89 (81.26 cM) [28]; QTw2H.86 (90.99 cM) [28] | |
| TWL | 11_10287 | 2H | 651,372,755 | 90.99 | |||||
| TWL | 12_30901 | 2H | 652,031,870 | 90.99 | |||||
| QTL_Q11 | GPC/GSC/EX/TWL | 11_21414 | 2H | 761,624,420 | - | ||||
| QTL_Q12 | GPC/GSC/EX/TWL | 11_21505 | 3H | 580,635,994 | 79.13 | ||||
| QTL_Q13 | TWL | 12_31161 | 3H | 667,790,880 | - | qAP-3.2 (667803604 bp) [31] | |||
| GPC/GSC/EX/TWL | 11_10935 | 3H | 678,512,385 | 149.85 | |||||
| QTL_Q14 | TWL | 11_21303 | 4H | 464,028,169 | 53.87 | DTDP (12_30839, 54.95 cM) d-TDP-glucose dehydratase [22]; PDI (12_30878, 53.87 cM) protein disulfide isomerase [22] | QBgsg.StMo-4H (54.4 cM) [22] | ||
| QTL_Q15 | GPC/GSC/EX | 11_10090 | 4H | 582,935,043 | 69.08 | QTL12_SC (65.05 cM) [30] | |||
| QTL_Q16 | TWL | 12_31139 | 4H | 624,584,147 | 102.38 | QGpc.ZiSc-4H.2 (102.38 cM) [18]; QTL13_CPC (101.62 cM) [30] | |||
| QTL_Q17 | TWL | 12_10077 | 5H | 556,603,185 | 87.71 | QGpc.ZiSc-5H.3 (85.58 cM) [18]; QTL14_CP (85.93 cM) [30]; QGp-5H.96 (87.71 cM) [34] | qAP-5.2 (551372936 bp) [31] | ||
| QTL_Q18 | GPC/GSC/EX | 12_30852 | 5H | 560,732,040 | 87.71 | QGpc.ZiSc-5H.3 (85.58 cM) [18]; QTL14_CP (85.93 cM) [30]; QGp-5H.96 (87.71 cM) [34] | qAP-5.2 (551372936 bp) [31] | ||
| GPC/GSC/EX | 12_30705 | 5H | 561,727,550 | 90.22 | |||||
| QTL_Q19 | TWL | 11_20008 | 5H | 612,229,115 | 134.67 | QGpc5H.137 (127.52 cM) [28] | QTwt-5H.131 (131.64 cM) [28] | ||
| QTL_Q20 | GSC | 11_20232 | 6H | 1,578,951 | 0 | qAP-6.1 (4816646 bp) [31] | |||
| TWL | 11_20493 | 6H | 5,065,147 | 0.5 | |||||
| TWL | 11_20886 | 6H | 5,362,408 | 1.4 | |||||
| QTL_Q21 | GPC | 12_30516 | 6H | 37,274,484 | 51.74 | QGpc6H.45 (54.7 cM) [28] | |||
| QTL_Q22 | GPC/GSC | 12_30658 | 6H | 50,346,904 | 54.14 | QGpc6H.45 (54.7 cM) [28]; Qcp6a (57.91 cM) [19]; | |||
| QTL_Q23 | GPC | 12_31274 | 6H | 64,747,230 | 55.28 | QGpc6H.45 (54.7 cM) [28]; Qcp6a (57.91 cM) [19]; | qAC-6.1 (70242665 bp) [31] | ||
| QTL_Q24 | GPC/GSC/EX/TWL | 12_31509 | 6H | 203,509,034 | 58.91 | QGpc6H.45 (54.7 cM) [28]; Qcp6a (57.91 cM) [19]; | |||
| QTL_Q25 | TWL | 11_20673 | 6H | 502,536,025 | 74.18 | QGpc.ZiSc-6H.1 (73.83 cM) [18]; | QTL18_SC (71.08 cM) [30] | QTw6H.75 (77.7 cM) [28] | |
| QTL_Q26 | TWL | 11_10185 | 6H | 529,879,937 | 81.48 | QTw6H.75 (77.7 cM) [28] | |||
| QTL_Q27 | GSC | 12_30576 | 7H | 66,410,739 | 61.13 | SS1 (12_30879, 67729209 bp) sucrose synthase 1 [22] | QGpc.ZiSc-7H.2 (59.48 cM) [18]; QGpc.ZiSc-7H.3 (63.19 cM) [18]; QTL20_CPC (61.32 cM) [30] | QBgnm.StMo-7H.1 (60.9 cM) [22]; QDp.StMo-2H.3 (60.9 cM) [22] | |
| QTL_Q28 | GPC/GSC | 12_31140 | 7H | 411,695,093 | 78.07 | QTL21_CPC (80.94 cM) [30]; QGpc6H.86 [28] (83.23 cM) | QTL22_SC (78.22 cM) [30] | QTwt-7H.91-94 (84.86 cM) [28] | |
| QTL_Q29 | GPC/GSC/EX/TWL | 11_21103 | 7H | 582,767,743 | - | ||||
| QTL_Q30 | GPC/GSC | 11_10182 | 7H | 628,806,795 | 133.92 | QGpc7H.130 [28] (135.99 cM) |
| # of MTA | QTL | Trait | SNP | Chr. | Pos. (bp) | p-Value | FDR Adjusted p-Value | R2 | Allele | Effect |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | QTL_Q2 | GPC | 11_11336 | 1H | 261,773,377 | 6.37 × 10−6 | 0.0039 | 0.023 | A | 0.495 |
| 2 | GSC | 1.03 × 10−4 | 0.0396 | 0.019 | G | 0.565 | ||||
| 3 | QTL_Q8 | GSC | 11_10178 | 2H | 48,475,931 | 7.64 × 10−5 | 0.0367 | 0.020 | G | 0.455 |
| 4 | QTL_Q12 | GPC | 11_21505 | 3H | 580,635,994 | 2.14 × 10−6 | 0.0021 | 0.026 | A | 0.590 |
| 5 | TWL | 1.15 × 10−7 | 0.0002 | 0.035 | A | 20.275 | ||||
| 6 | GSC | 6.01 × 10−5 | 0.0367 | 0.021 | G | 0.647 | ||||
| 7 | QTL_Q13 | GPC | 11_10935 | 3H | 678,512,385 | 1.25 × 10−4 | 0.0482 | 0.017 | C | 0.391 |
| 8 | TWL | 2.75 × 10−7 | 0.0003 | 0.033 | C | 16.244 | ||||
| 9 | QTL_Q20 | TWL | 11_20886 | 6H | 53,62,408 | 4.96 × 10−5 | 0.0190 | 0.020 | A | 8.744 |
| 10 | QTL_Q24 | EX | 12_31509 | 6H | 203,509,034 | 7.21 × 10−6 | 0.0138 | 0.027 | A | 0.656 |
| 11 | GPC | 9.10 × 10−8 | 0.0002 | 0.033 | G | 0.664 | ||||
| 12 | TWL | 3.53 × 10−6 | 0.0017 | 0.027 | G | 17.627 | ||||
| 13 | GSC | 2.40 × 10−6 | 0.0046 | 0.029 | A | 0.759 | ||||
| 14 | QTL_Q29 | GPC | 11_21103 | 7H | 582,767,743 | 8.17 × 10−6 | 0.0039 | 0.023 | A | 0.494 |
| 15 | TWL | 1.85 × 10−6 | 0.0012 | 0.028 | A | 16.331 | ||||
| 16 | GSC | 5.26 × 10−5 | 0.0367 | 0.021 | G | 0.591 |
| Traits (GWAS) | KASP | Chr. | MAF | 2020 | 2021 | MEAN | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TWL | GPC | EX | GSC | TWL | GPC | EX | GSC | TWL | GPC | EX | GSC | ||||
| GPC/GSC/TWL | ipbb_hv_6 | 3H | 0.18 | 0.684 | 3.47 × 10−4 | 2.88 × 10−4 | 1.23 × 10−5 | 0.283 | 9.16 × 10−5 | 5.89 × 10−5 | 1.14 × 10−5 | 0.703 | 4.69 × 10−6 | 4.85 × 10−5 | 5.38 × 10−8 |
| GPC/GSC/ EX/TWL | ipbb_hv_7 | 6H | 0 (mono) | - | - | - | - | - | - | - | - | - | - | - | - |
| GPC/GSC | ipbb_hv_116 | 1H | 0.12 | 0.444 | 0.495 | 0.875 | 0.525 | 0.152 | 0.768 | 0.105 | 0.994 | 0.308 | 0.906 | 0.531 | 0.718 |
| GPC/TWL | ipbb_hv_119 | 3H | 0 (mono) | - | - | - | - | - | - | - | - | - | - | - | - |
| GPC/GSC/TWL | ipbb_hv_128 | 7H | 0.17 | 0.497 | 0.005 | 0.647 | 0.016 | 0.966 | 0.264 | 0.649 | 0.092 | 0.642 | 0.123 | 0.665 | 0.037 |
| 2020 | |||||
| ipbb_hv_6 (chromosome 3H; 580,635,994 bp) | Genotype | N | Mean | SD | Effect |
| Grain protein content (GPC,%) | A:A | 6 | 11.45 | 0.23 | −1.15% |
| G:G | 28 | 12.60 | 1.44 | +1.15% | |
| Grain starch content (GSC,%) | A:A | 6 | 61.27 | 0.45 | +1.71% |
| G:G | 28 | 59.56 | 1.19 | −1.71% | |
| Extractivity (EX,%) | A:A | 6 | 78.32 | 0.42 | +1.10% |
| G:G | 28 | 77.22 | 0.84 | −1.10% | |
| ipbb_hv_128 (chromosome 7H; 582,767,743 bp) | Genotype | N | Mean | SD | Effect |
| Grain protein content (GPC,%) | T:T | 5 | 11.64 | 0.25 | −0.89% |
| C:C | 29 | 12.53 | 1.45 | +0.89% | |
| Grain starch content (GSC,%) | T:T | 5 | 60.95 | 0.76 | +1.26% |
| C:C | 29 | 59.69 | 1.27 | −1.26% | |
| 2021 | |||||
| ipbb_hv_6 (chromosome 3H; 580,635,994 bp) | Genotype | N | Mean | SD | Effect |
| Grain protein content (GPC,%) | A:A | 6 | 12.60 | 0.83 | −2.60% |
| G:G | 28 | 15.20 | 1.10 | +2.60% | |
| Grain starch content (GSC,%) | A:A | 6 | 62.13 | 0.53 | +2.05% |
| G:G | 28 | 60.08 | 0.78 | −2.05% | |
| Extractivity (EX,%) | A:A | 6 | 78.65 | 0.30 | +0.93% |
| A:A | 28 | 77.72 | 0.69 | −0.93% | |
| MEAN | |||||
| ipbb_hv_6 (chromosome 3H; 580,635,994 bp) | Genotype | N | Mean | SD | Effect |
| Grain protein content (GPC,%) | A:A | 6 | 12.03 | 19.80 | −1.87% |
| G:G | 28 | 13.90 | 31.69 | +1.87% | |
| Grain starch content (GSC,%) | A:A | 6 | 61.70 | 0.36 | +1.87% |
| G:G | 28 | 59.83 | 0.81 | −1.87% | |
| Extractivity (EX,%) | A:A | 6 | 78.48 | 0.33 | +0.99% |
| G:G | 28 | 77.49 | 0.68 | −0.99% | |
| ipbb_hv_128 (chromosome 7H; 582,767,743 bp) | Genotype | N | Mean | SD | Effect |
| Grain starch content (GSC,%) | T:T | 5 | 61.24 | 0.96 | +1.27% |
| C:C | 29 | 59.97 | 0.94 | −1.27% | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Genievskaya, Y.; Almerekova, S.; Abugalieva, S.; Chudinov, V.; Blake, T.; Abugalieva, A.; Turuspekov, Y. Identification of SNP Markers Associated with Grain Quality Traits in a Barley Collection (Hordeum vulgare L.) Harvested in Kazakhstan. Agronomy 2022, 12, 2431. https://doi.org/10.3390/agronomy12102431
Genievskaya Y, Almerekova S, Abugalieva S, Chudinov V, Blake T, Abugalieva A, Turuspekov Y. Identification of SNP Markers Associated with Grain Quality Traits in a Barley Collection (Hordeum vulgare L.) Harvested in Kazakhstan. Agronomy. 2022; 12(10):2431. https://doi.org/10.3390/agronomy12102431
Chicago/Turabian StyleGenievskaya, Yuliya, Shyryn Almerekova, Saule Abugalieva, Vladimir Chudinov, Thomas Blake, Aigul Abugalieva, and Yerlan Turuspekov. 2022. "Identification of SNP Markers Associated with Grain Quality Traits in a Barley Collection (Hordeum vulgare L.) Harvested in Kazakhstan" Agronomy 12, no. 10: 2431. https://doi.org/10.3390/agronomy12102431
APA StyleGenievskaya, Y., Almerekova, S., Abugalieva, S., Chudinov, V., Blake, T., Abugalieva, A., & Turuspekov, Y. (2022). Identification of SNP Markers Associated with Grain Quality Traits in a Barley Collection (Hordeum vulgare L.) Harvested in Kazakhstan. Agronomy, 12(10), 2431. https://doi.org/10.3390/agronomy12102431