
Longevity and Parasitism Capacity of Psyttalia concolor (Hymenoptera: Braconidae) Fed on Sugar Solutions and Insect Honeydew



Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Food Sources
2.3. Longevity and Parasitism Potential of P. concolor Fed on Different “Sugars”
2.3.1. Longevity
2.3.2. Parasitism Capacity
2.4. Longevity of P. concolor Fed on Honeydew
2.5. Data Analysis
2.5.1. Longevity
2.5.2. Parasitism Capacity
3. Results
3.1. Longevity and Parasitism Capacity of Psyttalia concolor Fed on Different Sugars
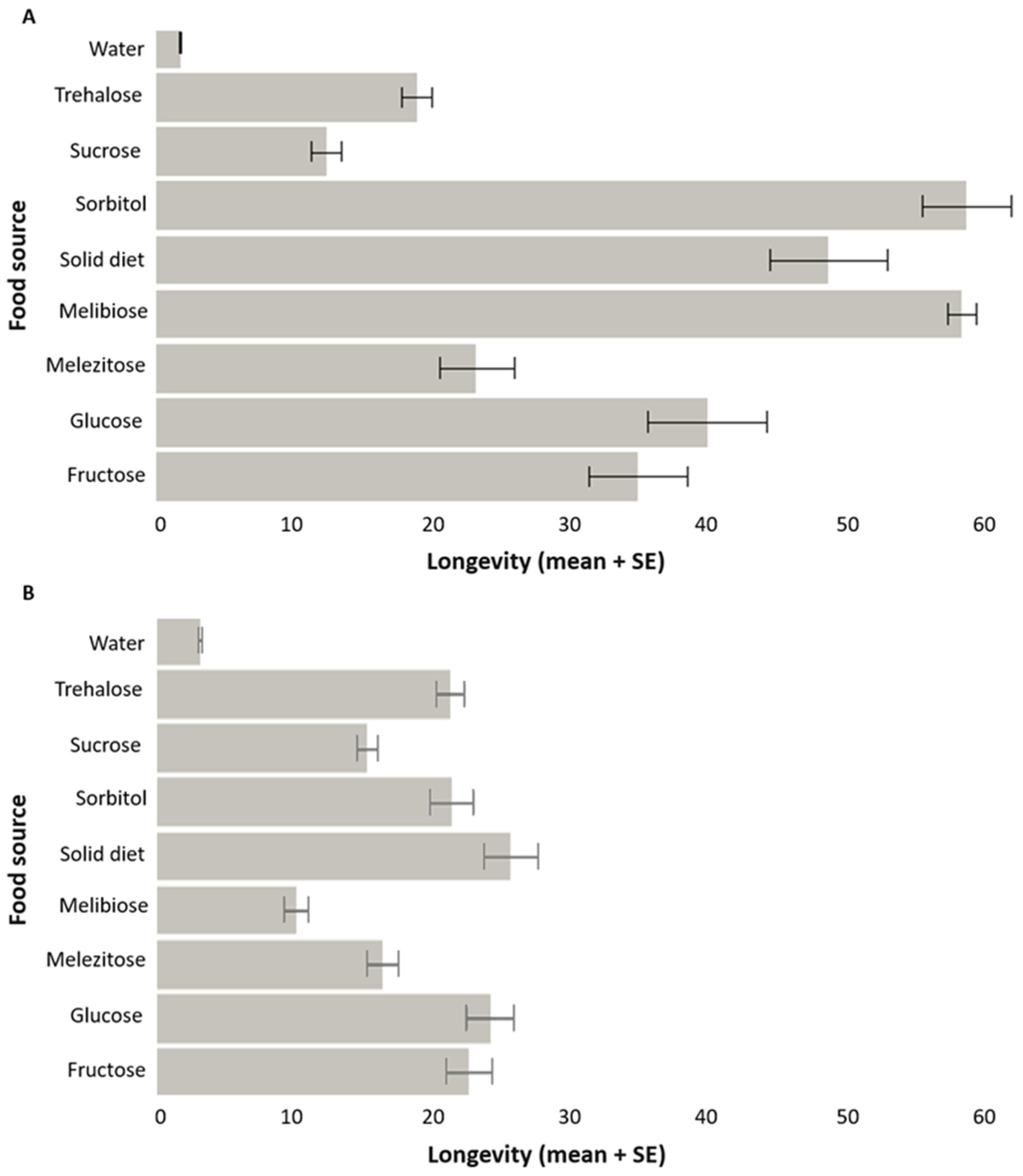
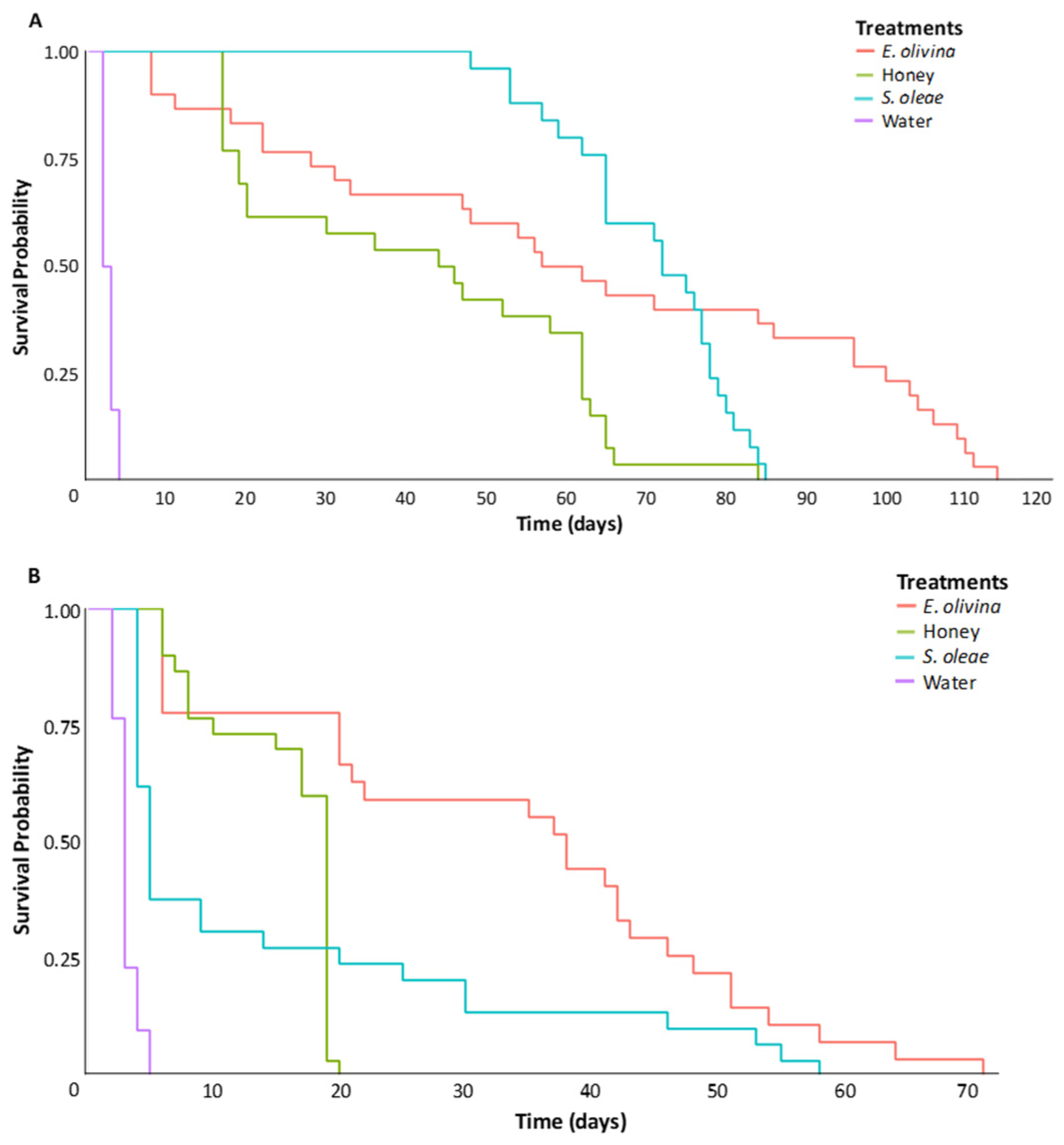
3.1.1. Longevity
3.1.2. Parasitism Capacity
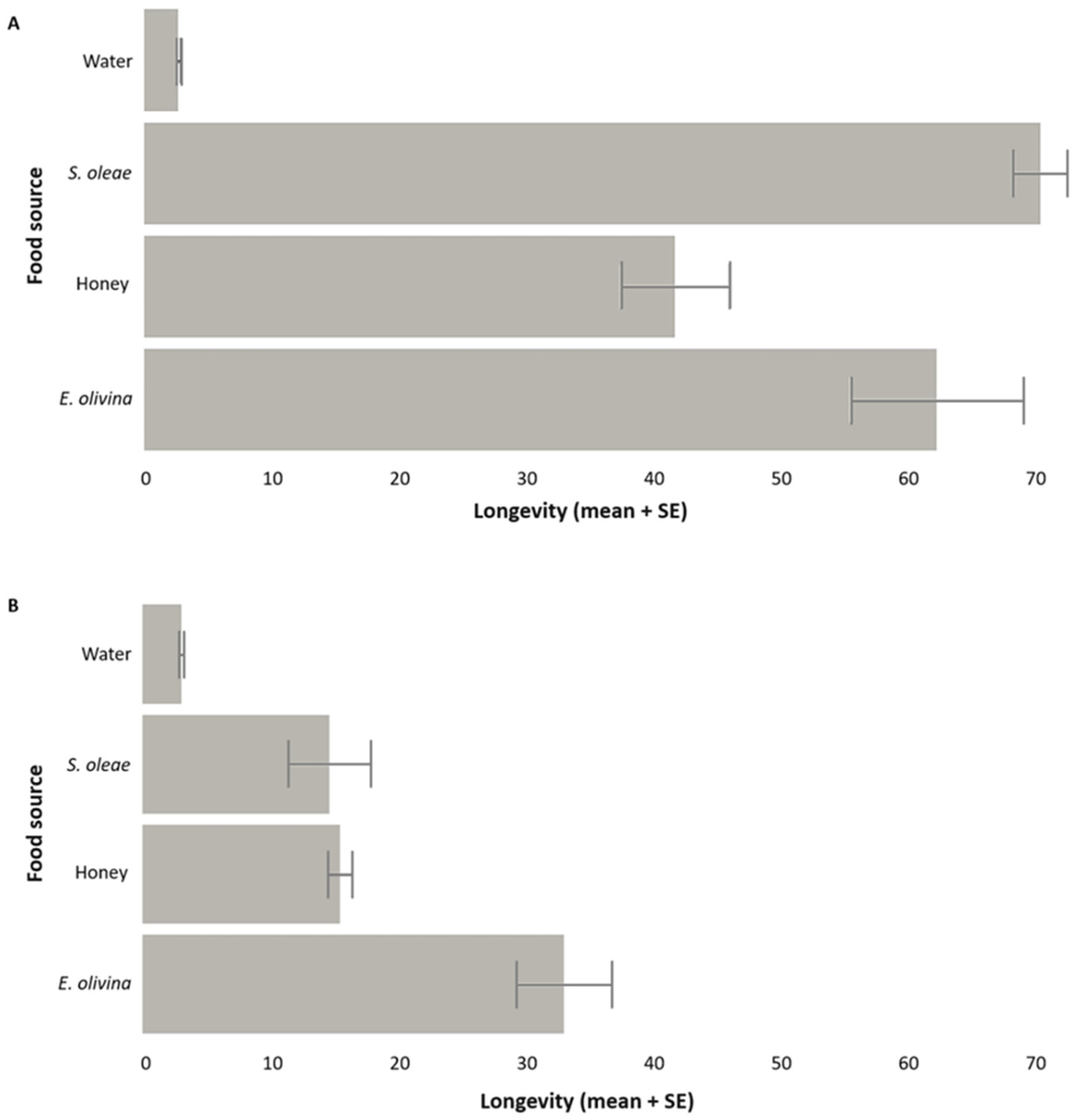
3.2. Longevity of P. concolor Fed on Honeydew
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wäckers, F.L. Suitability of (extra-)floral nectar, pollen, and honeydew as insect food sources. In Plant-Provided Food for Carnivorous Insects, A Protective Mutualism and its Applications; Wäckers, F.L., van Rijn, P.C.J., Bruin, J., Eds.; Cambridge University Press: Cambridge, UK, 2005; pp. 17–74. [Google Scholar]
- Wäckers, F.L.; van Rijn, P.C.J.; Heimpel, G.E. Honeydew as a food source for natural enemies: Making the best of a bad meal? Biol. Control 2008, 45, 176–184. [Google Scholar] [CrossRef]
- Benelli, G.; Giunti, G.; Tena, A.; Desneux, N.; Caselli, A.; Canale, A. The impact of adult diet on parasitoid reproductive performance. J. Pest Sci. 2017, 90, 807–823. [Google Scholar] [CrossRef]
- Jervis, M.A.; Kidd, N.A.C. Host-feeding strategies in hymenopteran parasitoids. Biol. Rev. 1986, 61, 395–434. [Google Scholar] [CrossRef]
- Baker, H.G.; Baker, I. Floral nectar sugar constituents in relation to pollinator type. In Handbook of Experimental Pollination Biology; Jones, C.E., Little, R.J., Eds.; Van Nostrand Reinhold: New York, NY, USA, 1983; pp. 117–141. [Google Scholar]
- Petanidou, T.; Van Laere, A.; Ellis, W.N.; Smets, E. What shapes amino acid and sugar composition in Mediterranean floral nectars? Oikos 2006, 115, 155–169. [Google Scholar] [CrossRef]
- Wäckers, F.L. A comparison of nectar- and honeydew sugars with respect to their utilization by the hymenopteran parasitoid Cotesia glomerata. J. Insect Physiol. 2001, 47, 1077–1084. [Google Scholar] [CrossRef]
- Russell, M. A meta-analysis of physiological and behavioral responses of parasitoid wasps to flowers of individual plant species. Biol. Control 2015, 82, 96–103. [Google Scholar] [CrossRef]
- Nave, A.; Gonçalves, F.; Nunes, F.M.; Campos, M.; Torres, L. Evaluating potential sugar food sources from the olive grove agroecosystems for Prays oleae parasitoid Chelonus elaeaphilus. Biocontrol. Sci. Techn. 2017, 27, 686–695. [Google Scholar] [CrossRef]
- Hogervorst, P.A.M.; Wäckers, F.L.; Romeis, J. Effects of honeydew sugar composition on the longevity of Aphidius ervi. Entomol. Exp. Appl. 2007, 122, 223–232. [Google Scholar] [CrossRef]
- Hogervorst, P.A.M.; Wäckers, F.L.; Romeis, J. Detecting nutritional state and food source use in field-collected insects that synthesize honeydew oligosaccharides. Funct. Ecol. 2007, 21, 936–946. [Google Scholar] [CrossRef]
- Winkler, K.; Wäckers, F.; Bukovinszkine-Kiss, G.; van Lenteren, J. Sugar resources are vital for Diadegma semiclausum fecundity under field conditions. Basic Appl. Ecol. 2006, 7, 133–140. [Google Scholar] [CrossRef]
- Lee, J.C.; Heimpel, G.E. Floral resources impact longevity and oviposition rate of a parasitoid in the field. J. Anim. Ecol. 2008, 77, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Tena, A.; Pekas, A.; Cano, D.; Wäckers, F.L.; Urbaneja, A. Sugar provisioning maximizes the biocontrol service of parasitoids. J. Appl. Ecol. 2015, 52, 795–804. [Google Scholar] [CrossRef]
- Olson, D.M.; Fadamiro, H.; Lundgren, J.G.; Heimpel, G.E. Effects of sugar feeding on carbohydrate and lipid metabolism in a parasitoid wasp. Physiol. Entomol. 2000, 25, 17–26. [Google Scholar] [CrossRef]
- Lee, J.C.; Heimpel, G.E.; Leibee, G.L. Comparing floral nectar and aphid honeydew diets on the longevity and nutrient levels of a parasitoid wasp. Entomol. Exp. Appl. 2004, 111, 189–199. [Google Scholar] [CrossRef]
- Nafziger, T.D.; Fadamiro, H.Y. Suitability of some farmscaping plants as nectar sources for the parasitoid wasp, Microplitis croceipes (Hymenoptera: Braconidae): Effects on longevity and body nutrients. Biol. Control 2011, 56, 225–229. [Google Scholar] [CrossRef]
- Snart, C.J.P.; Kapranas, A.; Williams, H.; Barrett, D.A.; Hardy, I.C.W. Sustenance and Performance: Nutritional Reserves, Longevity, and Contest Outcomes of Fed and Starved Adult Parasitoid Wasps. Front. Ecol. Environ. 2018, 6, 12. [Google Scholar] [CrossRef]
- Lewis, W.J.; Stapel, J.O.; Cortesero, A.M.; Takasu, K. Understanding how parasitoids balance food and host needs: Importance to biological control. Biol. Control 1998, 11, 175–183. [Google Scholar] [CrossRef]
- Lucchetta, P.; Bernstein, C.; Théry, M.; Lazzari, C.; Desouhant, E. Foraging and associative learning of visual signals in a parasitic wasp. Anim. Cogn. 2008, 11, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Delucchi, V. Les parasites de la mouche des olives. Entomophaga 1957, 2, 107–118. [Google Scholar] [CrossRef]
- Raspi, A.; Loni, A. Alcune note sull’allevamento massale di Opius concolor Szépligeti (Hym.: Braconidae) e su recenti tentative d’introduzione della specie in Toscana e Liguria [Some notes on the breeding of massale Opius concolor Szépligeti (Hym.: Braconidae) and recent tentative introduction of the species in Tuscany and Liguria]. Frustula Entomol. 1994, 30, 135–145. [Google Scholar]
- Daane, K.M.; Johnson, M.W. Olive fruit fly: Managing an ancient pest in modern times. Annu. Rev. Entomol. 2010, 55, 151–169. [Google Scholar] [CrossRef] [PubMed]
- Miranda, M.A.; Miquel, M.; Terrassa, J.; Melis, N.; Monerris, M. Parasitism of Bactrocera oleae (Diptera; Tephritidae) by Psyttalia concolor (Hymenoptera; Braconidae) in the Balearic Islands (Spain). J. Appl. Entomol. 2008, 132, 798–805. [Google Scholar] [CrossRef]
- Delrio, G.; Lentini, A.; Satta, A. Biological control of olive fruit fly through inoculative releases of Opius concolor Szépl. IOBC/WPRS Bull. 2005, 28, 53–58. [Google Scholar]
- Furtado, C.; Belo, A.F.; Nunes, F.M.; Ganhão, C.T.; Müller, C.T.; Torres, L.; Rei, F.T. Evaluating potential olive orchard sugar food sources for the olive fly parasitoid Psyttalia concolor. BioControl 2016, 61, 473–483. [Google Scholar] [CrossRef]
- Jacas, J.A.; Viñuela, E. Analysis of a lab method to test the effects of pesticides on adult females of Opius concolor, a parasitoid of the olive fruit fly Bactrocera oleae. Biocontrol Sci. Techn. 1994, 4, 147–154. [Google Scholar] [CrossRef]
- Bengochea, P.; Budia, F.; Viñuela, E.; Medina, P. Are kaolin and copper treatments safe to the olive fruit fly parasitoid Psyttalia concolor? J. Pest Sci. 2014, 87, 351–359. [Google Scholar] [CrossRef]
- Canale, A.; Benelli, G. Impact of mass-rearing on the host seeking behaviour and parasitism by the fruit fly parasitoid Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae). J. Pest Sci. 2012, 85, 65–74. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014; Available online: http://www.R-project.org/ (accessed on 20 April 2022).
- Williams, L., III; Deschodt, P.; Pointurier, O.; Wyckhuys, K.A.G. Sugar concentration and timing of feeding affect feeding characteristics and survival of a parasitic wasp. J. Insect Physiol. 2015, 79, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, N.; Wang, J.; Wan, F. Effect of six carbohydrate sources on the longevity of a whitefly parasitoid Eretmocerus hayati (Hymenoptera: Aphelinidae). J. Asia-Pac. Entomol. 2014, 17, 723–728. [Google Scholar] [CrossRef]
- Petanidou, T. Sugars in Mediterranean floral nectars: An ecological and evolutionary approach. J. Chem. Ecol. 2005, 31, 1065–1088. [Google Scholar] [CrossRef] [PubMed]
- Burke, C.J.; Waddell, S. Remembering nutrient quality of sugar in Drosophila. Curr. Biol. 2011, 21, 746–750. [Google Scholar] [CrossRef] [PubMed]
- Bale, J.S. Insects and low temperatures: From molecular biology to distributions and abundance. Philos. Trans. R. Soc. B 2002, 357, 849–862. [Google Scholar] [CrossRef] [PubMed]
- Wäckers, F.L. Gustatory response by the hymenopteran parasitoids Cotesia glomerata to a range of nectar and honeydew sugars. J. Chem. Ecol. 1999, 25, 2863–2877. [Google Scholar] [CrossRef]
- Wäckers, F.L.; Lee, J.; Heimpel, G.; Winkler, K.; Wagenaar, R. Hymenopteran parasitoids synthesize honeydew-specific’ oligosaccharides. Funct. Ecol. 2006, 20, 790–798. [Google Scholar] [CrossRef]
- Tompkins, J.M.L.; Wratten, S.D.; Wäckers, F.L. Nectar to improve parasitoid fitness in biological control: Does the sucrose: Hexose ratio matter? Basic App. Ecol. 2010, 11, 264–271. [Google Scholar] [CrossRef]
- Benelli, G.; Gennari, G.; Francini, A.; Canale, A. Longevity costs of same-sex interactions: First evidence from a parasitic wasp. Invertebr. Biol. 2013, 132, 156–162. [Google Scholar] [CrossRef]
- Wäckers, F.L. Do oligosaccharides reduce the suitability of honeydew for predators and parasitoids? A further facet to the function of insect-synthesized honeydew sugars. Oikos 2000, 90, 197–201. [Google Scholar] [CrossRef]
- Heimpel, G.E.; Rosenheim, J.A. Egg limitation in parasitoids: A review of the evidence and a case study. Biol. Control 1998, 11, 160–168. [Google Scholar] [CrossRef]
- Hausmann, C.H.; Wäckers, F.L.; Dorn, S. Sugar convertibility in the parasitoid Cotesia glomerata (Hymenoptera: Braconidae). Arch Insect Biochem. Physiol. 2005, 60, 223–229. [Google Scholar] [CrossRef]
- Tena, A.; Senft, M.; Desneux, N.; Dregni, J.; Heimpel, G.E. The influence of aphid-produced honeydew on parasitoid fitness and nutritional state: A comparative study. Basic App. Ecol. 2018, 29, 55–68. [Google Scholar] [CrossRef]
- Faria, C.A.; Wäckers, F.L.; Turlings, T.C.J. The nutritional value of aphid honeydew for non-aphid parasitoids. Basic App. Ecol. 2008, 9, 286–297. [Google Scholar] [CrossRef]
- Villa, M.; Santos, S.A.P.; Mexia, A.; Bento, A.; Pereira, J.A. Wild flower resources and insect honeydew are potential food items for Elasmus flabellatus. Agron. Sustain. Dev. 2017, 37, 15. [Google Scholar] [CrossRef]
- Wang, X.-G.; Johnson, M.W.; Opp, S.B.; Krugner, R.; Daane, K.M. Honeydew and insecticide bait as competing food resources for a fruit fly and common natural enemies in the olive agroecosystem. Entomol. Exp. Appl. 2011, 139, 128–137. [Google Scholar] [CrossRef]
- Wade, M.R.; Hopkinson, J.E.; Zalucki, M.P. Influence of food supplementation on the fitness of two biological control agents: A predatory nabid bug and a bollworm pupal parasitoid. J. Pest Sci. 2008, 81, 99–107. [Google Scholar] [CrossRef]
- Wade, M.R.; Zalucki, M.P.; Wratten, S.D.; Robinson, K.A. Conservation biological control of arthropods using artificial food sprays: Current status and future challenges. Biol. Control 2008, 45, 185–199. [Google Scholar] [CrossRef]
- Walters, A.S. Weed management systems for no-tillage vegetable production. In Herbicides: Theory and Applications; Soloneski, S., Larramendy, M.L., Eds.; IntechOpen: Rijeka, Croatia, 2011; pp. 2–40. [Google Scholar]
- Segoli, M.; Rosenheim, J.A. Spatial and temporal variation in sugar availability for insect parasitoids in agricultural fields and consequences for reproductive success. Biol. Control 2013, 67, 163–169. [Google Scholar] [CrossRef]
- Tena, A.; Wäckers, F.L.; Heimpel, G.E.; Urbaneja, A.; Pekas, A. Parasitoid nutritional ecology in a community context: The importance of honeydew and implications for biological control. Curr. Opin. Insect Sci. 2016, 14, 100–104. [Google Scholar] [CrossRef]
- Kishinevsky, M.; Cohen, N.; Chiel, E.; Wajnberg, E.; Keasar, T. Sugar feeding of parasitoids in an agroecosystem: Effects of community composition, habitat and vegetation. Insect Conserv. Diver. 2017, 11, 50–57. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Food Source | Non-Emerged Hosts (%) * | Parasitism Rate (%) * |
|---|---|---|
| Solid diet | 89.85 ± 2.25 a | 74.45 ± 2.52 a |
| Sugars | ||
| Monosaccharides | ||
| Fructose | 92.05 ± 1.61 a | 63.34 ± 1.51 ab |
| Glucose | 90.60 ± 1.57 a | 62.99 ± 1.51 ab |
| Disaccharides | ||
| Melibiose | 83.30 ± 3.77 a | 52.16 ± 3.35 bc |
| Sucrose | 94.95 ± 1.22 a | 64.60 ± 1.63 ab |
| Trehalose | 85.10 ± 3.44 a | 62.58 ± 2.59 ab |
| Trisaccharides | ||
| Melezitose | 48.90 ± 6.53 b | 48.36 ± 6.51 c |
| Sugar alcohol | ||
| Sorbitol | 85.20 ± 2.10 a | 62.42 ± 1.63 ab |
| Kruskal–Wallis chi-squared | 57.404 | 42.496 |
| Df | 7 | 7 |
| p-value | <0.001 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinheiro, L.A.; Pereira, J.A.; Medina, P.; Santos, S.A.P. Longevity and Parasitism Capacity of Psyttalia concolor (Hymenoptera: Braconidae) Fed on Sugar Solutions and Insect Honeydew. Agronomy 2022, 12, 2401. https://doi.org/10.3390/agronomy12102401
Pinheiro LA, Pereira JA, Medina P, Santos SAP. Longevity and Parasitism Capacity of Psyttalia concolor (Hymenoptera: Braconidae) Fed on Sugar Solutions and Insect Honeydew. Agronomy. 2022; 12(10):2401. https://doi.org/10.3390/agronomy12102401
Chicago/Turabian StylePinheiro, Lara A., José Alberto Pereira, Pilar Medina, and Sónia A. P. Santos. 2022. "Longevity and Parasitism Capacity of Psyttalia concolor (Hymenoptera: Braconidae) Fed on Sugar Solutions and Insect Honeydew" Agronomy 12, no. 10: 2401. https://doi.org/10.3390/agronomy12102401
APA StylePinheiro, L. A., Pereira, J. A., Medina, P., & Santos, S. A. P. (2022). Longevity and Parasitism Capacity of Psyttalia concolor (Hymenoptera: Braconidae) Fed on Sugar Solutions and Insect Honeydew. Agronomy, 12(10), 2401. https://doi.org/10.3390/agronomy12102401