
Pits of Date Palm: Bioactive Composition, Antibacterial Activity and Antimutagenicity Potentials

 ,
, 
 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Date Material and Extraction Method
2.3. Estimation of Total Flavonoids
2.4. Determination of Total Phenolics
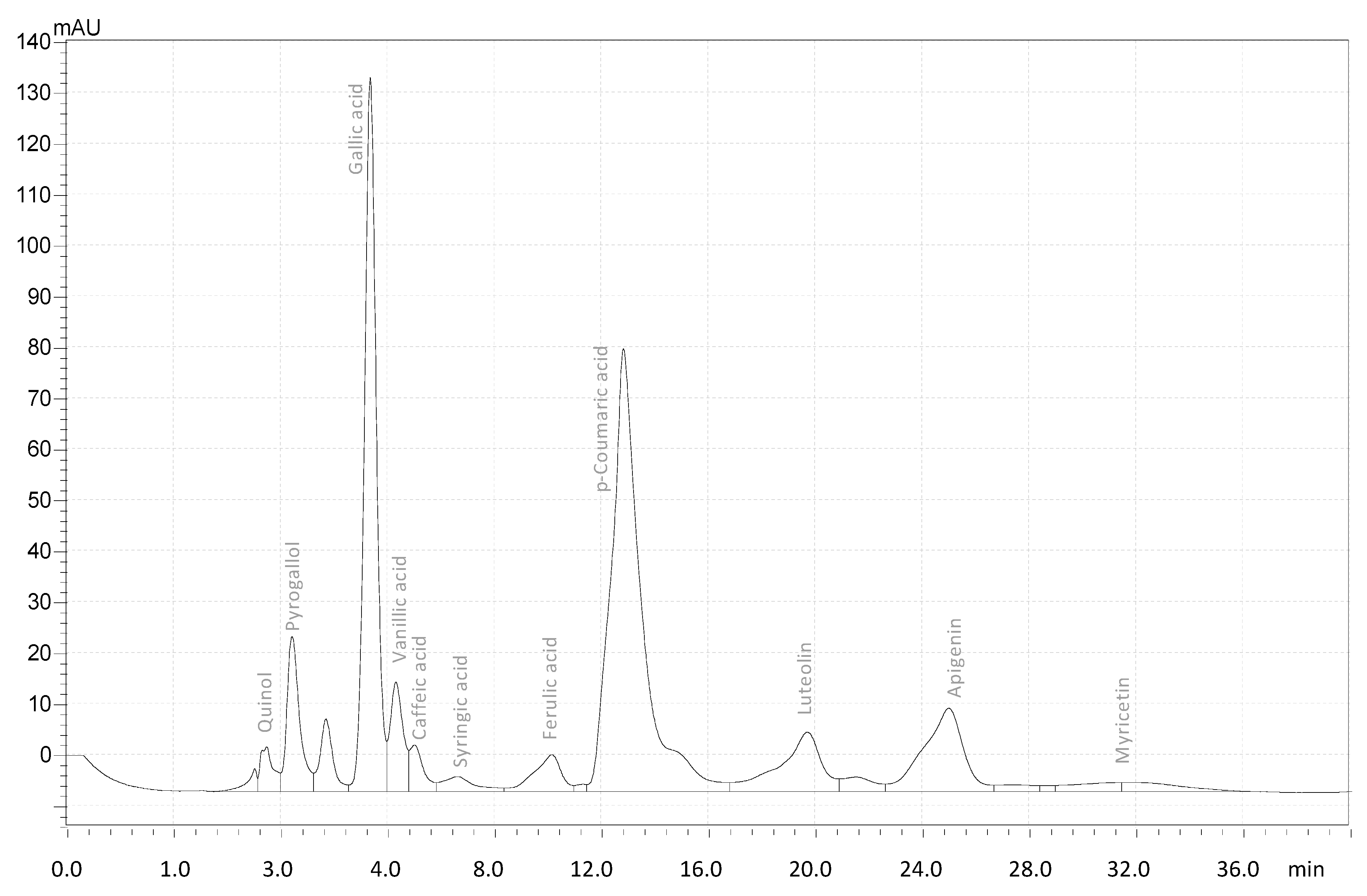
2.5. HPLC
2.6. Extraction, Fractionation, and Isolation of Bioactive Compound
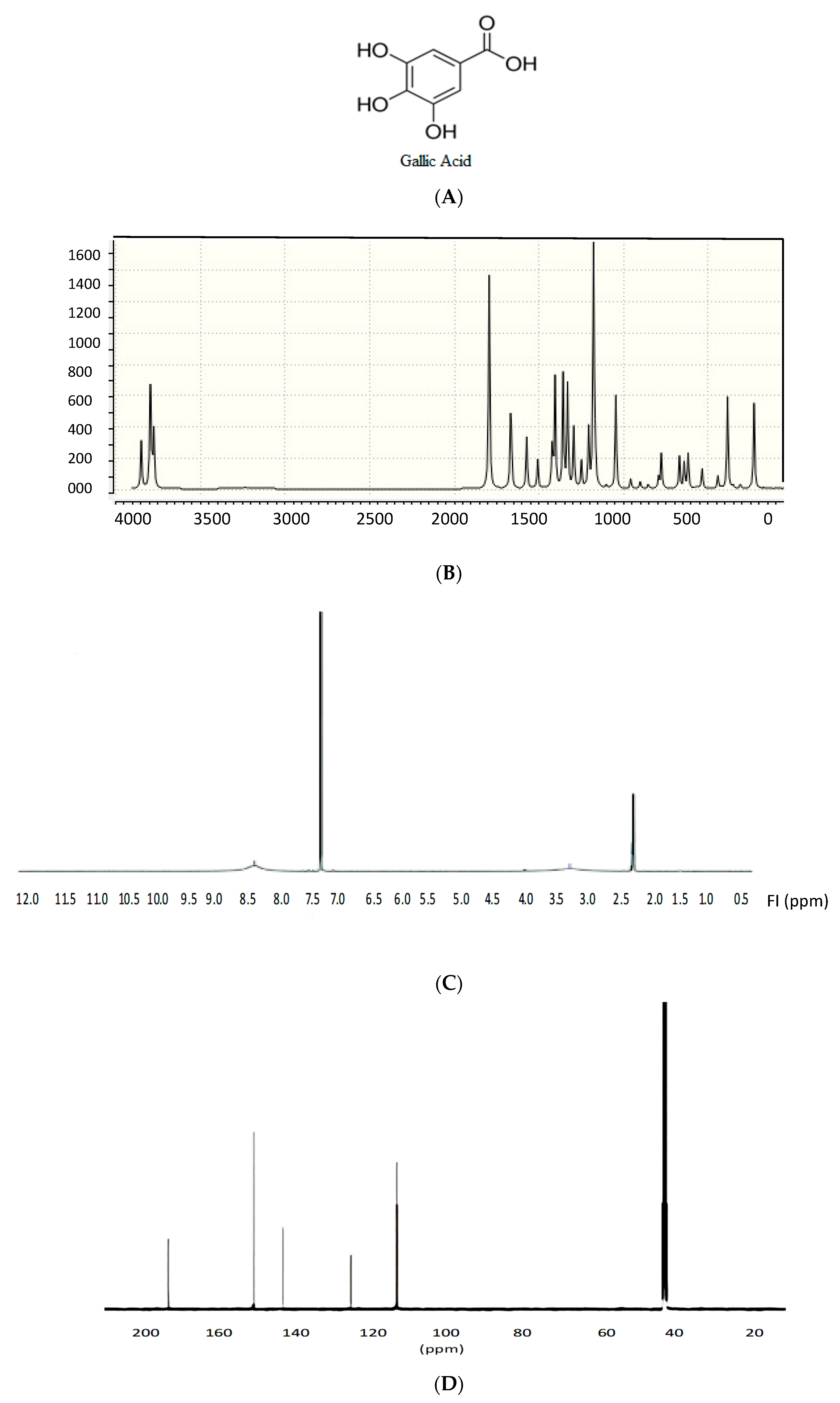
2.7. Identification of the Pure Compound
2.8. Antimicrobial Tests
2.8.1. Microbial Strains
2.8.2. Disc-Diffusion Assay
2.8.3. Micro-Well Dilution Assay of MIC and MBC
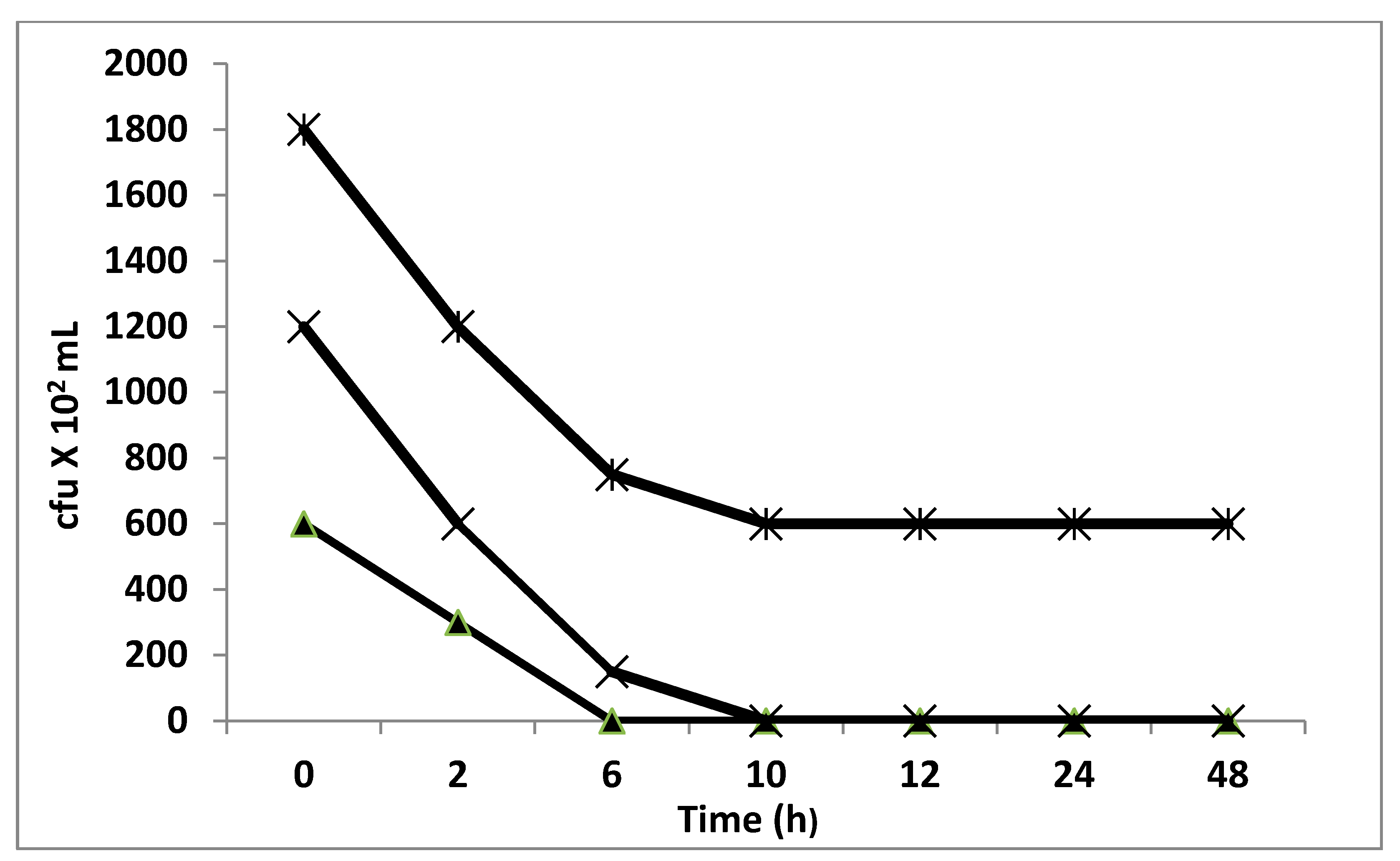
2.8.4. Time–Kill Studies
2.9. Antimutagenicity Assay (Ames Test)
2.10. Statistical Analysis
3. Results and Discussion
3.1. Determination of Total Phenolic and Flavonoid Contents of Date Pits
3.2. Extraction, Identification, and Elucidation of the Most Active Compound
3.3. Antimicrobial Activity of Date Pits
3.4. Antimutagenicity of Isolated Gallic Acid from Date Pits
4. Conclusion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Al-Farsi, M.; Lee, C.Y. Optimization of phenolics and dietary fiber extraction from date seeds. Food Chem. 2008, 108, 977–985. [Google Scholar] [CrossRef] [PubMed]
- Al-Shahib, W.; Marshal, R.J. The fruit of the date palm: It’s possible use as the best food for the future. Food Sci. Nutr. 2003, 54, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Besbes, S.; Blecker, C.; Deroanne, C.; Drira, N.; Attia, H. Date seeds Chemical composition and characteristic profiles of the lipid fraction. Food Chem. 2004, 84, 577–584. [Google Scholar] [CrossRef]
- Hussein, A.S.; Alhadrami, G.A.; Khalil, Y.H. The use of dates and date pits in broiler starter and finisher diets. Biores. Technol. 1998, 66, 219–223. [Google Scholar] [CrossRef]
- Hossain, M.Z.; Waly, M.I.; Singh, V.; Sequeira, V.; Rahman, M.S. Chemical Composition of Date-Pits and Its Potential for Developing Value-Added Product—A Review. Polish J. Food Nutrit. Sci. 2014, 4, 215–226. [Google Scholar] [CrossRef]
- Al–Farsi, M.; Alasalvar, C.; Al–Abid, M.; Al–Shoaily, K.; Al–Amry, M.; Al–Rawahy, F. Compositional and functional characteristics of dates, syrups and their by–products. Food Chem. 2007, 104, 943–947. [Google Scholar] [CrossRef]
- Waly, M.I.; Al–Ghafri, B.R.; Guizani, N.; Rahman, M.S. Phytonutrients effect of date pit extract against azoxymethane—Induced oxidative stress in rat colon. Asian Pacific J. Cancer Prev. 2015, 16, 3473–3477. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Al-Qarawi, A.A.; Mousa, H.M.; Ali, B.E.H.; Abdel-Rahman, H.; El-Mougy, A.A. Protective effect of extracts from dates (Phoenix dactylifera L.) on carbon tetrachloride—Induced hepatotoxicity in rats. Int. J. Appl. Res. Vet. Med. 2004, 2, 176–180. [Google Scholar]
- Mossa, J.S.; Hifnawy, M.S.; Mekkawi, A.G. Phytochemical and biological investigations on date seeds (Phoenix dactylifera L.) produced in Saudi Arabia. Arab. Gulf J. Sci Res. 1986, 4, 495–507. [Google Scholar]
- Saddiq, A.A.; Bawazir, A.E. Antimicrobial activity of date palm (Phoenix dactylifera) pits extracts and its role in reducing the side effect of methyl prednisolone on some neurotransmitter content in the brain. In Proceedings of the IV International Date Palm Conference, Abu Dhabi, UAE, 15 March 2010. [Google Scholar]
- Rajan, V.K.; Muraleedharan, K. A computational investigation on the structure, global parameters and antioxidant capacity of a polyphenol, Gallic acid. Food Chem. 2017, 220, 93–99. [Google Scholar] [CrossRef]
- Bai, J.; Zhang, Y.; Tang, C.; Hou, Y.; Ai, X.; Chen, X.; Zhang, Y.; Wang, X.; Meng, X. Gallic acid: Pharmacological activities and molecular mechanisms involved in inflammation-related diseases. Biomed. Pharm. 2021, 133, 110985. [Google Scholar] [CrossRef]
- Chanwitheesuk, A.; Teerawutgulrag, A.; Kilburn, J.D.; Rakariyatham, N. Antimicrobial gallic acid from Caesalpinia mimosoides Lamk. Food Chem. 2007, 100, 1044–1048. [Google Scholar] [CrossRef]
- Alu’datt, M.H.; Rababah, T.; Alhamad, M.N.; Al-Mahasneh, M.A.; Almajwal, A.; Gammoh, S.; Ereifej, K.; Johargy, A.; Alli, I. A review of phenolic compounds in oil-bearing plants: Distribution, identification and occurrence of phenolic compounds. Food Chem. 2017, 218, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Waterman, P.G.; Mole, S. Method in Ecology: Analysis of Phenolic Plant Metabolites; Blackwell Scientific Publishers: London, UK, 1994. [Google Scholar]
- Panizzi, L.; Caponi, C.; Catalano, S.; Cioni, P.L.; Morelli, I. In vitro antimicrobial activity of extracts and isolated constituents of Rubus ulmifolius. J. Ethnopharmacol. 2002, 79, 165–168. [Google Scholar] [CrossRef]
- Kroes, B.H.; van den Berg, A.J.; Quarles van Ufford, H.C.; van Dijk, H.; Labadie, R.P. Anti-inflammatory activity of gallic acid. Planta Med. 1992, 58, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Yang, M.; Wen, H. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug Anal. 2002, 10, 178–182. [Google Scholar]
- Velioglu, Y.S.; Mazza, G.; Gao, L.; Oomah, B.D. Antioxidant activity and total phenolics in selected fruits, vegetables and grain products. J. Agric. Food Chem. 1998, 46, 4113–4117. [Google Scholar] [CrossRef]
- Okla, M.K.; Abdel-Mawgoud, M.; Alamri, S.A.; Abbas, Z.K.; Al-Qahtani, W.H.; Al-Qahtani, S.M.; Al-Harbi, N.A.; Hassan, A.H.; Selim, S.; Alruhaili, M.H.; et al. Developmental Stages-Specific Response of Anise Plants to Laser-Induced Growth, Nutrients Accumulation, and Essential Oil Metabolism. Plants 2021, 10, 2591. [Google Scholar] [CrossRef]
- Selim, S.A.; El Alfy, S.; Al-Ruwaili, M.; Abdo, A.; Al Jaouni, S. Susceptibility of imipenem-resistant Pseudomonas aeruginosa to flavonoid glycosides of date palm (Phoenix dactylifera L.) tamar Growing in Al Madinah, Saudi Arabia. Afr. J. Biotech. 2012, 11, 416–422. [Google Scholar] [CrossRef]
- Maron, D.M.; Ames, B.N. Revised method for Salmonella mutagenicity test. Mutation Res. 1983, 113, 173–215. [Google Scholar] [CrossRef]
- Chen, H.J.; Ho, C.T. Antioxidant activities of caffeic acid and its related hydroxycinnamic acid compounds. J. Agri. Food Chem. 1997, 45, 2374–2378. [Google Scholar] [CrossRef]
- Cai, Y.Z.; Sun, M.; Xing, J.; Luo, Q.; Corke, H. Structure-radical scavenging activity relationship of phenolic compounds from traditional Chinese medicinal plants. Life Sci. 2006, 78, 2872–2888. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.M.; Wu, Y.C.; Chia, Y.C.; Chang, F.R.; Hsu, H.K.; Hsieh, Y.C.; Chen, C.C.; Yuan, S.S. Gallic acid, a major component of Toona sinensis leaf extracts, contains a ROS mediated anticancer activity in human prostate cancer cells. Cancer Lett. 2009, 286, 161–171. [Google Scholar] [CrossRef]
- Steenkamp, V.; Nkwane, O.; van Tonder, J.; Dinsmore, A.; Gulumian, M. Evaluation of the phenolic and flavonoid contents and radical scavenging activity of three southern African medicinal plants. Afr. J. Pharm. Pharmacol. 2013, 7, 703–709. [Google Scholar]
- Tunon, M.J.; Garcia-Mediavilla, M.V.; Sanchez-Campos, S.; Gonzales-Gallego, J. Potential of flavonoids as anti-inflammatory: Modulation of proinflammatory gene expression and signal transduction pathways. Curr. Drug Metab. 2009, 10, 256–271. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, H.; Fujii, K.; Yamasaki, O.; Oono, T.; Iwatsuki, K. Antibacterial action of several tannins against Staphylococcus aureus. J. Antimicrob. Chemother. 2001, 4, 487–491. [Google Scholar] [CrossRef] [PubMed]
- Penna, C.; Marino, S.; Vivot, E.; Cruañes, M.C.; de D Muñoz, J.; Cruañes, J.; Ferraro, G.; Gutkind, G.; Martino, V. Antimicrobial activity of Argentine plants used in the treatment of infectious diseases: Isolation of active compounds from Sebastiania brasiliensis. J. Ethnopharm. 2001, 77, 37–40. [Google Scholar] [CrossRef]
- Bhalodia Nayan, R.; Shukla, V.J. Antibacterial and antifungal activities from leaf extracts of Cassia Fistula L.: An Ethnomedicinal plant. J. Adv. Pharm. Technol. Res. 2011, 2, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Cowan, M.M. Plant Products as Antimicrobial Agents. Clin. Microbiology Rev. 1999, 12, 564–582. [Google Scholar] [CrossRef]
- Amensour, M.; Sendra, E.; Abrini, J.; Bouhdid, S.; Pérez-Alvarez, J.A.; Fernández-López, J. Total phenolic content and antioxidant activity of myrtle (Myrtus communis) extracts. Nat. Prod. Commun. 2009, 4, 819–824. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, D.; Khan, M.M.; Saeed, R. Comparative Analysis of Phenolics, Flavonoids, and Antioxidant and Antibacterial Potential of Methanolic, Hexanic and Aqueous Extracts from Adiantum caudatum Leaves. Antioxidants 2015, 4, 394–409. [Google Scholar] [CrossRef]
- Fukumoto, L.R.; Maza, G. Assessing antioxidant and prooxidant activities of phenolic compounds. J. Agric. Food Chem. 2000, 48, 3597–3604. [Google Scholar] [CrossRef] [PubMed]
- Puupponen-Pimia, R.; Nohynek, L.; Alakomi, H.L.; Oksman-Caldentey, K.M. Bioactive berry compounds novel tools against human pathogens. Appl. Microbiol. Biotechnol. 2005, 67, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Cushnie, T.P.T.; Lamb, A.J. Antimicrobial activity of flavonoids. Int. J. Antimicrob. Agents 2005, 26, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Provan, G.J.; Helliwell, K. Tea flavonoids: Their functions, utilization and analysis. Trends Food Sci. Technol. 2000, 11, 152–160. [Google Scholar] [CrossRef]
- Sanchez-Maldonado, A.F.; Schieber, A.; Ganzle, M.G. Structure-function relationships of the antibacterial activity of phenolic acids and their metabolism by lactic acid bacteria. J. Appl. Microbiol. 2011, 111, 1176–1184. [Google Scholar] [CrossRef]
- Campos, F.M.; Couto, J.A.; Figueiredo, A.R.; Tóth, I.V.; Rangel, A.O.S.S.; Hogg, T.A. Cell membrane damage induced by phenolic acids on wine lactic acid bacteria. Int. J. Food Microbiol. 2009, 135, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Ramel, C.; Alekperov, B.N.; Ames, T.; Kada, L.W. Wattenberg, Inhibitors of mutagenesis and their relevance to carcinogenesis. Mutat. Res. 1986, 168, 47–65. [Google Scholar] [CrossRef]
- Hung, Y.H.; Wang, Y.J.; Chou, C.C. Antimutagenic activity of Aspergillus awamori-fermented black soyabean response to stimulated digestive juice treatments and its antimutagenic mechanisms. LWT-Food Sci. Technol. 2009, 42, 56–62. [Google Scholar] [CrossRef]
- Ames, B.N.; McCann, J.; Yamasaki, E. Methods for detecting carcinogens and mutagens with the Salmonella/mammalian microsome mutagenicity test. Mutat. Res. 1975, 31, 347–364. [Google Scholar] [CrossRef]
- Negi, P.S.; Jayaprakasha, G.K.; Jena, B.S. Antioxidant and antimutagenic activities of pomegranate peel extracts. Food Chem. 2003, 80, 393–397. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Compounds | Concentration |
|---|---|
| Hydroxybenzoic acids | |
| Syringic acid | 0.62 ± 0.015 |
| Gallic acid Pyrogallol Quinol | 11.85 ± 0.25 1.870 ± 0.12 0.354 ± 0.04 |
| Hydroxycinnamic acids | |
| Ferulic acid | 3.6 ± 0.78 |
| p-Coumaric acid | 5.58 ± 1.68 |
| Vanillic acid | 0.92 ± 0.75 |
| Caffeic acid | 0.05 ± 0.0023 |
| Total Phenolic a | 24.84 ± 0.19 |
| Flavones | |
| Apigenin | 0.198 ± 0.90 |
| Luteolin | 1.52 ± 0.13 |
| Flavonols | |
| Myricetin | 0.65 ± 0.007 |
| Quercetin | 2.986 ± 0.135 |
| Total Flavonoid b | 5.324 ± 0.069 |
| Microorganisms | IZD a | MIC | MBC |
|---|---|---|---|
| Streptococcus sp | 13 | 500 | 500 |
| Staphylococcus aureus | 22 | 250 | 250 |
| Serratia marcescens | – | – | – |
| Escherichia coli | 11 | 500 | 500 |
| Klebsiella pneumoniae | 11 | 500 | 500 |
| Proteus vulgaris | – | – | – |
| Pseudomonas aeruginosa | – | – | – |
| Candida albicans | 10 | 500 | 500 |
| Compound | Concentration (μg/mL) | Inhibition % of Revertants | |
|---|---|---|---|
| TA98 | TA100 | ||
| Gallic acid | 3000 | 52 | 39 |
| 1700 | 43 | 27 | |
| 700 | 38 | 21 | |
| 500 | 32 | 15 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Selim, S.; Abdel-Mawgoud, M.; Al-sharary, T.; Almuhayawi, M.S.; Alruhaili, M.H.; Al Jaouni, S.K.; Warrad, M.; Mohamed, H.S.; Akhtar, N.; AbdElgawad, H. Pits of Date Palm: Bioactive Composition, Antibacterial Activity and Antimutagenicity Potentials. Agronomy 2022, 12, 54. https://doi.org/10.3390/agronomy12010054
Selim S, Abdel-Mawgoud M, Al-sharary T, Almuhayawi MS, Alruhaili MH, Al Jaouni SK, Warrad M, Mohamed HS, Akhtar N, AbdElgawad H. Pits of Date Palm: Bioactive Composition, Antibacterial Activity and Antimutagenicity Potentials. Agronomy. 2022; 12(1):54. https://doi.org/10.3390/agronomy12010054
Chicago/Turabian StyleSelim, Samy, Mohamed Abdel-Mawgoud, Tarak Al-sharary, Mohammed S. Almuhayawi, Mohammed H. Alruhaili, Soad K. Al Jaouni, Mona Warrad, Hussein S. Mohamed, Nosheen Akhtar, and Hamada AbdElgawad. 2022. "Pits of Date Palm: Bioactive Composition, Antibacterial Activity and Antimutagenicity Potentials" Agronomy 12, no. 1: 54. https://doi.org/10.3390/agronomy12010054
APA StyleSelim, S., Abdel-Mawgoud, M., Al-sharary, T., Almuhayawi, M. S., Alruhaili, M. H., Al Jaouni, S. K., Warrad, M., Mohamed, H. S., Akhtar, N., & AbdElgawad, H. (2022). Pits of Date Palm: Bioactive Composition, Antibacterial Activity and Antimutagenicity Potentials. Agronomy, 12(1), 54. https://doi.org/10.3390/agronomy12010054