
Ferulago campestris Essential Oil as Active Ingredient in Chitosan Seed-Coating: Chemical Analyses, Allelopathic Effects, and Protective Activity against the Common Bean Pest Acanthoscelides obtectus

 ,
,  ,
,  ,
,  ,
,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plant Material and EO Hydrodistillation
2.2. Gas Chromatography–Mass Spectrometry Analyses and Peaks Identification
2.3. Antigerminative Test
2.4. Determination of Hydrogen Peroxide Concentration
2.5. Determination of Lipid Peroxidation
2.6. Insect Rearing
2.7. Behavioral Assays
2.8. Chitosan and Chitosan–Essential Oil Coating
2.9. Environmental Scanning Electronic Microscope Investigations and Measurements
2.10. Oviposition-Repellent Activity of CH–EO of F. campestris
2.11. Effects of F. campestris EO on Phaseolus Vulgaris Germination
2.12. Data Analysis
3. Results and Discussion
3.1. Essential Oil Composition
3.2. Antigerminative Activity
3.3. Behavioral Assays
3.4. Environmental Scanning Electronic Microscope Investigations and Measurements
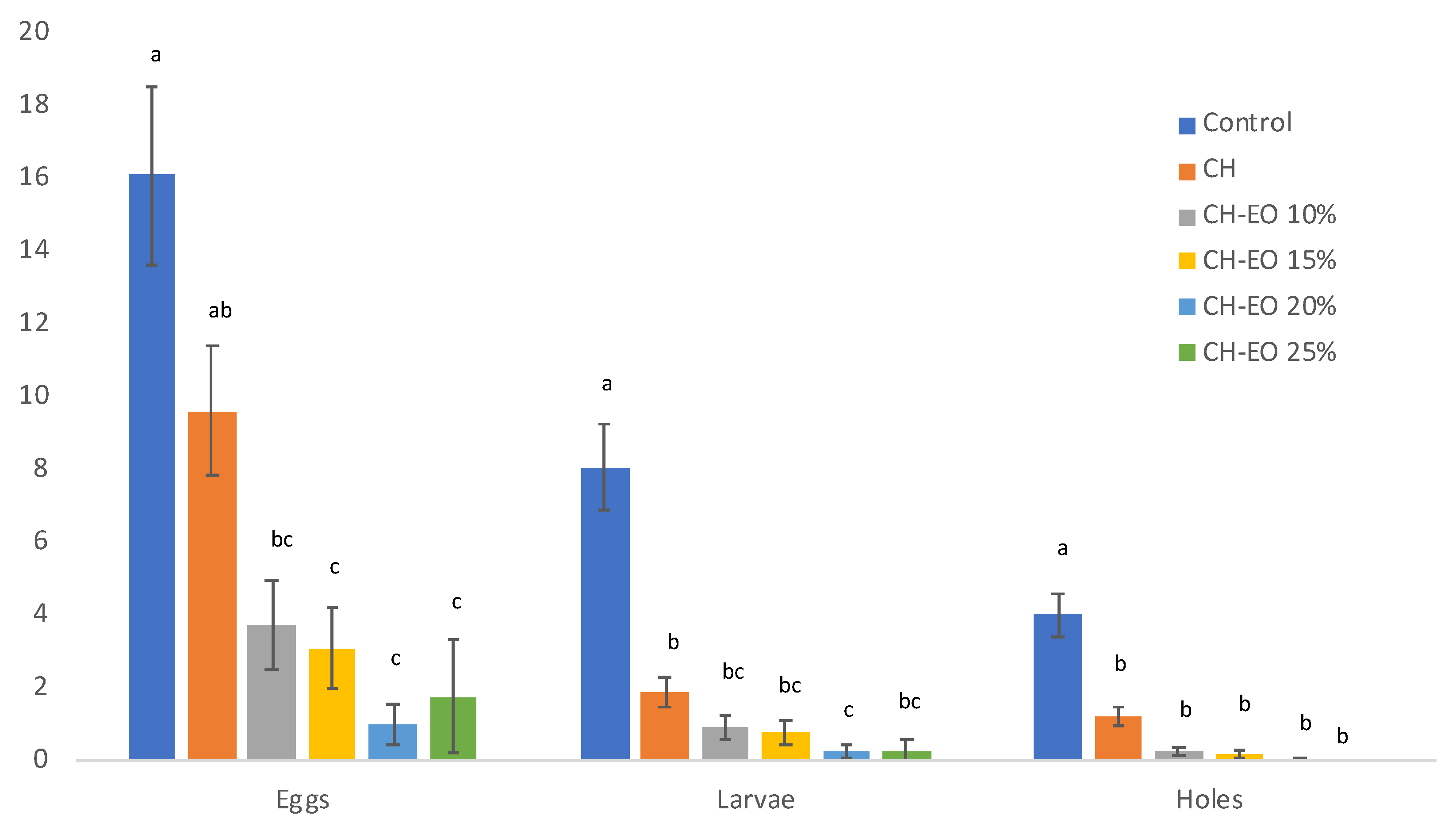
3.5. Chitosan Coating Protection Test of P. vulgaris Beans
3.6. Effects of Chitosan–F. campestris EO on P. vulgaris Germination
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- The Plant List Ferulago campestris (Besser) Grecescu. The Plant List. Available online: http://www.theplantlist.org/tpl1.1/record/kew-2808738 (accessed on 1 April 2020).
- Cannon, J. Flora Europaea; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar]
- Maggi, F.; Tirillini, B.; Papa, F.; Sagratini, G.; Vittori, S.; Cresci, A.; Comam, M.M.; Cecchini, C. Chemical composition and antimicrobial activity of the essential oil of Ferulago campestris (Besser) Grecescu growing in central Italy. Flavour. Frag. J. 2009, 24, 309–315. [Google Scholar] [CrossRef]
- Cecchini, C.; Coman, M.M.; Cresci, A.; Tirillini, B.; Cristalli, G.; Papa, F.; Sagratini, G.; Vittori, S.; Maggi, F. Essential oil from fruits and roots of Ferulago campestris (Besser) Grecescu (Apiaceae): Composition and antioxidant and anti-Candida activity. Flavour Frag. J. 2010, 25, 493–502. [Google Scholar] [CrossRef]
- Riela, S.; Bruno, M.; Rosselli, S.; Saladino, M.L.; Caponetti, E.; Formisano, C.; Senatore, F. A study on the essential oil of Ferulago campestris: How much does extraction method influence the oil composition? J. Sep. Sci. 2011, 34, 483–492. [Google Scholar] [CrossRef] [Green Version]
- Sabbieti, M.G.; Agas, D.; Maggi, F.; Vittori, S.; Marchetti, L. Molecular mediators involved in Ferulago campestris essential oil effects on osteoblast metabolism. J. Cell Biochem. 2011, 112, 3742–3754. [Google Scholar] [CrossRef]
- Li, W.; Papa, F.; Shi, J.; Maggi, F.; Chen, X. The Chemical Constituents and the Hepato-protective Effect of the Essential Oil of Ferulago campestris (Besser) Grecescu (Apiaceae). J. Essent. 2016, 19, 1701–1708. [Google Scholar]
- Conti, B.; Canale, A.; Bertoli, A.; Gozzini, F.; Pistelli, L. Essential oil composition and larvicidal activity of six Mediterranean aromatic plants against the mosquito Aedes albopictus (Diptera: Culicidae). Parasitol. Res. 2010, 107, 1455–1461. [Google Scholar] [CrossRef] [PubMed]
- Bedini, S.; Bougherra, H.H.; Flamini, G.; Cosci, F.; Belhamel, K.; Ascrizzi, R.; Conti, B. Repellency of anethole-and estragole-type fennel essential oils against stored grain pests: The different twins. Bull. Insectol. 2016, 69, 149–157. [Google Scholar]
- Ghasemi, V.; Moharramipour, S.; Tahmasbi, G. Biological activity of some plant essential oils against Varroa destructor (Acari: Varroidae), an ectoparasitic mite of Apis mellifera (Hymenoptera: Apidae). Exp. Appl. Acarol. 2011, 55, 147–154. [Google Scholar] [CrossRef]
- Muturi, E.J.; Ramirez, J.L.; Doll, K.M.; Bowman, M.J. Combined toxicity of three essential oils against Aedes aegypti (Diptera: Culicidae) larvae. J. Med. Entomol. 2017, 54, 1684–1691. [Google Scholar] [CrossRef] [PubMed]
- Yeom, H.J.; Kang, J.S.; Kim, G.H.; Park, I.K. Insecticidal and Acetylcholine Esterase Inhibition Activity of Apiaceae Plant Essential Oils and Their Constituents against Adults of German Cockroach (Blattella germanica). J. Agric. Food Chem. 2012, 60, 7194–7203. [Google Scholar] [CrossRef]
- Traboulsi, A.F.; El-Haj, S.; Tueni, M.; Taoubi, K.; Nader, N.A.; Mrad, A. Repellency and toxicity of aromatic plant extracts against the mosquito Culex pipiens molestus (Diptera: Culicidae). Pest Manag. Sci. 2005, 61, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Paul, U.V.; Lossini, J.S.; Edwards, P.J.; Hilbeck, A. Effectiveness of products from four locally grown plants for the management of Acanthoscelides obtectus (Say) and Zabrotes subfasciatus (Boheman) (both Coleoptera: Bruchidae) in stored beans under laboratory and farm conditions in Northern Tanzania. J. Stored Prod.Res. 2009, 45, 97–107. [Google Scholar] [CrossRef]
- Baier, A.H.; Webster, B.D. Control of Acanthoscelides obtectus Say (Coleoptera: Bruchidae) in Phaseolus vulgaris L. seed stored on small farms-II. Germination and cooking time. J. Stored Prod. Res. 1992, 28, 295–299. [Google Scholar] [CrossRef]
- Mallqui, K.S.V.; Oliveira, E.E.; Guedes, R.N.C. Competition between the bean weevils Acanthoscelides obtectus and Zabrotes subfasciatus in common beans. J. Stored Prod. Res. 2013, 55, 32–35. [Google Scholar] [CrossRef]
- Gołębiowski, M.; Maliński, E.; Nawrot, J.; Stepnowski, P. Identification and characterization of surface lipid components of the dried-bean beetle Acanthoscelides obtectus (Say) (Coleoptera: Bruchidae). J. Stored Prod. Res. 2008, 44, 386–388. [Google Scholar] [CrossRef]
- Oliveira, M.R.D.; Bonome, L.T.D.S.; Bittencourt, H.V.H.; Zarowni, E.; Lefchak, L.D.S. Alternative treatments in bean seeds for repelling Acanthoscelides obtectus (SAY). Res. J. Seed Sci. 2018, 40, 362–369. [Google Scholar] [CrossRef] [Green Version]
- Girolami, V.; Mazzon, L.; Squartini, A.; Mori, N.; Marzaro, M.; Di Bernardo, A.; Greatti, M.; Giorio, C.; Tapparo, A. Translocation of neonicotinoid insecticides from coated seeds to seedling guttation drops: A novel way of intoxication for bees. J. Econ. Entomol. 2009, 102, 1808–1815. [Google Scholar] [CrossRef]
- Badawy, M.E.; El-Aswad, A.F. Insecticidal activity of chitosans of different molecular weights and chitosan-metal complexes against cotton leafworm Spodoptera littoralis and oleander aphid Aphis nerii. Plant Protect Sci. 2012, 48, 131–141. [Google Scholar] [CrossRef] [Green Version]
- Satyal, P.; Paudel, P.; Poudel, A.; Dosoky, N.S.; Moriarity, D.M.; Vogler, B.; Setzer, W.N. Chemical compositions, phytotoxicity, and biological activities of Acorus calamus essential oils from Nepal. Nat. Prod. Commun. 2013, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Ricci, D.; Epifano, F.; Fraternale, D. The essential oil of Monarda didyma L.(Lamiaceae) exerts phytotoxic activity in vitro against various weed seed. Molecules 2017, 22, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matoušková, M.; Jurová, J.; Gruľová, D.; Wajs-Bonikowska, A.; Renčo, M.; Sedlák, V.; Poracovà, J.; Gogaľová, Z.; Kalemba, D. Phytotoxic effect of invasive Heracleum mantegazzianum essential oil on dicot and monocot species. Molecules 2019, 24, 425. [Google Scholar] [CrossRef] [Green Version]
- Mutlu, S.; Atici, Ö.; Esim, N.; Mete, E. Essential oils of catmint (Nepeta meyeri Benth.) induce oxidative stress in early seedlings of various weed species. Acta Physiol. Plant 2011, 33, 943–951. [Google Scholar] [CrossRef]
- Ogunwande, I.A.; Avoseh, O.N.; Olasunkanmi, K.N.; Lawal, O.A.; Ascrizzi, R.; Flamini, G. Chemical composition, anti-nociceptive and anti-inflammatory activities of essential oil of Bougainvillea glabra. J. Ethnopharmacol. 2019, 232, 188–192. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry; Allured Publishing Corporation: Carol Stream, IL, USA, 2007; p. 456. [Google Scholar]
- Romani, R.; Bedini, S.; Salerno, G.; Ascrizzi, R.; Flamini, G.; Echeverria, M.C.; Farina, P.; Conti, B. Andean flora as a source of new repellents against insect pests: Behavioral, morphological and electrophysiological studies on Sitophilus zeamais (coleoptera: Curculionidae). Insects 2019, 10, 171. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Li, Y. Combined effects of two kinds of essential oils on physical, mechanical and structural properties of chitosan films. Food Hydrocoll. 2014, 36, 287–293. [Google Scholar] [CrossRef]
- Nahdy, M.S. Bean sieving, a possible control measure for the dried bean beetles, Acanthoscelides obtectus (Say) (Coleoptera: Bruchidae). J. Stored Prod. Res. 1994, 30, 65–69. [Google Scholar] [CrossRef]
- Lombardi, T.; Bedini, S.; Bertacchi, A. Germination ecology of the aromatic halophyte Artemisia caerulescens L.: Influence of abiotic factors and seed after-ripening time. Folia Geobot. 2019, 54, 115–124. [Google Scholar] [CrossRef]
- Hsiung, Y.C.; Chen, Y.A.; Chen, S.Y.; Chi, W.C.; Lee, R.H.; Chiang, T.Y.; Huang, H.J. Volatilized myrcene inhibits growth and activates defense responses in rice roots. Acta Physiol. Plant 2013, 35, 2475–2482. [Google Scholar] [CrossRef]
- Scrivanti, L.R.; Zunino, M.P.; Zygadlo, J.A. Tagetes minuta and Schinus areira essential oils as allelopathic agents. Biochem. Syst. Ecol. 2003, 31, 563–572. [Google Scholar] [CrossRef]
- Singh, H.P.; Batish, D.R.; Kaur, S.; Arora, K.; Kohli, R.K. α-Pinene inhibits growth and induces oxidative stress in roots. Ann. Bot. 2006, 98, 1261–1269. [Google Scholar] [CrossRef] [Green Version]
- Chowhan, N.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Phytotoxic effects of β-pinene on early growth and associated biochemical changes in rice. Acta Physiol. Plant. 2011, 33, 2369–2376. [Google Scholar] [CrossRef]
- De Feo, V.; De Simone, F.; Senatore, F. Potential allelochemicals from the essential oil of Ruta graveolens. Phytochemistry 2002, 61, 573–578. [Google Scholar] [CrossRef]
- Martino, L.D.; Mancini, E.; Almeida, L.F.R.D.; Feo, V.D. The antigerminative activity of twenty-seven monoterpenes. Molecules 2010, 15, 6630–6637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amri, I.; Gargouri, S.; Hamrouni, L.; Hanana, M.; Fezzani, T.; Jamoussi, B. Chemical composition, phytotoxic and antifungal activities of Pinus pinea essential oil. J. Pest. Sci. 2012, 85, 199–207. [Google Scholar] [CrossRef]
- Papachristos, D.P.; Stamopoulos, D.C. Repellent, toxic and reproduction inhibitory effects of essential oil vapours on Acanthoscelides obtectus (Say) (Coleoptera: Bruchidae). J. Stored Prod. Res. 2002, 38, 117–128. [Google Scholar] [CrossRef]
- Jumbo, L.O.V.; Faroni, L.R.; Oliveira, E.E.; Pimentel, M.A.; Silva, G.N. Potential use of clove and cinnamon essential oils to control the bean weevil, Acanthoscelides obtectus Say, in small storage units. Ind. Crops Prod. 2014, 56, 27–34. [Google Scholar] [CrossRef]
- Chaudhari, A.K.; Singh, V.K.; Kedia, A.; Das, S.; Dubey, N.K. Essential oils and their bioactive compounds as eco-friendly novel green pesticides for management of storage insect pests: Prospects and retrospects. Environ. Sci. Pollut. Res. 2021, 28, 18918–18940. [Google Scholar] [CrossRef]
- Angelini, L.G.; Carpanese, G.; Cioni, P.L.; Morelli, I.; Macchia, M.; Flamini, G. Essential oils from Mediterranean Lamiaceae as weed germination inhibitors. J. Agric. Food Chem. 2003, 51, 6158–6164. [Google Scholar] [CrossRef] [PubMed]
- De Almeida, L.F.R.; Frei, F.; Mancini, E.; De Martino, L.; De Feo, V. Phytotoxic activities of Mediterranean essential oils. Molecules 2010, 15, 4309–4323. [Google Scholar] [CrossRef] [Green Version]
- Turek, C.; Stintzing, F.C. Stability of essential oils: A review. Compr. Rev. Food Sci. Food Saf. 2013, 12, 40–53. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | l.r.i. a | Relative Abundance (%) |
|---|---|---|
| tricyclene | 928 | tr b |
| α-thujene | 931 | 1.1 |
| α-pinene | 941 | 19.7 |
| α-fenchene | 954 | tr |
| camphene | 954 | 0.8 |
| thuja-2,4(10)-diene | 959 | tr |
| sabinene | 976 | 1.8 |
| β-pinene | 982 | 2.8 |
| myrcene | 993 | 26.1 |
| α-phellandrene | 1005 | 2.8 |
| δ-3-carene | 1011 | 5.9 |
| α-terpinene | 1018 | 0.7 |
| p-cymene | 1027 | 2.6 |
| limonene | 1032 | 6.6 |
| (Z)-β-ocimene | 1042 | 3.5 |
| (E)-β-ocimene | 1052 | 2.2 |
| γ-terpinene | 1062 | 18.5 |
| terpinolene | 1088 | 1.6 |
| 1,3,8-p-menthatriene | 1113 | tr |
| 4-terpineol | 1178 | tr |
| α-terpineol | 1189 | tr |
| trans-carveol | 1218 | tr |
| citronellol | 1230 | tr |
| cis-chrysanthenyl acetate | 1264 | 1.9 |
| n-decanol | 1272 | tr |
| bornyl acetate | 1287 | tr |
| perilla alcohol | 1295 | tr |
| trans-pinocarvyl acetate | 1297 | tr |
| δ-elemene | 1340 | 0.1 |
| α-copaene | 1376 | tr |
| β-elemene | 1392 | tr |
| β-caryophyllene | 1420 | 0.1 |
| β-copaene | 1429 | tr |
| α-humulene | 1456 | tr |
| germacrene D | 1478 | tr |
| valencene | 1492 | 0.1 |
| bicyclogermacrene | 1496 | tr |
| α-muurolene | 1498 | tr |
| δ-cadinene | 1524 | tr |
| elemol | 1549 | tr |
| germacrene B | 1554 | tr |
| (E)-nerolidol | 1565 | tr |
| spathulenol | 1576 | tr |
| caryophyllene oxide | 1581 | tr |
| humulene epoxide II | 1607 | tr |
| neointermedeol | 1660 | tr |
| nootkatone | 1803 | 0.1 |
| m-camphorene | 1960 | 0.3 |
| p-camphorene | 1995 | 0.1 |
| Monoterpene hydrocarbons | 96.6 | |
| Oxygenated monoterpenes | 1.9 | |
| Sesquiterpene hydrocarbons | 0.4 | |
| Oxygenated sesquiterpenes | 0.1 | |
| Diterpene hydrocarbons | 0.5 | |
| Non-terpene derivatives | tr | |
| Total identified (%): | 99.4 | |
| Number of Seeds Germinated of the 10 Used for Each Sample a | ||||||
|---|---|---|---|---|---|---|
| Plant | Sample/Conc. | 0.06 μg/mL | 0.125 μg/mL | 0.250 μg/mL | 0.625 μg/mL | 1.250 μg/mL |
| P. rhoeas | EO | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 8.3 ± 0.5 | 7.6 ± 0.5 |
| PCM | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 9.5 ± 0.4 | 8.0 ± 0.8 | |
| Myrcene/26.1% | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 8.0 ± 1.4 | |
| α-pinene/19.7% | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 8.3 ± 1.7 | |
| γ-terpinene/18.5% | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | |
| Pendimethalin | 0.0 ± 0.0 | |||||
| Control (H2O) | 10.0 ± 0.0 | |||||
| T. campylodes | EO | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 7.6 ± 1.2 | 8.6 ± 1.2 |
| PCM | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 8.6 ± 0.4 | 8.3 ± 0.9 | |
| Myrcene/26.1% | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 8.3 ± 1.2 | 8.0 ± 1.4 | |
| α-pinene/19.7% | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 8.3 ± 0.4 | 8.6 ± 1.2 | |
| γ-terpinene/18.5% | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | |
| Pendimethalin | 0.0 ± 0.0 | |||||
| Control (H2O) | 10.0 ± 0.0 | |||||
| P. annua | EO | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 6.6 ± 0.9 * | 6.0 ± 0.8 * |
| PCM | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 7.3 ± 0.4 * | 6.3 ± 0.4 ** | |
| Myrcene/26.1% | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 7.6 ± 1.2 | 7.3 ± 0.4 * | |
| α-pinene/19.7% | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 6.6 ± 0.4 ** | 6.6 ± 0.9 * | |
| γ-terpinene/18.5% | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | |
| Pendimethalin | 0.0 ± 0.0 | |||||
| Control (H2O) | 10.0 ± 0.0 | |||||
| S. verticillata | EO | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 6.6 ± 0.4 ** | 5.6 ± 0.4 ** |
| PCM | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 7.0 ± 0.8 * | 6.6 ± 0.4 ** | |
| Myrcene/26.1% | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 7.3 ± 0.4 * | 7.0 ± 0.8 * | |
| α-pinene/19.7% | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 6.6 ± 1.2 | 6.3 ± 1.2 | |
| γ-terpinene/18.5% | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | |
| Pendimethalin | 0.0 ± 0.0 | |||||
| Control (H2O) | 10.0 ± 0.0 | |||||
| P. vulgaris | EO | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 |
| PCM | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | |
| Myrcene/26.1% | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | |
| α-pinene/19.7% | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | |
| γ-terpinene/18.5% | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | 10.0 ± 0.0 | |
| Pendimethalin | 0.0 ± 0.0 | |||||
| Control (H2O) | 10.0 ± 0.0 | |||||
| Radicle Elongation (cm) | ||||||
|---|---|---|---|---|---|---|
| Plant | Sample/Conc. | 0.06 μg/mL | 0.125 μg/mL | 0.250 μg/mL | 0.625 μg/mL | 1.250 μg/mL |
| P. rhoeas | EO | 1.50 ± 0.09 | 1.46 ± 0.10 | 1.28 ± 0.11 ** | 1.08 ± 0.07 ** | 0.84 ± 0.10 ** |
| PCM | 1.52 ± 0.07 | 1.44 ± 0.10 | 1.32 ± 0.07 * | 1.00 ± 0.09 ** | 0.82 ± 0.18 | |
| Myrcene/26.1% | 1.54 ± 0.05 | 1.34 ± 0.20 | 1.24 ± 0.10 ** | 0.96 ± 0.08 ** | 0.80 ± 0.06 | |
| α-pinene/19.7% | 1.48 ± 0.07 | 1.42 ± 0.12 | 1.12 ± 0.11 * | 0.92 ± 0.07 ** | 0.74 ± 0.16 | |
| γ-terpinene/18.5% | 1.58 ± 0.04 | 1.56 ± 0.05 | 1.54 ± 0.05 | 1.55 ± 0.05 | 1.56 ± 0.04 | |
| Pendimethalin | 0.0 ± 0.0 | |||||
| Control (H2O) | 1.61 ± 0.20 | |||||
| T. campylodes | EO | 2.16 ± 0.12 | 2.02 ± 0.13 | 1.92 ± 0.17 ** | 1.36 ± 0.08 ** | 0.96 ± 0.10 ** |
| PCM | 2.10 ± 0.11 | 1.98 ± 0.04 | 1.92 ± 0.11 ** | 1.28 ± 0.11 ** | 0.98 ± 0.12 ** | |
| Myrcene/26.1% | 2.12 ± 0.14 | 1.94 ± 0.07 | 1.86 ± 0.08 ** | 1.34 ± 0.08 ** | 0.88 ± 0.10 ** | |
| α-pinene/19.7% | 2.06 ± 0.08 | 1.96 ± 0.08 | 1.92 ± 0.07 ** | 1.26 ± 0.10 ** | 0.90 ± 0.11 ** | |
| γ-terpinene/18.5% | 2.42 ± 0.07 | 2.38 ± 0.42 | 2.42 ± 0.07 ** | 2.36 ± 0.05 ** | 2.30 ± 0.06 | |
| Pendimethalin | 0.0 ± 0.0 | |||||
| Control (H2O) | 2.48 ± 0.21 | |||||
| P. annua | EO | 1.08 ± 0.07 | 0.94 ± 0.08 | 0.52 ± 0.07 ** | 0.36 ± 0.08 ** | 0.16 ± 0.08 ** |
| PCM | 1.02 ± 0.14 | 0.82 ± 0.16 | 0.62 ± 0.07 ** | 0.32 ± 0.11 ** | 0.14 ± 0.05 ** | |
| Myrcene/26.1% | 0.90 ± 0.12 | 0.86 ± 0.05 | 0.58 ± 0.11 ** | 0.42 ± 0.07 ** | 0.18 ± 0.07 ** | |
| α-pinene/19.7% | 1.06 ± 0.13 | 0.91 ± 0.16 | 0.58 ± 0.07 ** | 0.36 ± 0.10 * | 0.18 ± 0.04 ** | |
| γ-terpinene/18.5% | 1.12 ± 0.11 | 0.96 ± 0.08 | 1.08 ± 0.07 | 1.00 ± 0.06 | 0.96 ± 0.10 * | |
| Pendimethalin | 0.0 ± 0.0 | |||||
| Control (H2O) | 1.31 ± 0.21 | |||||
| S. verticilata | EO | 1.42 ± 0.15 | 0.98 ± 0.07 ** | 0.76 ± 0.21 * | 0.34 ± 0.10 ** | 0.13 ± 0.04 ** |
| PCM | 1.42 ± 0.14 | 0.91 ± 0.14 * | 0.71 ± 0.27 * | 0.41 ± 0.10 ** | 0.13 ± 0.02 ** | |
| Myrcene/26.1% | 1.44 ± 0.13 | 0.94 ± 0.08 * | 0.78 ± 0.13 ** | 0.36 ± 0.10 ** | 0.15 ± 0.01 ** | |
| α-pinene/19.7% | 1.46 ± 0.13 | 0.88 ± 0.12 * | 0.76 ± 0.18 ** | 0.32 ± 0.13 ** | 0.12 ± 0.01 ** | |
| γ-terpinene/18.5% | 1.54 ± 0.08 | 1.24 ± 0.22 | 1.28 ± 0.21 | 1.28 ± 0.22 | 1.52 ± 0.30 | |
| Pendimethalin | 0.0 ± 0.0 | |||||
| Control (H2O) | 1.60 ± 0.31 | |||||
| P. vulgaris | EO | 2.04 ± 0.10 | 2.03 ± 0.01 | 2.16 ± 0.12 | 2.14 ± 0.06 | 2.14 ± 0.04 |
| PCM | 2.01 ± 0.16 | 2.07 ± 0.07 | 2.07 ± 0.05 | 2.05 ± 0.07 | 2.20 ± 0.06 | |
| Myrcene/26.1% | 2.06 ± 0.08 | 2.09 ± 0.03 | 2.05 ± 0.05 | 2.08 ± 0.06 | 2.16 ± 0.04 | |
| α-pinene/19.7% | 2.02 ± 0.14 | 2.08 ± 0.05 | 2.05 ± 0.06 | 2.15 ± 0.03 | 2.06 ± 0.03 | |
| γ-terpinene/18.5% | 2.06 ± 0.08 | 2.10 ± 0.05 | 2.13 ± 0.05 | 2.12 ± 0.06 | 2.08 ± 0.06 | |
| Pendimethalin | 0.00 ± 0.00 | |||||
| Control (H2O) | 2.20 ± 0.20 | |||||
| H2O2 (ng/g FW) | |||||
| P. rhoeas | T. campylodes | P. annua | S. verticillata | P. vulgaris | |
| CTRL | 144.66 ± 4.51 | 155.66 ± 7.51 | 213.26 ± 5.71 | 209.90 ± 7.32 | 180.47 ± 4.91 |
| FEO | 312.66 ± 6.23 * | 451.00 ± 6.16 * | 412.66 ± 6.23 * | 419.69 ± 12.48 * | 182.33 ± 5.24 |
| PCM myrc. α-pin. | 314.33 ± 6.94 * 250.08 ± 2.72 * 209.34 ± 2.54 * | 437.23 ± 4.48 * 283.06 ± 6.09 * 211.83 ± 5.21 * | 416.66 ± 3.85 * 336.42 ± 5.21 * 229.36 ± 4.15 * | 421.52 ± 3.68 * 317.55 ± 5.13 * 284.41 ± 4.36 * | 181.84 ± 1.89 183.42 ± 4.25 179.81 ± 4.12 |
| MDA (nmol/g FW) | |||||
| CTRL | 39.95 ± 2.03 | 45.28 ± 2.76 | 46.90 ± 1.96 | 43.78 ± 1.11 | 32.04 ± 0.12 |
| FEO | 73.26 ± 1.42 * | 84.03 ± 2.25 * | 85.44 ± 2.28 * | 80.07 ± 2.11 * | 30.13 ± 1.42 |
| PCM myrc. α-pin. | 71.65 ± 1.51 * 53.57 ± 3.43 * 52.28 ± 2.11 * | 81.99 ± 1.19 * 74.31 ± 4.25 * 65.41 ± 2.64 * | 86.09 ± 3.30 * 61.39 ± 2.14 * 63.67 ± 2.38 * | 81.53 ± 0.64 * 65.13 ± 2.27 * 74.09 ± 3.24 * | 31.01 ± 1.45 30.16 ± 1.65 32.49 ± 1.66 |
| % Germination a | Root Length b | Hypocotyl Elongation b | |
|---|---|---|---|
| Control | 83.33b ± 12.02 | 7.72 ± 0.61 | 9.40 ± 0.47 |
| CH | 80.00 ± 11.55 | 7.25 ± 0.92 | 8.17 ± 0.14 |
| CH–EO 25% | 90.00 ± 5.77 | 7.70 ± 0.49 | 8.80 ± 0.35 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ascrizzi, R.; Flamini, G.; Bedini, S.; Tani, C.; Giannotti, P.; Lombardi, T.; Conti, B.; Fraternale, D. Ferulago campestris Essential Oil as Active Ingredient in Chitosan Seed-Coating: Chemical Analyses, Allelopathic Effects, and Protective Activity against the Common Bean Pest Acanthoscelides obtectus. Agronomy 2021, 11, 1578. https://doi.org/10.3390/agronomy11081578
Ascrizzi R, Flamini G, Bedini S, Tani C, Giannotti P, Lombardi T, Conti B, Fraternale D. Ferulago campestris Essential Oil as Active Ingredient in Chitosan Seed-Coating: Chemical Analyses, Allelopathic Effects, and Protective Activity against the Common Bean Pest Acanthoscelides obtectus. Agronomy. 2021; 11(8):1578. https://doi.org/10.3390/agronomy11081578
Chicago/Turabian StyleAscrizzi, Roberta, Guido Flamini, Stefano Bedini, Camilla Tani, Paolo Giannotti, Tiziana Lombardi, Barbara Conti, and Daniele Fraternale. 2021. "Ferulago campestris Essential Oil as Active Ingredient in Chitosan Seed-Coating: Chemical Analyses, Allelopathic Effects, and Protective Activity against the Common Bean Pest Acanthoscelides obtectus" Agronomy 11, no. 8: 1578. https://doi.org/10.3390/agronomy11081578
APA StyleAscrizzi, R., Flamini, G., Bedini, S., Tani, C., Giannotti, P., Lombardi, T., Conti, B., & Fraternale, D. (2021). Ferulago campestris Essential Oil as Active Ingredient in Chitosan Seed-Coating: Chemical Analyses, Allelopathic Effects, and Protective Activity against the Common Bean Pest Acanthoscelides obtectus. Agronomy, 11(8), 1578. https://doi.org/10.3390/agronomy11081578