Abstract
Grazing exclusion has been widely implemented in degraded grassland. However, the changes of plant communities and soil nutrients in response to fencing are still controversial. Thus, the effects of free grazing, 17 and 36 years of fencing on the plant biomass and litter biomass, carbon (C), nitrogen (N) and phosphorus (P) concentrations and stocks of plant, litter and soil were investigated in the temperate steppe grasslands of northern China. The results indicated that fencing increased the aboveground live biomass and litter biomass. In addition, fencing increased C, N and P stocks of aboveground live biomass, litter biomass and soil. Although root biomass and its nutrient stocks were also significantly increased by 17 years of fencing, they were decreased with fencing extending from 17 to 36 years. Moreover, there were no significant differences in aboveground live biomass and soil N and P stocks between 17 and 36 years of fencing. Litter biomass and its C, N and P stocks were positively correlated with soil C, N and P stocks. Our results demonstrated that 17 years of fencing is an effective way to restore vegetation and soil nutrients in the temperate steppe of Inner Mongolia, but a longer fencing duration has no further positive effects on biomass production and soil nutrients accumulation.
1. Introduction
Globally, grassland ecosystems cover approximately 40% of the land surface, which play an important role in preserving primary productivity, species diversity and soil fertility [1,2,3,4]. Artificial disturbances often cause drastic changes in vegetation characteristics and soil properties [5,6,7]. Grazing is considered as the most important grassland utilization way throughout the world, which not only alters plant community composition and productivity, but also affects carbon (C), nitrogen (N) and phosphorus (P) stocks in grassland ecosystems [8,9,10,11,12]. Currently, continuous overgrazing has resulted in widespread grassland degradation. Therefore, some strategies, such as fencing, reseeding and rotational grazing have been used to control grassland degradation for protecting the ecological environment and utilizing grassland sustainably [13,14,15,16,17].
Fencing has become the major grassland restoration practice in recent decades [18,19,20,21]. However, the effects of fencing on grassland ecosystems were inconsistent and even opposite in many previous studies. For example, some studies reported that fencing significantly increased aboveground live biomass and root biomass [22,23,24,25], but other studies reported a decline with fencing [26,27,28]. Additionally, it is often assumed that fencing has positive effects on soil C and N stocks [29,30,31,32], while other studies showed that C and N stocks in fenced areas is equal to or less than those in grazing areas [33,34,35,36,37,38]. A better understanding of the effects of fencing on grassland ecosystems is crucial for adopting a suitable fencing strategy to improve the sustainable use of grassland resources.
Soil nutrient pools play an important role in assessing the effects of fencing on degraded grasslands [39,40,41,42]. In natural grasslands, without artificial fertilization, soil nutrients are mainly affected by vegetation characteristics. To clarify and predict the changes of soil nutrients after grazing exclusion, it is necessary to identify the major vegetable factors that influence soil C, N and P stocks. Some studies indicated that soil C, N and P stocks were closely associated with plant roots in different grassland types, such as alpine meadow, desert grassland and temperate steppe grassland [43,44,45]. De Deyn et al. [46] reported that litter was the major factor in determining soil C stock in different biomes across the globe. However, soil nutrient pools and related influencing factors in grasslands with fencing have not well characterized.
The temperate grasslands in Inner Mongolia are located in the middle of the Eurasian steppe region, which is the main part of the temperate steppe in northern China. It covers about 880.000 km2 and provides goods, services and resources for humans, which also has a profound influence on the global C cycle. Unfortunately, the large area of grasslands suffered severe deterioration due to increasing population pressure, livestock quantity and overgrazing [31,47,48,49]. Although fencing has been extensively employed in our study sites, its effectiveness has not been fully understood. Therefore, we explored the effects of fencing on plant biomass and soil nutrient stocks in these grassland ecosystems. Specifically, our objectives were to investigate: (1) How does the aboveground, root and litter biomass respond to fencing? (2) How does fencing affect the C, N and P concentrations and stocks of plant, litter and soil? (3) Which vegetation factors exert dominant effects on soil nutrient pools in our study?
2. Materials and Methods
2.1. Experimental Sites
The study was performed at the Inner Mongolia Grassland Ecosystem Research Station (116°42′ E 43°38′ N) of the Chinese Academy of Sciences, which is located in Xilin River Basin of Inner Mongolia, China. This area is situated in a semi-arid steppe climate with a mean annual temperature of 2.3 °C, and average annual rainfall of approximately 330 mm. Historical climate data shows that the ratio of annual evaporation to precipitation is 4 to 5. Wind erosion and dust storms occur most frequently during spring, and the average monthly wind speed is 4.9 m/s. Data from the National Meteorological Information Center of China, the annual temperature and precipitation from 1981 to 2017, are presented in Figure 1. The dominant plant species are Leymus chinensis and Stipa grandis. The soil in this study area is chestnut soil with a loamy sand texture, and the sand content is more than 78% [50]. The study sites are comprised of three grassland management sequences: Free grazing, 17 years of fencing (F17) since 2000, and 36 years of fencing (F36) since 1981. Free grazing plots were grazed by sheep all year round, and the stocking rate was approximately 3.0 sheep hm2. Before the fencing was established, F17 and F36 plots had similar grazing management with free grazing plots. The total treatment area is 12 hm2. Soil and vegetation properties were not recorded before fencing in the F36 site. However, background data were investigated before fencing in the F17 site. Both the study sites had the same degradation status at the beginning of the fencing according to the description from the pasture manager. The properties of soil and vegetation before fencing are shown in Table 1.
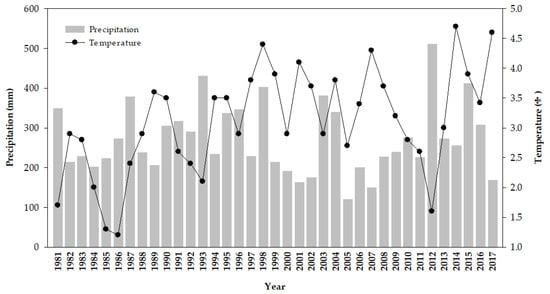
Figure 1.
Annual mean precipitation and temperature from 1981 to 2017 in the study site.

Table 1.
Soil and vegetation properties of the study site before fencing.
2.2. Sampling and Measurements
A typical transect (50 m in length, 10 m in width) was selected at the center in each treatment site on 9 August 2017, and randomly assigned five quadrats of 1 m × 1 m in each transect. All plant species were recorded in each quadrat and divided into different functional groups, including rhizome grass, bunchgrass, legume, forb and carex. Aboveground live biomass of each individual species was harvested by cutting all plants at ground level. The entire litter layer was collected and sorted into two different parts: The L1 layer (fresh undecomposed litter) and the L2 layer (decomposing but still recognizable, fragmented litter). Root biomass was investigated using a root auger (9 cm diameter) to collect soil samples at depths of 0–10 and 10–20 cm. Then soil samples were washed through a 2 mm sieve to get plant roots. Aboveground live biomass, root and litter biomass were oven-dried at 65 °C for 72 h and weighed to determine the dry biomass. Within each quadrat, soil samples were collected at different depths (0–10 and 10–20 cm) from three random points using a bucker auger, and three soil samples in the same layer were mixed into a composite sample. After removing visible roots and debris by hand, the soil samples were air-dried and passed through a 2 mm sieve for soil nutrient analysis. At different soil layers (0–10 and 10–20 cm), we investigated soil bulk density with a 100 cm3 soil bulk sampler for estimating soil C, N and P stocks. The C, N and P concentrations of plant, litter and soil were measured using the dichromate oxidation method [51], the Kjeldahl method [52] and the NaHCO3 alkali digestion method and by molybdenum antimony colorimetry [53], respectively.
2.3. Calculations and Data Analysis
Soil organic carbon stock (SOCS; g m−2), soil total nitrogen stock (STNS; g m−2) and soil total phosphorus stock (STPS; g m−2) were calculated for each soil depth interval by [54]:
where BD is the soil bulk density (g cm−3); CSOC, CSTN and CSTP are the SOC concentration (g kg−1), the STN concentration (g kg−1) and the STP concentration (g kg−1); D is the soil thickness (cm).
SOCS = BD × CSOC × D × 10
STNS = BD × CSTN × D × 10
STPS = BD × CSTP × D × 10
The C, N and P stocks (g m−2) of the plant were calculated by the following equations [55]:
where B is the aboveground live biomass, root biomass and litter biomass (g m−2); C, N and P are the C, N and P concentrations of aboveground live biomass, root biomass and litter biomass (g kg−1).
All data were tested for normality and homogeneity of variances before performing the statistical analysis. Fencing effects on the vegetation, litter and soil indicators were assessed using one-way analysis of variance (ANOVA). A least significant difference (LSD) test was used for multiple comparisons. In addition, stepwise regression analysis was used to examine the relationships of soil C, N and P stocks with the vegetable factors. Figures were created using SigmaPlot for Windows version 10.0 (Systat Software, Inc., Chicago, IL, USA) and all statistical analyses were performed using SPSS 16.0 software (Chicago, IL, USA).
3. Results
3.1. Plant Biomass
Seventeen years of fencing (F17) significantly increased the aboveground live biomass, litter biomass and root biomass compared to free grazing (p < 0.001; Figure 2). The litter biomass in 36 years of fencing (F36) was higher than that in the F17 treatment (p < 0.001). However, aboveground live biomass showed no significant difference between F17 and F36 (p = 0.078). F36 had lower root biomass in 0–10 and 10–20 cm soil layers compared to F17 (p < 0.05). The litter biomass of the L2 layer was 58.5% higher than that of the L1 layer under free grazing. Conversely, the litter biomass of the L1 layer was 184.2% and 163.7% higher than that of the L2 layer under F17 and F36, respectively.
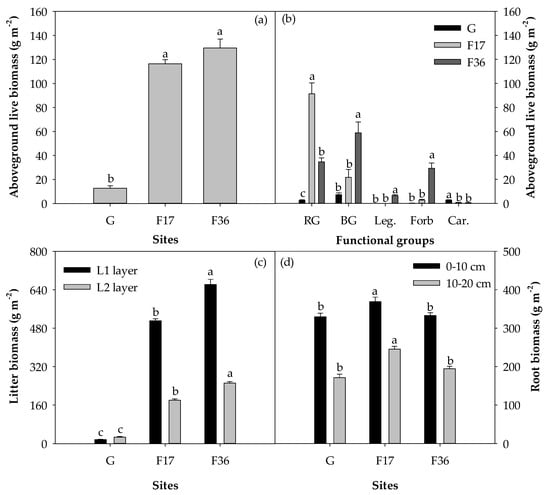
Figure 2.
Variations in (a) aboveground live biomass, (b) aboveground live biomass of rhizome grass (RG), bunchgrass (BG), legume (Leg.), forb (Forb) and carex (Car.), (c) litter biomass (L1 and L2 layers) and (d) root biomass (0–10 and 10–20 cm) among free grazing (G), 17 (F17) and 36 years of fencing (F36) treatments. The values (mean ± SE) are means of five blocks; the same letters above the bars indicate no significant difference at the 0.05 level.
Fencing has varying effects on the aboveground live biomass of different plant functional groups in our study (Figure 2). Aboveground live biomass of rhizome grass was higher, but aboveground live biomass of carex was lower in F17 compared to free grazing (p < 0.001). Aboveground live biomass of bunchgrass, legume and forb significantly increased, but the aboveground live biomass of rhizome grass significantly decreased in F36 compared to F17 (p < 0.01). The aboveground live biomass of bunchgrass in free grazing and F36 treatments accounted for 57% and 45% of the total aboveground live biomass, respectively. The aboveground live biomass of rhizome grass in F17 treatment accounted for 78% of the total aboveground live biomass.
3.2. C, N and P Concentrations of Plant and Soil
Fencing increased the C, N and P concentrations of litter biomass and soil (p < 0.05), as well as the N and P concentrations of aboveground live biomass and root biomass (p < 0.05; Figure 3 and Figure 4). However, there were no significant differences in the C concentrations of aboveground live biomass (p = 0.075) and root biomass (p = 0.426) among different treatments (Figure 3 and Figure 4). Fencing also increased the N and P concentrations of different plant functional groups except for the P concentration of forb (p < 0.05) but had no significant effects on the C concentrations of different plant functional groups (p > 0.05). In different treatments, forb displayed higher N and P concentrations than other plant functional groups except for legume (Figure 3).
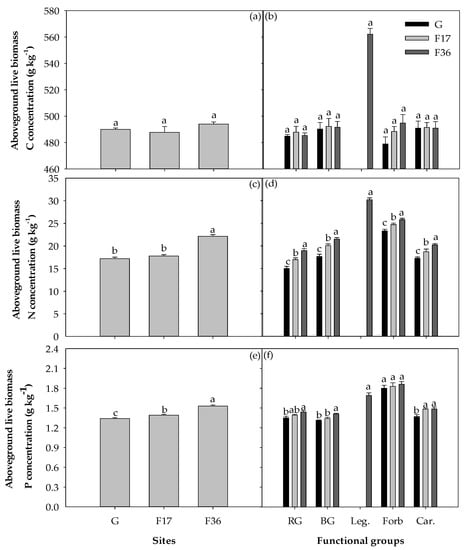
Figure 3.
Variations in (a) aboveground live biomass C concentration, (b) aboveground live biomass C concentration of rhizome grass (RG), bunchgrass (BG), legume (Leg.), forb (Forb) and carex (Car.), (c) aboveground live biomass N concentration, (d) aboveground live biomass N concentration of five plant functional groups, (e) aboveground live biomass P concentration, (f) aboveground live biomass P concentration for five plant functional groups among free grazing (G), 17 (F17) and 36 years of fencing (F36) treatments. The values (mean ± SE) are means of five blocks; the same letters above the bars indicate no significant difference at the 0.05 level.
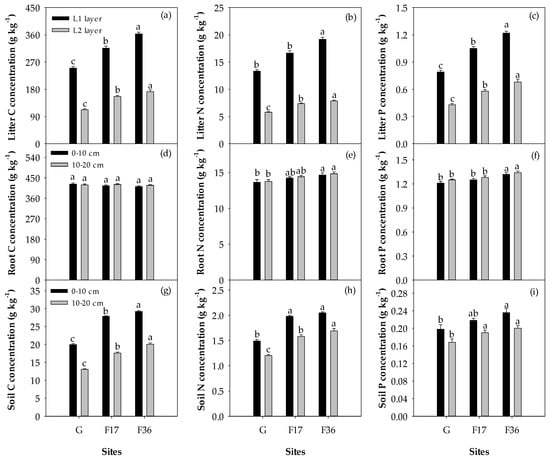
Figure 4.
Variations in (a) litter C concentration (L1 and L2 layers), (b) litter N concentration (L1 and L2 layers), (c) litter P concentration (L1 and L2 layers), (d) root C concentration (0–10 and 10–20 cm), (e) root N concentration (0–10 and 10–20 cm), (f) root P concentration (0–10 and 10–20 cm), (g) soil C concentration (0–10 and 10–20 cm), (h) soil N concentration (0–10 and 10–20 cm), (i) soil P concentration (0–10 and 10–20 cm) among free grazing (G), 17 years of fencing (F17) and 36 years of fencing (F36) treatments. The values (mean ± SE) are means of five blocks; the same letters above the bars indicate no significant difference at the 0.05 level.
3.3. The C, N and P stocks and Ecological Stoichiometry of Plant and Soil
C, N and P stocks of aboveground live biomass and litter biomass were significantly increased by fencing (p < 0.001), especially under F36 (Table 2). F17 increased the C, N and P stocks of root biomass, as well as the soil C and N stocks compared to free grazing (p < 0.001; Table 2). However, F36 had lower C, N and P stocks of root biomass than F17 (p < 0.01). Soil N (p = 0.184) and P stocks (p = 0.397) had no significant differences between F17 and F36. Additionally, soil C stock in the 0–10 cm layer also had no difference between F17 and F36 (p = 0.097).
Table 2.
Ecosystem nutrient stocks and ecological stoichiometry in free grazing, 17 and 36 years of fencing treatments.
The C:N and C:P ratios of aboveground live biomass and root biomass were decreased by fencing (p < 0.05; Table 2). In contrast, fencing increased the C:N and C:P ratios of soil (p < 0.01). Fencing increased the N:P ratios of aboveground live biomass and soil (p < 0.001), but the influences of fencing on the N:P ratios of root biomass were not significant (p = 0.716).
3.4. Relationship between Litter Properties and Soil Nutrient Stocks
Based on the stepwise regression analysis, the litter biomass was positively correlated with soil C, N and P stocks (p < 0.05). In addition, the C, N and P stocks of litter biomass were also significantly positively correlated with soil C, N and P stocks, respectively (p < 0.05; Figure 5).
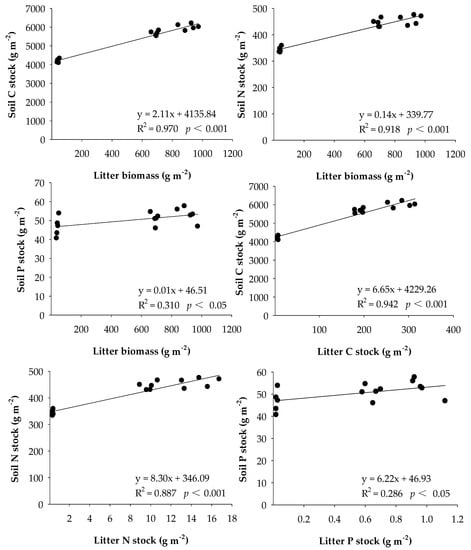
Figure 5.
Relationships between soil nutrient stocks and litter properties.
4. Discussion
Significant increases in aboveground live biomass, root biomass and litter biomass after 17 years of fencing were found in a typical steppe in Inner Mongolia. The results are consistent with previous studies in temperate steppe grasslands [43] and in the alpine meadow [56,57]. However, we found that 36 years of fencing decreased the root biomass and had no effect on the aboveground live biomass compared to 17 years of fencing. The phenomenon could be explained by the fact that the aboveground live biomass of rhizome grass presented a great proportion of total aboveground live biomass after 17 years of fencing, which was significantly decreased by 36 years of fencing. In the present study, fencing prevents the removal of aboveground live biomass by livestock, and results in an increase in litter biomass. Litter accumulation restrained the growth of rhizome grass following the long-term exclusion of disturbances [58]. Furthermore, higher aboveground live biomass of rhizome grass and bunchgrass has been observed in the fenced grasslands. Several reports suggested that fencing tended to improve the growth of gramineous grass because of higher competitive ability in grassland community [59,60].
Our study indicated that fencing had no effects on the C concentrations of aboveground live biomass and root biomass. This might be due to C, as it plays a major role in forming the structural basis of plants [61,62], and had a relatively stable ratio in the dry plant biomass [63]. Meanwhile, we did not detect differences in the C concentrations across different plant functional groups, despite that the significant changes in their aboveground live biomass were found after fencing. A significant increase in the N and P concentrations of aboveground live biomass and root biomass was found after fencing. Contrary results were reported by previous studies, which found that the N and P concentrations of plants were higher in free-grazing grasslands than those in fenced grasslands, because foraging by herbivores resulted in plant regrowth, and young tissue of shoots and leaves is richer in nutrients than aged ones [64,65]. Increased in N and P concentrations of plants by fencing in the present study had two causes. Firstly, plant tissue nutrients were positively related to soil nutrients [66,67], suggesting that higher soil nutrients could be responsible for the significant variation in the N and P concentrations of plants after fencing. Secondly, changes in different plant functional groups indicated that fencing increased the biomass of forbs, and the N and P concentrations of forbs were relatively higher than that in other functional groups. This supports our results that fencing had the potential to enhance the N and P concentrations of plants. We found that fencing significantly improved the C, N and P concentrations of litter biomass and soil, which was consistent with previous results [23,68]. Larger amounts of litter were decomposed in the soil, leading to the increases of soil nutrients through vegetation recovery [69]. Additionally, previous studies reported that feed intake and digestibility by sheep were approximately 1.49 kg d−1 sheep−1 and 60.7% in free grazing grasslands in Inner Mongolia, respectively [70,71]. Therefore, the effects of animal manure input with foraging on soil nutrients also cannot be ignored. In the present study, the C concentrations of litter biomass showed an increasing trend with fencing, which may be attributed to the slower rate of C release than mass loss during the process of decomposition.
Many studies have shown that fencing is an effective approach to improve ecosystem nutrient stocks for degraded grasslands [72,73]. We also found that C, N and P stocks of aboveground live biomass, litter biomass and soil were significantly increased by fencing. Although root nutrient stocks were increased after 17 years of fencing, the opposite results were found when the fencing duration increased from 17 to 36 due to the decline in root biomass along the fencing time gradient. Additionally, soil N and P stocks showed insignificant changes between 17 and 36 years of fencing. In other grassland ecosystems, similar findings were reported where soil nutrient stocks probably reached a steady state with the increase in grazing exclusion duration [30,73]. Results from multiple stepwise regression analysis demonstrated that litter biomass and its nutrient stocks were positively correlated with soil nutrient stocks. This may explain the changes in soil C, N and P stocks after 17 years of fencing. Although litter biomass was significantly increased after 36 years of fencing, the quantity of litter entered the soil was decreased due to the absence of physical breakdown, which was in agreement with a previous study [40]. This may retard the transfer of nutrients from litter to the soil, therefore the differences in soil nutrients stocks were not statistically significant between our two fencing treatments. In addition to the dynamics of litter, soil bulk density also plays a major role in determining soil nutrient stocks. Previous studies have reported that fencing resulted in a reduction in soil bulk density by eliminating the trampling effects of grazing livestock [19,74]. Therefore, continuous fencing is likely to restrain soil nutrients sequestration [75], and increases the risk of wildfires [58]. Compared to free grazing, fencing decreased the C:N and C:P ratios of aboveground live biomass and root biomass, but increased the C:N and C:P ratios of soil. The opposite trend may partly be because the increased plant production led to large amounts of N and P transferred from soil to plants. Additionally, soil C can have a faster rate of accumulation relative to soil N and P due to the faster release rate of C than N and P during litter decomposition [76].
5. Conclusions
Our results indicated that 17 years of fencing had positive effects on vegetation and soil properties. However, 36 years of fencing did not further increase the aboveground live biomass, but reduced the root biomass as a result of the significant change in the biomass of rhizome grass. Soil N and P stocks showed no significant differences between 17 and 36 years of fencing. This study also suggests that litter biomass and its C, N and P stocks were the major factors in determining soil C, N and P stocks. In conclusion, an appropriate fencing strategy (e.g., 17 years of fencing) can be a good management tool for restoring degraded grasslands according to our results, while 36 years of fencing has had no positive effects on biomass production and soil nutrients accumulation in the Inner Mongolia steppe.
Author Contributions
Conceptualization, Q.L. and D.Z.; formal analysis, S.C.; investigation, S.C., Y.H. and Q.L.; writing—original draft preparation, S.C.; writing—review and editing, Q.L.; supervision, Q.L. and D.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Strategic Science and Technology Guide Project of CAS (XDA23060403) and National Key Research and Development Program of China, grant number 2016YFC0500606.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wang, W.; Fang, J.Y. Soil respiration and human effects on global grasslands. Glob. Planet. Chang. 2009, 67, 20–28. [Google Scholar] [CrossRef]
- Ebrahimi, M.; Khosravi, H.; Rigi, M. Short-term grazing exclusion from heavy livestock rangelands affects vegetation cover and soil properties in natural ecosystems of southeastern Iran. Ecol. Eng. 2016, 95, 10–18. [Google Scholar] [CrossRef]
- Scurlock, J.M.O.; Johnson, K.; Olson, R.J. Estimating net primary productivity from grassland biomass dynamics measurements. Glob. Chang. Biol. 2002, 8, 736–753. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.H.; Fang, J.Y.; Ma, W.H.; Guo, D.L.; Mohammat, A. Large-scale pattern of biomass partitioning across China’s grasslands. Glob. Ecol. Biogeogr. 2010, 19, 268–277. [Google Scholar]
- Deng, L.; Sweeney, S.; Shangguan, Z.P. Grassland responses to grazing disturbance: Plant diversity changes with grazing intensity in a desert steppe. Grass Forage Sci. 2014, 69, 524–533. [Google Scholar]
- Su, Y.Z.; Li, Y.L.; Cui, J.Y.; Zhao, W.Z. Influences of continuous grazing and livestock exclusion on soil properties in a degraded sandy grassland, Inner Mongolia, northern China. Catena 2005, 59, 267–278. [Google Scholar]
- Liang, Y.; Han, G.D.; Zhou, H.; Zhao, M.L.; Snyman, H.A.; Shan, D.; Havstad, K.A. Grazing intensity on vegetation dynamics of a typical steppe in northeast Inner Mongolia. Rangel. Ecol. Manag. 2009, 62, 328–336. [Google Scholar] [CrossRef]
- Wu, G.L.; Shang, Z.H.; Zhu, Y.J.; Ding, L.M.; Wang, D. Species-abundance-seed-size patterns within a plant community affected by grazing disturbance. Ecol. Appl. 2015, 25, 848–855. [Google Scholar]
- Zhang, J.; Zuo, X.A.; Zhou, X.; Lv, P.; Lian, J.; Yue, X.Y. Long-term grazing effects on vegetation characteristics and soil properties in a semiarid grassland, northern China. Environ. Monit. Assess. 2017, 189, 216. [Google Scholar] [PubMed]
- López-Mársico, L.; Altesor, A.; Oyarzabal, M.; Baldassini, P.; Paruelo, J.M. Grazing increases below-ground biomass and net primary production in a temperate grassland. Plant Soil 2015, 392, 155–162. [Google Scholar] [CrossRef]
- Derner, J.D.; Boutton, T.W.; Briske, D.D. Grazing and ecosystem carbon storage in the North American great plains. Plant Soil 2006, 280, 77–90. [Google Scholar] [CrossRef]
- Altesor, A.; Oesterheld, M.; Leoni, E.; Lezama, F.; Rodríguez, C. Effect of grazing on community structure and productivity of a Uruguayan grassland. Plant Ecol. 2005, 179, 83–91. [Google Scholar] [CrossRef]
- Akiyama, T.; Kawamura, K. Grassland degradation in China: Methods of monitoring, management and restoration. Grassl. Sci. 2007, 53, 1–17. [Google Scholar] [CrossRef]
- Li, Q.; Zhou, D.W.; Jin, Y.H.; Wang, M.L.; Song, Y.T.; Li, G.D. Effects of fencing on vegetation and soil restoration in a degraded alkaline grassland in northeast China. J. Arid Land 2014, 6, 478–487. [Google Scholar] [CrossRef] [Green Version]
- Barros, A.; Pickering, C.M.; Renison, D. Short-term effects of pack animal grazing exclusion from Andean alpine meadows. Arct. Antarct. Alp. Res. 2014, 46, 333–343. [Google Scholar] [CrossRef] [Green Version]
- Haugland, E.; Froud-Williams, R.J. Improving grasslands: The influence of soil moisture and nitrogen fertilization on the establishment of seedlings. J. Appl. Ecol. 1999, 36, 263–270. [Google Scholar] [CrossRef]
- Bailey, D.W.; Brown, J.R. Rotational grazing systems and livestock grazing behavior in shrub-dominated semi-arid and arid rangelands. Rangel. Ecol. Manag. 2011, 64, 1–9. [Google Scholar] [CrossRef]
- Zhu, G.Y.; Deng, L.; Zhang, X.B.; Shangguan, Z.P. Effects of grazing exclusion on plant community and soil physicochemical properties in a desert steppe on the loess plateau, China. Ecol. Eng. 2016, 90, 372–381. [Google Scholar] [CrossRef]
- Wu, G.L.; Liu, Z.H.; Zhang, L.; Chen, J.M.; Hu, T.M. Long-term fencing improved soil properties and soil organic carbon storage in an alpine swamp meadow of western China. Plant Soil 2010, 332, 331–337. [Google Scholar] [CrossRef]
- He, N.P.; Wu, L.; Wang, Y.S.; Han, X.G. Changes in carbon and nitrogen in soil particle-size fractions along a grassland restoration chronosequence in northern China. Geoderma 2009, 150, 302–308. [Google Scholar] [CrossRef]
- Souter, N.J.; Milne, T. Grazing exclusion as a conservation measure in a South Australian temperate native grassland. Grassl. Sci. 2009, 55, 79–88. [Google Scholar] [CrossRef]
- Cheng, J.M.; Jing, G.H.; Wei, L.; Jing, Z.B. Long-term grazing exclusion effects on vegetation characteristics, soil properties and bacterial communities in the semi-arid grasslands of China. Ecol. Eng. 2016, 97, 170–178. [Google Scholar] [CrossRef]
- Wang, T.W.; Zhang, Z.; Li, Z.B.; Li, P. Grazing management affects plant diversity and soil properties in a temperate steppe in northern China. Catena 2017, 158, 141–147. [Google Scholar] [CrossRef]
- Yan, Y.C.; Tang, H.P.; Chang, R.Y.; Liu, L. Study on the difference of vegetation and soil in typical steppe communities under different fenced time. J. Arid Land Resour. Environ. 2008, 22, 145–151. (In Chinese) [Google Scholar]
- Sun, Z.J.; An, S.Z.; Duan, J.J. Effect of enclosure on vegetation and soil nutrient of sagebrush desert grassland in Xinjiang. Arid Zone Res. 2009, 26, 877–882. (In Chinese) [Google Scholar] [CrossRef]
- Wu, X.W.; Wang, Y.C.; Sun, S.C. Long-term fencing decreases plant diversity and soil organic carbon concentration of the Zoige alpine meadows on the eastern Tibetan plateau. Plant Soil 2019, 458, 191–200. [Google Scholar] [CrossRef]
- Schonbach, P.; Wan, H.W.; Gierus, M.; Bai, Y.F.; Muller, K.; Lin, L.J.; Susenbeth, A.; Taube, F. Grassland responses to grazing: Effects of grazing intensity and management system in an Inner Mongolian steppe ecosystem. Plant Soil 2011, 340, 103–115. [Google Scholar] [CrossRef]
- Frank, D.A.; Kuns, M.M.; Guido, D.R. Consumer control of grassland plant production. Ecology 2002, 83, 602–606. [Google Scholar] [CrossRef]
- Deng, L.; Sweeney, S.; Shangguan, Z.P. Long-term effects of natural enclosure: Carbon stocks, sequestration rates and potential for grassland ecosystems in the Loess Plateau. Clean Soil Air Water 2014, 42, 617–625. [Google Scholar] [CrossRef]
- Zhou, Z.Y.; Li, F.R.; Chen, S.K.; Zhang, H.R.; Li, G.D. Dynamics of vegetation and soil carbon and nitrogen accumulation over 26 years under controlled grazing in a desert shrubland. Plant Soil 2011, 341, 257–268. [Google Scholar] [CrossRef]
- Pei, S.F.; Fu, H.; Wan, C.G. Changes in soil properties and vegetation following exclosure and grazing in degraded Alxa desert steppe of Inner Mongolia, China. Agric. Ecosyst. Environ. 2008, 124, 33–39. [Google Scholar] [CrossRef]
- Steffens, M.; Kolbl, A.; Totsche, K.U.; Kogel-Knabner, I. Grazing effects on soil chemical and physical properties in a semiarid steppe of Inner Mongolia (P.R. China). Geoderma 2008, 143, 63–72. [Google Scholar] [CrossRef]
- Shi, X.M.; Li, X.G.; Li, C.T.; Zhao, Y.; Shang, Z.H.; Ma, Q.F. Grazing exclusion decreases soil organic C storage at an alpine grassland of the Qinghai–Tibetan Plateau. Ecol. Eng. 2013, 57, 183–187. [Google Scholar] [CrossRef]
- Medina-Roldán, E.; Paz-Ferreiro, J.; Bardgett, R.D. Grazing exclusion affects soil and plant communities, but has no impact on soil carbon storage in an upland grassland. Agric. Ecosyst. Environ. 2012, 149, 118–123. [Google Scholar] [CrossRef]
- Reeder, J.D.; Schuman, G.E. Influence of livestock grazing on C sequestration in semi-arid mixed-grass and short-grass rangelands. Environ. Pollut. 2002, 116, 457–463. [Google Scholar] [CrossRef]
- Nosetto, M.D.; Jobbagy, E.G.; Paruelo, J.M. Carbon sequestration in semi-arid rangelands: Comparison of Pinus ponderosa plantations and grazing exclusion in NW Patagonia. J. Arid Environ. 2006, 67, 142–156. [Google Scholar] [CrossRef]
- Shrestha, G.; Stahl, P.D. Carbon accumulation and storage in semi-arid sagebrush steppe: Effects of long-term grazing exclusion. Agric. Ecosyst. Environ. 2008, 125, 173–181. [Google Scholar] [CrossRef]
- Hafner, S.; Unteregelsbacher, S.; Seeber, E.; Lena, B.; Xu, X.L.; Li, X.G.; Guggenberger, G.; Miehe, G.; Kuzyakov, Y. Effect of grazing on carbon stocks and assimilate partitioning in a Tibetan montane pasture revealed by 13CO2 pulse labeling. Glob. Chang. Biol. 2012, 18, 528–538. [Google Scholar] [CrossRef]
- Mekuria, W.; Aynekulu, E. Exclosure land management for restoration of the soils in degraded communal grazing lands in northern Ethiopia. Land Degrad. Dev. 2013, 24, 528–538. [Google Scholar] [CrossRef]
- Lu, X.Y.; Yan, Y.; Sun, J.; Zhang, X.K.; Chen, Y.C.; Wang, X.D.; Cheng, G.W. Carbon, nitrogen, and phosphorus storage in alpine grassland ecosystems of Tibet: Effects of grazing exclusion. Ecol. Evol. 2015, 5, 4492–4504. [Google Scholar] [CrossRef]
- McSherry, M.E.; Ritchie, M.E. Effects of grazing on grassland soil carbon: A global review. Glob. Chang. Biol. 2013, 19, 1347–1357. [Google Scholar] [CrossRef]
- Wang, D.; Wu, G.L.; Zhu, Y.J.; Shi, Z.H. Grazing exclusion effects on above- and below-ground C and N pools of typical grassland on the Loess Plateau (China). Catena 2014, 123, 113–120. [Google Scholar] [CrossRef]
- Bi, X.; Fu, Q.; Fan, Y.; Ma, L.X.; Yang, Z.H.; Nan, B.; Dai, X.H.; Zhang, X.S. Effects of grazing exclusion on the grassland ecosystems of mountain meadows and temperate typical steppe in a mountain-basin system in Central Asia’s arid regions, China. Sci. Total Environ. 2018, 630, 254–263. [Google Scholar] [CrossRef]
- Yang, Z.P.; Ouyang, H.; Zhang, X.Z.; Xu, X.L.; Zhou, C.P.; Yang, W.B. Spatial variability of soil moisture at typical alpine meadow and steppe sites in the Qinghai-Tibetan Plateau permafrost region. Environ. Earth Sci. 2011, 63, 477–488. [Google Scholar] [CrossRef] [Green Version]
- Wen, H.Y.; Niu, D.C.; Fu, H.; Kang, J. Experimental investigation on soil carbon, nitrogen, and their components under grazing and livestock exclusion in steppe and desert steppe grasslands, Northwestern China. Environ. Earth Sci. 2013, 70, 3131–3141. [Google Scholar] [CrossRef]
- De Deyn, G.B.; Cornelissen, J.H.C.; Bardgett, R.D. Plant functional traits and soil carbon sequestration in contrasting biomes. Ecol. Lett. 2008, 11, 516–531. [Google Scholar] [CrossRef]
- Chen, Y.P.; Li, Y.Q.; Zhao, X.Y.; Awada, T.; Shang, W.; Han, J.J. Effects of grazing exclusion on soil properties and on ecosystem carbon and nitrogen storage in a sandy rangeland of Inner Mongolia, Northern China. Environ. Manag. 2012, 50, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Li, Z.S.; Fu, B.J.; Zhou, W.M.; Liu, H.F.; Liu, G.H. Restoration of ecosystem carbon and nitrogen storage and microbial biomass after grazing exclusion in semi-arid grasslands of Inner Mongolia. Ecol. Eng. 2014, 73, 395–403. [Google Scholar] [CrossRef]
- Wiesmeier, M.; Kreyling, O.; Steffens, M.; Schoenbach, P.; Wan, H.W.; Gierus, M.; Taube, F.; Kölbl, A.; Kögel-Knabner, I. Short-term degradation of semiarid grasslands—results from a controlled-grazing experiment in Northern China. J. Plant Nutr. Soil Sci. 2012, 175, 434–442. [Google Scholar] [CrossRef]
- Bai, Y.F.; Wu, J.G.; Clark, C.M.; Naeem, S.; Pan, Q.M.; Huang, J.H.; Zhang, L.X.; Han, X.G. Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: Evidence from Inner Mongolia grasslands. Glob. Chang. Biol. 2010, 16, 358–372. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon and organic matter. In Methods of Soil Analysis, Part 2, 2nd ed.; Page, A.L., Ed.; American Society of Agronomy: Madison, WI, USA, 1982; pp. 539–577. [Google Scholar]
- Bremner, J.M. Nitrogen Total Methods of Soil Analysis. Part 3: Chemical Methods; America Society of Agronomy: Madison, WI, USA, 1996; pp. 1085–1121. [Google Scholar]
- Cao, Y.Z.; Wang, X.D.; Lu, X.Y.; Yan, Y.; Fan, J.H. Soil organic carbon and nutrients along an alpine grassland transect across Northern Tibet. J. Mt. Sci. 2013, 10, 564–573. [Google Scholar] [CrossRef] [Green Version]
- He, N.P.; Yu, Q.; Wu, L.; Wang, Y.S.; Han, X.G. Carbon and nitrogen store and storage potential as affected by land-use in a Leymus chinensis grassland of northern China. Soil Biol. Biochem. 2008, 40, 2952–2959. [Google Scholar] [CrossRef]
- Fang, J.Y.; Guo, Z.D.; Piao, S.L.; Chen, A.P. Terrestrial vegetation carbon sinks in China, 1981–2000. Sci. China Ser. D Earth Sci. 2007, 50, 1341–1350. [Google Scholar] [CrossRef]
- Shang, Z.H.; Deng, B.; Ding, L.M.; Ren, G.H.; Xin, G.S.; Liu, Z.Y.; Wang, Y.L.; Long, R.J. The effects of three years of fencing enclosure on soil seed banks and the relationship with aboveground vegetation of degraded alpine grasslands of the Tibetan plateau. Plant Soil 2013, 364, 229–244. [Google Scholar] [CrossRef]
- Wang, Y.J.; Shen, X.J.; Jiang, M.; Tong, S.Z.; Lu, X.G. Spatiotemporal change of aboveground biomass and its response to climate change in marshes of the Tibetan Plateau. Int. J. Appl. Earth Obs. Geoinf. 2021, 102, 102385. [Google Scholar] [CrossRef]
- Yu, Q.; Wu, H.H.; Wang, Z.W.; Flynn, D.F.B.; Yang, H.; Lv, F.M.; Smith, M.; Han, X.G. Long term prevention of disturbance induces the collapse of a dominant species without altering ecosystem function. Sci. Rep. 2015, 5, 14320. [Google Scholar] [CrossRef] [Green Version]
- Chai, Q.L.; Ma, Z.Y.; Chang, X.F.; Wu, G.L.; Zheng, J.Y.; Li, Z.W.; Wang, G.J. Optimizing management to conserve plant diversity and soil carbon stock of semi-arid grasslands on the Loess Plateau. Catena 2019, 172, 781–788. [Google Scholar] [CrossRef]
- Kull, O.; Aan, A. The relative share of graminoid and forb life-forms in a natural gradient of herb layer productivity. Ecography 1997, 20, 146–154. [Google Scholar] [CrossRef]
- Shen, X.J.; Jiang, M.; Lu, X.G.; Liu, X.T.; Liu, B.H.; Zhang, J.Q.; Wang, X.W.; Tong, S.Z.; Lei, G.C.; Wang, S.Z.; et al. Aboveground biomass and its spatial distribution pattern of herbaceous marsh vegetation in China. Sci. China Earth Sci. 2021, 64, 1115–1125. [Google Scholar] [CrossRef]
- Shen, X.J.; Liu, B.H.; Henderson, M.H.; Wang, L.; Wu, Z.F.; Wu, H.T.; Jiang, M.; Lu, X.G. Asymmetric effects of daytime and nighttime warming on spring phenology in the temperate grasslands of China. Agric. For. Meteorol. 2018, 259, 240–249. [Google Scholar] [CrossRef]
- Schade, J.D.; Kyle, M.; Hobbie, S.E.; Fagan, W.F.; Elser, J.J. Stoichiometric tracking of soil nutrients by a desert insect herbivore. Ecol. Lett. 2003, 6, 96–101. [Google Scholar] [CrossRef] [Green Version]
- Niu, K.C.; He, J.S.; Zhang, S.T.; Lechowicz, M.J. Tradeoffs between forage quality and soil fertility: Lessons from Himalayan rangelands. Agric. Ecosyst. Environ. 2016, 234, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Lambers, H.; Mougel, C.; Jaillard, B.; Hinsinger, P. Plant-microbe-soil interactions in the rhizosphere: An evolutionary perspective. Plant Soil 2009, 321, 83–115. [Google Scholar] [CrossRef]
- Zeng, Q.C.; Li, X.; Dong, Y.H.; An, S.S.; Darboux, F. Soil and plant components ecological stoichiometry in four steppe communities in the Loess Plateau of China. Catena 2016, 147, 481–488. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, B.R.; An, S.S. Ecological stoichiometry in leaves, roots, litters and soil among different plant communities in a desertified region of Northern China. Catena 2018, 166, 328–338. [Google Scholar] [CrossRef]
- Wang, K.B.; Deng, L.; Ren, Z.P.; Li, J.P.; Shangguan, Z.P. Grazing exclusion significantly improves grassland ecosystem C and N pools in a desert steppe of northwest China. Catena 2016, 137, 441–448. [Google Scholar] [CrossRef]
- Jing, Z.B.; Cheng, J.M.; Su, J.S.; Bai, Y.; Jin, J.W. Changes in plant community composition and soil properties under 3-decade grazing exclusion in semiarid grassland. Ecol. Eng. 2014, 64, 171–178. [Google Scholar] [CrossRef]
- Wei, Z.J.; Bai, Y.J.; Wu, R.T.; Yang, J.; Yang, S.M. Studies on grazing behavior under different grazing systems on desert steppe. Acta Agrestia Sin. 2005, 13, 57–61. (In Chinese) [Google Scholar]
- Wu, R.N. Study on Grazing Behavior, Herbage Intake and Digestibility of Grazing Sheep in the Inner Mongolian Typical Steppe. Master’s Thesis, Inner Mongolia Agriculture University, Hohhot, China, May 2010. [Google Scholar]
- Qiu, L.P.; Wei, X.R.; Zhang, X.C.; Cheng, J.M. Ecosystem carbon and nitrogen accumulation after grazing exclusion in semiarid grassland. PLoS ONE 2013, 8, e55433. [Google Scholar]
- Liu, J.; Zhang, Q.C.; Li, Y.; Di, H.J.; Xu, J.M.; Li, J.Y.; Guan, X.M.; Xu, X.Y.; Pan, H. Effects of pasture management on soil fertility and microbial communities in the semi-arid grasslands of Inner Mongolia. J. Soils Sediments 2016, 16, 235–242. [Google Scholar] [CrossRef]
- Hu, Z.M.; Li, S.G.; Guo, Q.; Niu, S.L.; He, N.P.; Li, L.H.; Yu, G.R. A synthesis of the effect of grazing exclusion on carbon dynamics in grasslands in China. Glob. Chang. Biol. 2016, 22, 1385–1393. [Google Scholar] [CrossRef]
- Shao, C.; Chen, J.; Li, L.; Zhang, L. Ecosystem responses to mowing manipulations in an arid Inner Mongolia steppe: An energy perspective. J. Arid Environ. 2012, 82, 1–10. [Google Scholar] [CrossRef]
- Yin, X.R.; Liang, C.Z.; Wang, L.X.; Wang, W.; Liu, Z.L.; Liu, X.P. Ecological stoichiometry of plant nutrients at different restoration succession stages in typical steppe of Inner Mongolia, China. Chin. J. Plant Ecol. 2010, 34, 39–47. (In Chinese) [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).