
Effects of Melatonin Treatment on Sweet Cherry Tree Yield and Fruit Quality


 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Quality Parameters
2.3. Total Phenolic and Anthocyanin Quantification
2.4. Statistical Analysis
3. Results
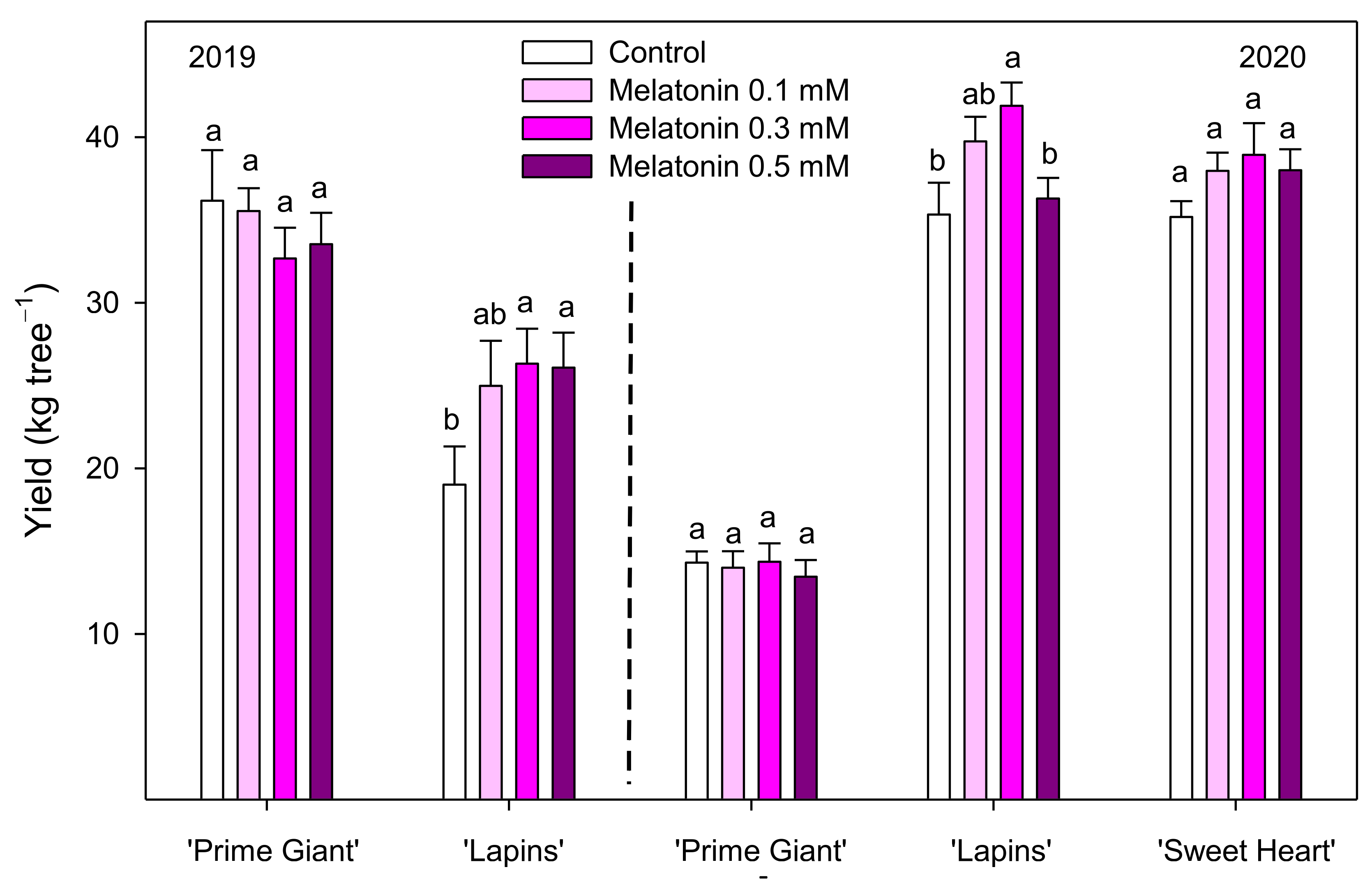
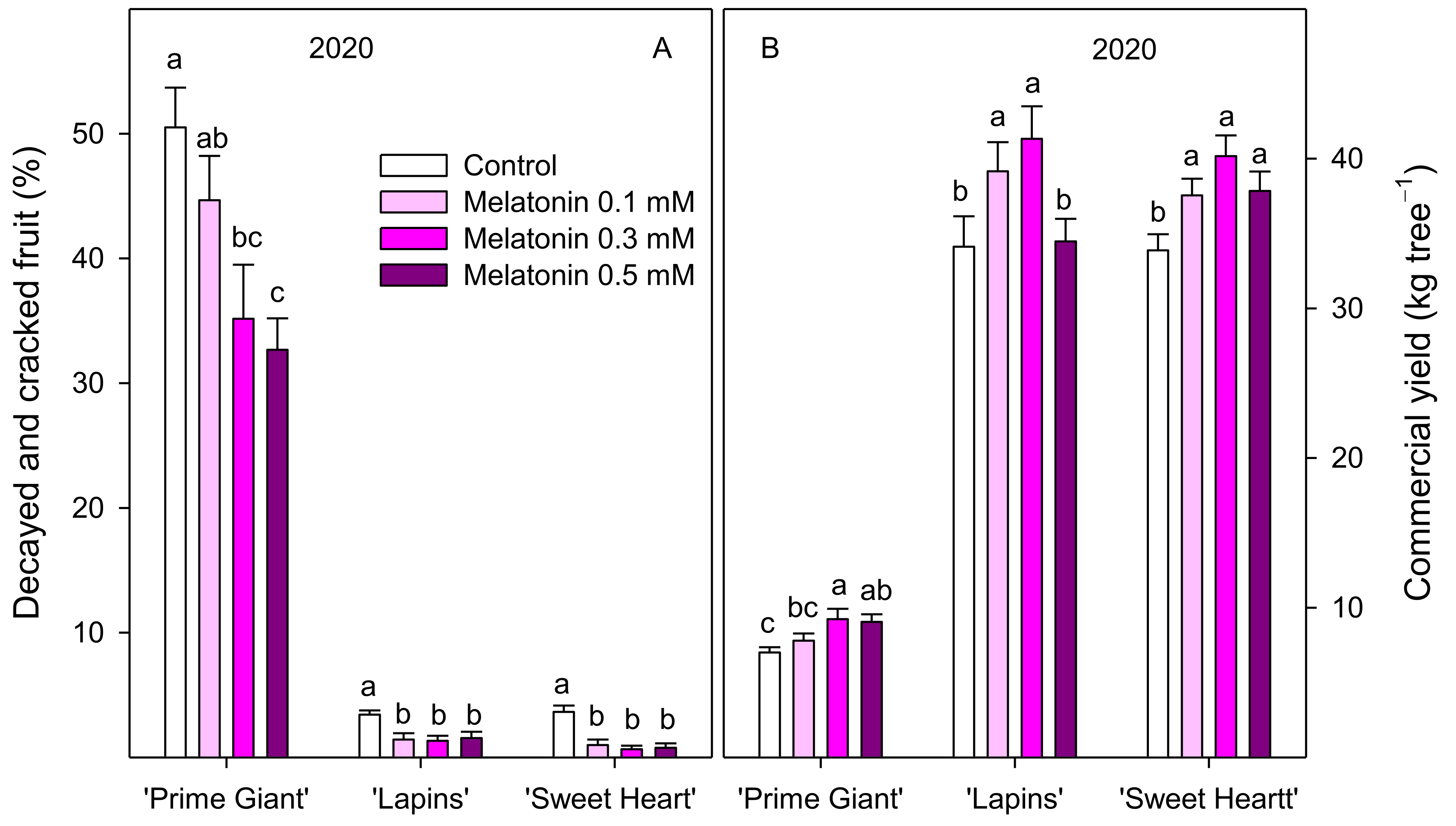
3.1. Crop Yield and Fruit Weight
3.2. Fruit Quality Parameters
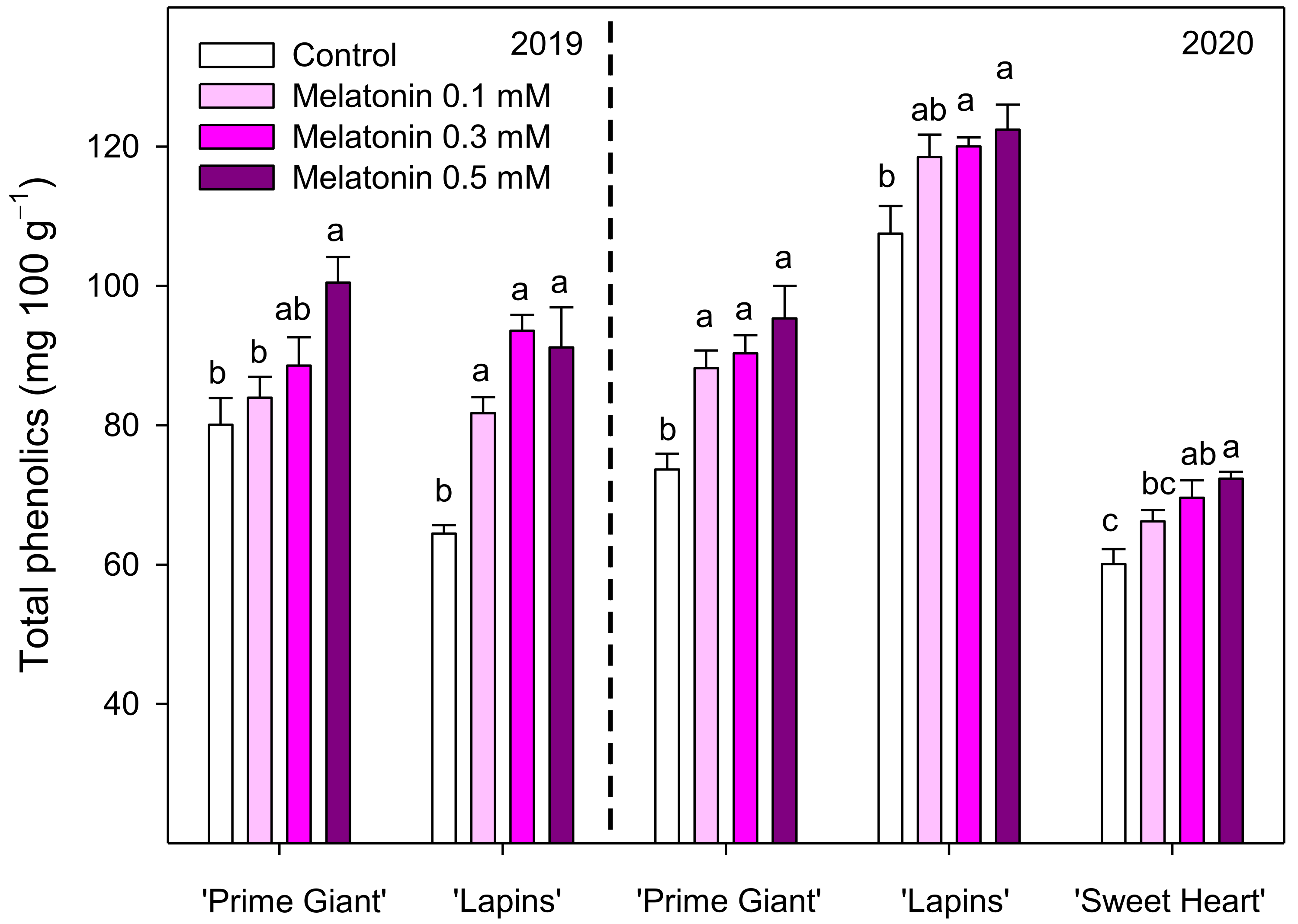
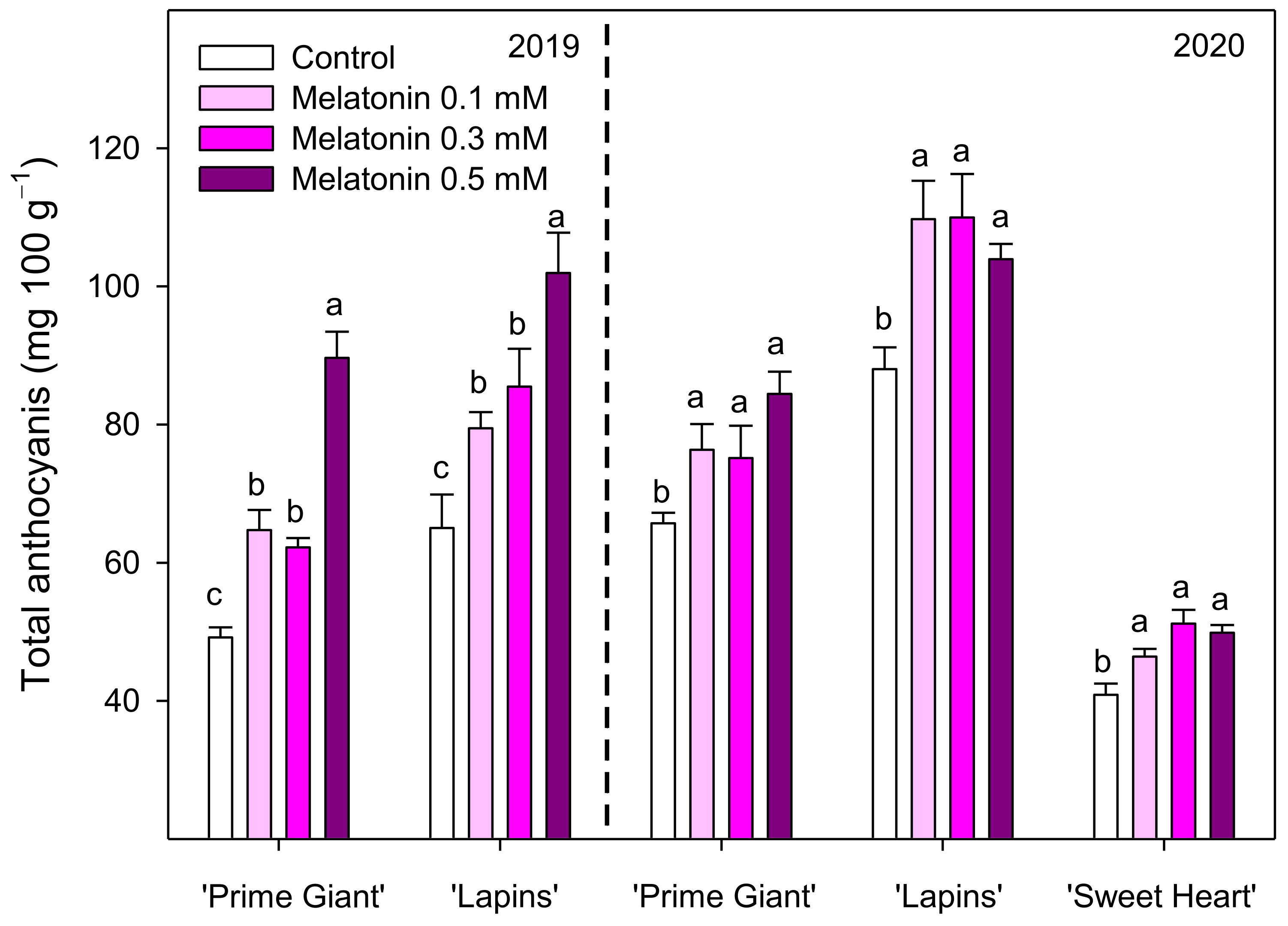
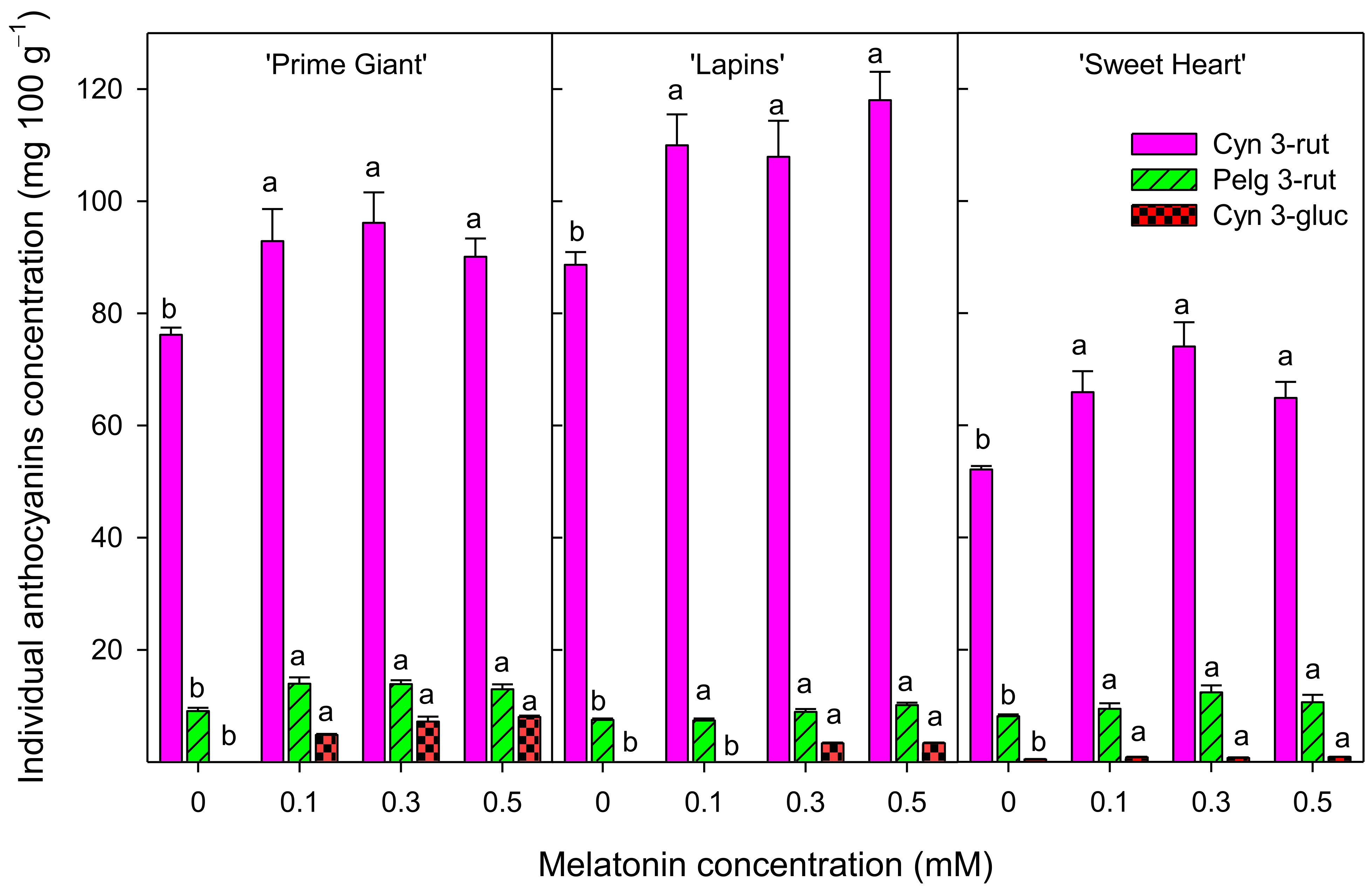
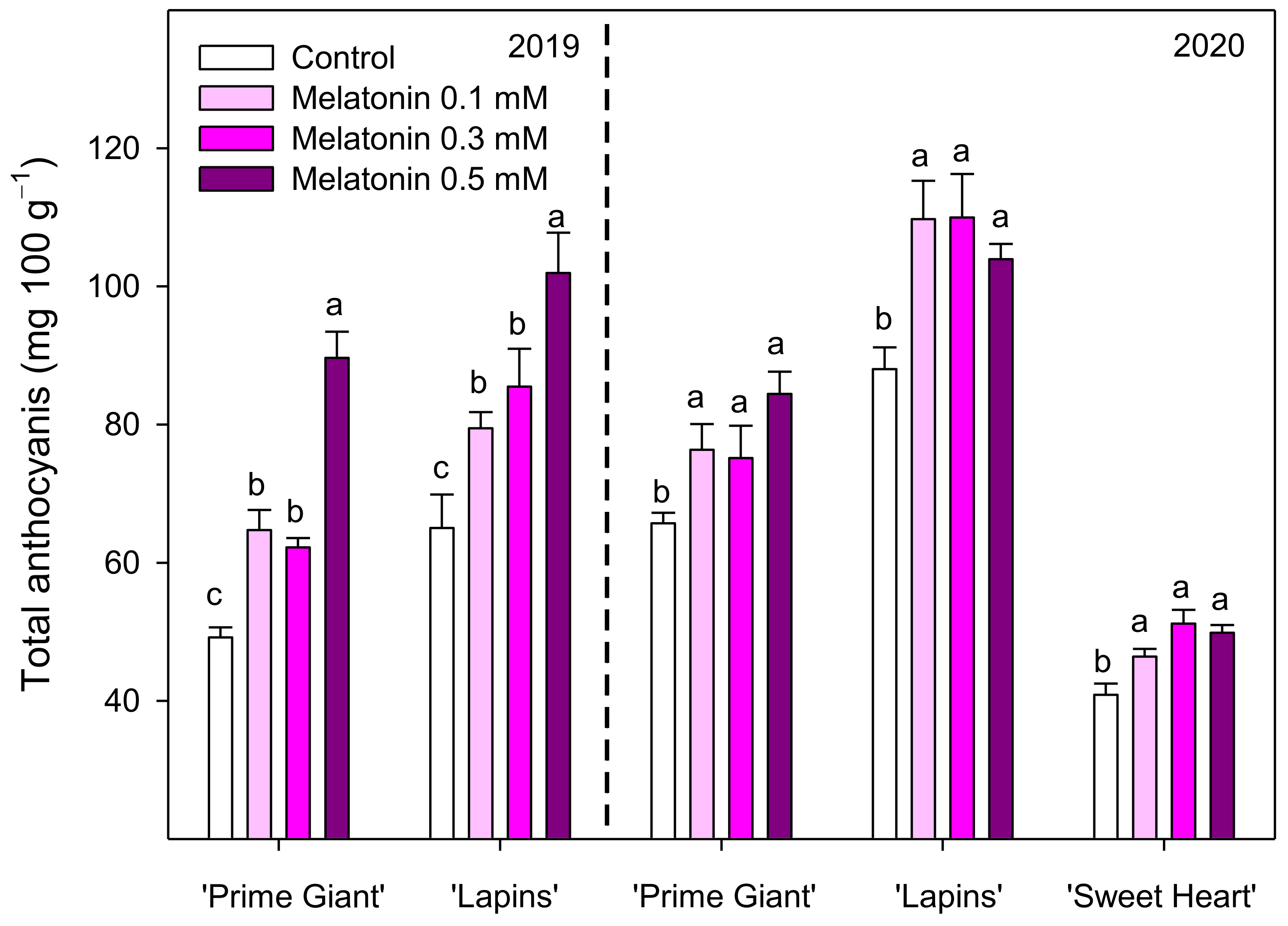
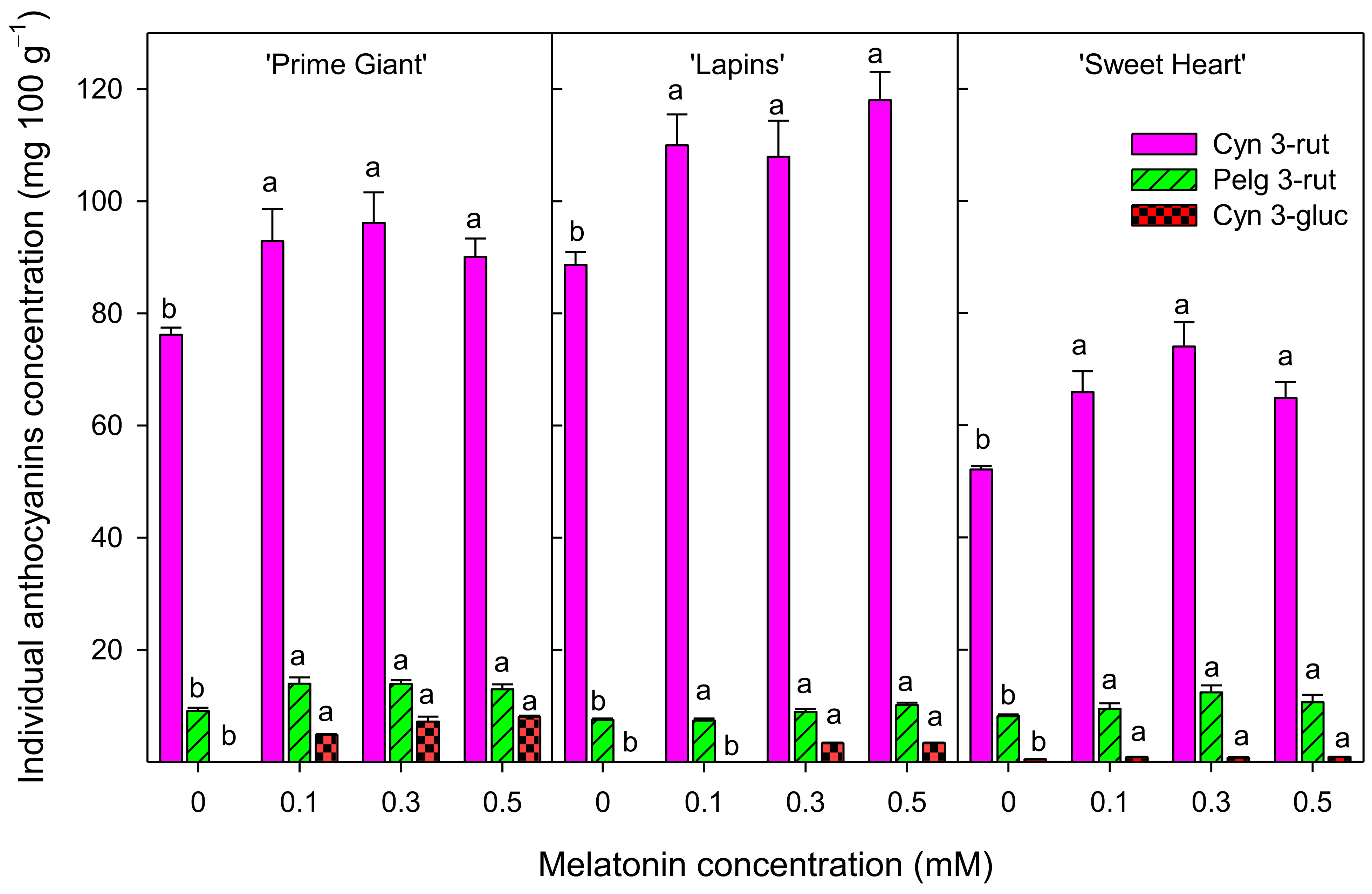
3.3. Total Phenolics and Total and Individual Anthocyanins
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Díaz-Mula, H.M.; Castillo, S.; Martínez-Romero, D.; Valero, D.; Zapata, P.J.; Guillén, F.; Serrano, M. Sensory, nutritive and functional properties of sweet cherry as affected by cultivar and ripening stage. Food Sci. Technol. Int. 2009, 15, 535–543. [Google Scholar] [CrossRef]
- Usenik, V.; Fabčič, J.; Štampar, F. Sugars, organic acids, phenolic composition and antioxidant activity of sweet cherry (Prunus avium L.). Food Chem. 2008, 107, 185–192. [Google Scholar] [CrossRef]
- Martínez-Esplá, A.; Zapata, P.J.; Valero, D.; García-Viguera, C.; Castillo, S.; Serrano, M. Preharvest application of oxalic acid increased fruit size, bioactive compounds, and antioxidant capacity in sweet cherry cultivars (Prunus avium L.). J. Agric. Food Chem. 2014, 62, 3432–3437. [Google Scholar] [CrossRef] [PubMed]
- McCune, L.M.; Kubota, C.; Stendell-Hollins, N.R.; Thomson, C.A. Cherries and health: A review. Crit. Rev. Food Sci. Nutr. 2011, 51, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Blando, F.; Oomah, B.D. Sweet and sour cherries: Origin, distribution, nutritional composition and health benefits. Trends Food Sci. Technol. 2019, 86, 517–529. [Google Scholar] [CrossRef]
- Faienza, M.F.; Corbo, F.; Carocci, A.; Catalano, A.; Clodoveo, M.L.; Grano, M.; Wang, D.Q.H.; D’Amato, G.; Muraglia, M.; Franchini, C.; et al. Novel insights in health-promoting properties of sweet cherries. J. Funct. Foods 2020, 69, 103945. [Google Scholar] [CrossRef]
- FAOSTAT. Statistical Databases On Global Food Production and Trade. Food and Agriculture Organization. 2019. Available online: https://www.fao.org/statistics/en/ (accessed on 3 November 2021).
- Dubbels, R.; Reiter, R.J.; Klenke, E.; Goebel, A.; Schnakenberg, E.; Ehleers, C.; Schiwara, H.W.; Schloot, W. Melatonin in edible plants identified by radioimmunoassay and by high performance liquid chromatography-mass spectrometry. J. Pineal Res. 1995, 18, 28–31. [Google Scholar] [CrossRef]
- Shi, H.; Chen, K.; Wei, Y.; He, C. Fundamental issues of melatonin-mediated stress signalling in plants. Front. Plant Sci. 2016, 7, 1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnao, M.B.; Hernández-Ruiz, J. Is phytomelatonin a new plant hormone? Agronomy 2020, 10, 95. [Google Scholar] [CrossRef] [Green Version]
- Xu, T.; Chen, Y.; Kang, H. Melatonin is a potential target for improving post-harvest preservation of fruits and vegetables. Front. Plant Sci. 2019, 10, 1388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin in flowering, fruit set and fruit ripening. Plant Reprod. 2020, 33, 77–87. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, R.; Sun, Y.; Liu, Z.; Jin, W.; Sun, Y. The beneficial effects of exogenous melatonin on tomato fruit properties. Sci. Hortic. 2016, 207, 14–20. [Google Scholar] [CrossRef]
- Abd El-Naby, S.K.M.A.; Mohamed, A.A.A.; El-Naggar, Y.I.M. Effect of melatonin, GA3 and NAA on vegetative growth, yield and quality of ‘Canino’ apricot fruits. Acta Sci. Pol. Hortorum Cultus 2019, 18, 167–174. [Google Scholar] [CrossRef]
- Medina-Santamarina, J.; Zapata, P.J.; Valverde, J.M.; Valero, D.; Serrano, M.; Guillén, F. Melatonin treatment of apricot trees leads to maintenance of fruit quality attributes during storage at chilling and non-chilling temperatures. Agronomy 2021, 11, 917. [Google Scholar] [CrossRef]
- Lorente-Mento, J.M.; Guillén, F.; Castillo, S.; Martínez-Romero, D.; Valverde, J.M.; Valero, D.; Serrano, M. Melatonin treatment to pomegranate trees enhances fruit bioactive compounds and quality traits at harvest and during postharvest storage. Antioxidants 2021, 10, 820. [Google Scholar] [CrossRef]
- Sharafi, Y.; Jannatizadeh, A.; Fard, J.R.; Aghdam, M.S. Melatonin treatment delays senescence and improves antioxidant potential of sweet cherry fruits during cold storage. Sci. Hortic. 2021, 288, 110304. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, X.; Yang, Q.; Zhao, Q. Exogenous melatonin delays postharvest fruit senescence and maintains the quality of sweet cherries. Food Chem. 2019, 301, 125311. [Google Scholar] [CrossRef]
- Miranda, S.; Vilches, P.; Suazo, M.; Pavez, L.; García, K.; Méndez, M.; González, M.; Meisel, L.; Defilippi, B.G.; Del Pozo, T. Melatonin triggers metabolic and gene expression changes making possible improved quality traits of two sweet cherry cultivars during cold storage. Food Chem. 2020, 319, 126360. [Google Scholar] [CrossRef]
- Tijero, V.; Muñoz, P.; Munné-Bosch, S. Melatonin as an inhibitor of sweet cherries ripening in orchard trees. Plant Physiol. Biochem. 2019, 140, 88–95. [Google Scholar] [CrossRef]
- Michailidis, M.; Tanou, G.; Sarrou, E.; Karagiannis, E.; Ganopoulos, I.; Martens, S.; Molassiotis, A. Pre- and post-harvest melatonin application boosted phenolic compounds accumulation and altered respiratory characters in sweet cherry fruit. Front. Nutr. 2021, 8, 695061. [Google Scholar] [CrossRef] [PubMed]
- Medina-Santamarina, J.; Serrano, M.; Lorente-Mento, J.M.; García-Pastor, M.E.; Zapata, P.J.; Valero, D.; Guillén, F. Melatonin treatment of pomegranate trees increases crop yield and quality parameters at harvest and during storage. Agronomy 2021, 11, 861. [Google Scholar] [CrossRef]
- Ibrahim, M.F.M.; Abd Elbar, O.H.; Farag, R.; Hikal, M.; El-Kelish, A.; El-Yazied, A.A.; Alkahtani, J.; Abd El-Gawad, H.G. Melatonin counteracts drought induced oxidative damage and stimulates growth, productivity and fruit quality properties of tomato plants. Plants 2020, 9, 1276. [Google Scholar] [CrossRef]
- Debnath, B.; Hussain, M.; Li, M.; Lu, X.; Sun, Y.; Qiu, D. Exogenous melatonin improves fruit quality features, health promoting antioxidant compounds and yield traits in tomato fruits under acid rain stress. Molecules 2018, 23, 1868. [Google Scholar] [CrossRef] [Green Version]
- Debnath, B.; Islam, W.; Li, M.; Sun, Y.T.; Lu, X.C.; Mitra, S.; Hussain, M.; Liu, S.; Qiu, D. Melatonin mediates enhancement of stress tolerance in plants. Int. J. Mol. Sci. 2019, 20, 1040. [Google Scholar] [CrossRef] [Green Version]
- Correia, S.; Queirós, F.; Ferreira, H.; Morais, M.C.; Afonso, S.; Silva, A.P.; Gonçalves, B. Foliar application of calcium and growth regulators modulate sweet cherry (Prunus avium L.) tree performance. Plants 2020, 9, 410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, J.F.; Xu, T.F.; Song, C.Z.; Yu, Y.; Hu, F.; Zhang, L.; Zhang, Z.W.; Xi, Z.M. Melatonin treatment of pre-veraison grape berries to increase size and synchronicity of berries and modify wine aroma components. Food Chem. 2015, 185, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Drake, S.R.; Elfving, D.C. Indicators of maturity and storage quality of ‘Lapins’ sweet cherry. HortTechnology 2002, 12, 687–690. [Google Scholar] [CrossRef]
- Serrano, M.; Guillén, F.; Martínez-Romero, D.; Castillo, S.; Valero, D. Chemical constituents and antioxidant activity of sweet cherry at different ripening stages. J. Agric. Food Chem. 2005, 53, 2741–2745. [Google Scholar] [CrossRef]
- Valero, D.; Serrano, M. Postharvest Biology and Technology for Preserving Fruit Quality; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar] [CrossRef]
- Xia, H.; Shen, Y.; Shen, T.; Wang, X.; Zhang, X.; Hu, P.; Liang, D.; Lin, L.; Deng, Q.; Lv, X. Melatonin accumulation in sweet cherry and its influence on fruit quality and antioxidant properties. Molecules 2020, 25, 753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ze, Y.; Gao, H.; Li, T.; Yang, B.; Jiang, Y. Insights into the roles of melatonin in maintaining quality and extending shelf life of postharvest fruits. Trends Food Sci. Technol. 2021, 109, 569–578. [Google Scholar] [CrossRef]
- Correia, S.; Schouten, R.; Silva, A.P.; Gonçalves, B. Factors affecting quality and health promoting compounds during growth and postharvest life of sweet cherry (Prunus avium L.). Front. Plant Sci. 2017, 8, 2166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonçalves, A.C.; Bento, C.; Jesus, F.; Alves, G.; Silva, L.R. Sweet cherry phenolic compounds: Identification, characterization, and health benefits. Stud. Nat. Prod. Chem. 2018, 59, 31–78. [Google Scholar] [CrossRef]
- Gonçalves, A.C.; Bento, C.; Silva, B.; Simões, M.; Silva, L.R. Nutrients, bioactive compounds and bioactivity: The health benefits of sweet cherries (Prunus avium L.). Curr. Nutr. Food Sci. 2019, 15, 208–227. [Google Scholar] [CrossRef]
- Sureda, A.; Bibiloni, M.M.; Julibert, A.; Bouzas, C.; Argelich, E.; Llompart, I.; Pons, A.; Tur, J.A. Adherence to the Mediterranean diet and inflammatory markers. Nutrients 2018, 10, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarangon, E.; Moreno, J.J. Polyphenols and taste 2 receptors. Physiological, pathophysiological and pharmacological implications. Biochem. Pharmacol. 2020, 78, 114086. [Google Scholar] [CrossRef]
- Luo, J.; Si, H.; Jia, Z.; Liu, D. Dietary anti-aging polyphenols and potential mechanisms. Antioxidants 2021, 10, 283. [Google Scholar] [CrossRef]
- Antognoni, F.; Potente, G.; Mandrioli, R.; Angeloni, C.; Freschi, M.; Malaguti, M.; Hrelia, S.; Lugli, S.; Gennari, F.; Muzzi, E.; et al. Fruit quality characterization of new sweet cherry cultivars as a good source of bioactive phenolic compounds with antioxidant and neuroprotective potential. Antioxidants 2020, 9, 677. [Google Scholar] [CrossRef] [PubMed]
- Mozetič, B.; Simčič, M.; Trebše, P. Anthocyanins and hidroxycinnamic acids of Lambert Compact cherries (Prunus avium L.) after cold storage and 1-methylcyclorpoene treatments. Food Chem. 2006, 97, 302–309. [Google Scholar] [CrossRef]
- Gonçalves, B.; Silva, A.P.; Moutinho-Pereia, J.; Bacelar, E.; Rosa, E.; Meyer, A.S. Effect of ripeness and postharvest storage on the evolution of color and anthocyanins in cherries (Prunus avium L.). Food Chem. 2007, 103, 976–984. [Google Scholar] [CrossRef]
- Serrano, M.; Díaz-Mula, H.; Zapata, P.J.; Castillo, S.; Guillén, F.; Martínez-Romero, D.; Valverde, J.M.; Valero, D. Maturity stage at harvest determines the fruit quality and antioxidant potential after storage of sweet cherry cultivars. J. Agric. Food Chem. 2009, 57, 3240–3246. [Google Scholar] [CrossRef]
- Sharma, M.; Jacob, J.K.; Subramanian, J.; Paliyath, G. Hexanal and 1-MCP treatments for enhancing the shelf life and quality of sweet cherry (Prunus avium L.). Sci. Hortic. 2010, 125, 239–247. [Google Scholar] [CrossRef]
- Bal, E. Physicochemical changes in ‘Santa Rosa’ plum fruit treated with melatonin during cold storage. J. Food Meas. Charact. 2019, 13, 1713–1720. [Google Scholar] [CrossRef]
- Liu, C.H.; Zheng, H.H.; Sheng, K.L.; Liu, W.; Zheng, L. Effects of melatonin treatment on the postharvest quality of strawberry fruit. Postharvest Biol. Technol. 2018, 139, 47–55. [Google Scholar] [CrossRef]
- Bal, E. Effect of melatonin treatments on biochemical quality and postharvest life of nectarines. J. Food Meas. Charact. 2021, 15, 288–295. [Google Scholar] [CrossRef]
- Zhang, Y.; Huber, D.J.; Hu, M.; Jiang, G.; Gao, Z.; Xu, X.; Jiang, Y.; Zhang, Z. Delay of postharvest browning in litchi fruit by melatonin via the enhancing of antioxidative processes and oxidation repair. J. Agric. Food Chem. 2018, 66, 7475–7484. [Google Scholar] [CrossRef]
- Aghdam, M.S.; Luo, Z.; Li, L.; Jannatizadeh, A.; Fard, J.R.; Pirzad, F. Melatonin treatment maintains nutraceutical properties of pomegranate fruits during cold storage. Food Chem. 2020, 303, 125385. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Cultivar | T1 | T2 | T3 | Harvest |
|---|---|---|---|---|---|
| 2019 | ‘Prime Giant’ | 4 May | 22 May | 8 June | 11 June |
| ‘Lapins’ | 5 May | 1 June | 17 June | 21 June | |
| 2020 | ‘Prime Giant’ | 23 April | 15 May | 30 May | 4 June |
| ‘Lapins’ | 24 April | 26 May | 13 June | 17 June | |
| ‘Sweet heart’ | 28 April | 1 June | 27 June | 2 July |
| Melatonin Concentration | |||||
|---|---|---|---|---|---|
| Cultivar | Year | Control | 0.1 mM | 0.3 mM | 0.5 mM |
| Fruit Colour (a*/b*) | |||||
| ‘Prime Giant’ | 2019 | 3.20 ± 0.07c | 3.47 ± 0.07b | 3.47 ± 0.06b | 3.92 ± 0.06a |
| ‘Prime Giant’ | 2020 | 3.12 ± 0.09c | 3.37 ± 0.11b | 3.50 ± 0.07b | 3.72 ± 0.11a |
| ‘Lapins’ | 2019 | 4.25 ± 0.07c | 4.55 ± 0.07b | 4.50 ± 0.07b | 4.76 ± 0.07a |
| ‘Lapins’ | 2020 | 4.33 ± 0.11b | 4.77 ± 0.14a | 4.73 ± 0.09a | 4.68 ± 0.09a |
| ‘Sweet Heart’ | 2020 | 3.13 ± 0.17c | 3.43 ± 0.11b | 3.59 ± 0.03b | 3.79 ± 0.07a |
| Fruit firmness (N mm−1) | |||||
| ‘Prime Giant’ | 2019 | 1.59 ± 0.04c | 1.83 ± 0.04b | 2.02 ± 0.04a | 1.91 ± 0.03a |
| ‘Prime Giant’ | 2020 | 1.49 ± 0.08b | 1.84 ± 0.09a | 1.99 ± 0.06a | 1.78 ± 0.10a |
| ‘Lapins’ | 2019 | 1.56 ± 0.05b | 1.73 ± 0.04a | 1.80 ± 0.04a | 1.77 ± 0.04a |
| ‘Lapins’ | 2020 | 1.60 ± 0.05a | 1.83 ± 0.06a | 1.81 ± 0.04a | 1.80 ± 0.04a |
| ‘Sweet Heart’ | 2020 | 1.54 ± 0.07b | 1.87 ± 0.06a | 2.05 ± 0.09a | 1.85 ± 0.05a |
| Total soluble solids (g 100 g−1) | |||||
| ‘Prime Giant’ | 2019 | 20.43 ± 0.24c | 22.45 ± 0.12b | 23.00 ± 0.12ab | 23.47 ± 0.41a |
| ‘Prime Giant’ | 2020 | 22.28 ± 0.22b | 23.93 ± 0.45a | 23.77 ± 0.23a | 24.27 ± 0.17a |
| ‘Lapins’ | 2019 | 20.36 ± 0.14b | 21.40 ± 0.26a | 22.10 ± 0.47a | 22.17 ± 0.16a |
| ‘Lapins’ | 2020 | 18.98 ± 0.12c | 20.07 ± 0.06b | 20.52 ± 0.22ab | 21.05 ± 0.08a |
| ‘Sweet Heart’ | 2020 | 19.70 ± 0.21b | 20.92 ± 0.07a | 20.75 ± 0.16a | 20.72 ± 0.38a |
| Titratable acidity (g 100 g−1) | |||||
| ‘Prime Giant’ | 2019 | 1.10 ± 0.01b | 1.31 ± 0.02a | 1.24 ± 0.03a | 1.22 ± 0.01a |
| ‘Prime Giant’ | 2020 | 1.25 ± 0.01b | 1.40 ± 0.02a | 1.39 ± 0.02a | 1.36 ± 0.03a |
| ‘Lapins’ | 2019 | 0.95 ± 0.01b | 1.15 ± 0.03a | 1.12 ± 0.01a | 1.09 ± 0.02a |
| ‘Lapins’ | 2020 | 1.08 ± 0.02b | 1.21 ± 0.01a | 1.18 ± 0.01a | 1.17 ± 0.01a |
| ‘Sweet Heart’ | 2020 | 1.22 ± 0.01b | 1.37 ± 0.04a | 1.35 ± 0.02a | 1.36 ± 0.03a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carrión-Antolí, A.; Lorente-Mento, J.M.; Valverde, J.M.; Castillo, S.; Valero, D.; Serrano, M. Effects of Melatonin Treatment on Sweet Cherry Tree Yield and Fruit Quality. Agronomy 2022, 12, 3. https://doi.org/10.3390/agronomy12010003
Carrión-Antolí A, Lorente-Mento JM, Valverde JM, Castillo S, Valero D, Serrano M. Effects of Melatonin Treatment on Sweet Cherry Tree Yield and Fruit Quality. Agronomy. 2022; 12(1):3. https://doi.org/10.3390/agronomy12010003
Chicago/Turabian StyleCarrión-Antolí, Alberto, José Manuel Lorente-Mento, Juan Miguel Valverde, Salvador Castillo, Daniel Valero, and María Serrano. 2022. "Effects of Melatonin Treatment on Sweet Cherry Tree Yield and Fruit Quality" Agronomy 12, no. 1: 3. https://doi.org/10.3390/agronomy12010003
APA StyleCarrión-Antolí, A., Lorente-Mento, J. M., Valverde, J. M., Castillo, S., Valero, D., & Serrano, M. (2022). Effects of Melatonin Treatment on Sweet Cherry Tree Yield and Fruit Quality. Agronomy, 12(1), 3. https://doi.org/10.3390/agronomy12010003