
Differential Morphophysiological, Biochemical, and Molecular Responses of Maize Hybrids to Salinity and Alkalinity Stresses


 ,
,  ,
, 
 , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Data Recorded
2.2.1. Morphological Growth
2.2.2. Photosynthetic Pigments
2.2.3. Inorganic Ion
2.2.4. Lipid Peroxidation and H2O2 Content
2.2.5. Antioxidant Enzymes Activities
2.2.6. RNA Isolation, cDNA Synthesis, and Transcriptional Analysis
2.3. Statistical Analysis
3. Results
3.1. Seedling Fresh and Dry Weights
3.2. Seedling Length and Number of Leaves
3.3. Chlorophyll Content and Carotenoid Content
3.4. Ions Accumulation
3.5. Hydrogen Peroxide and Lipid Peroxidation
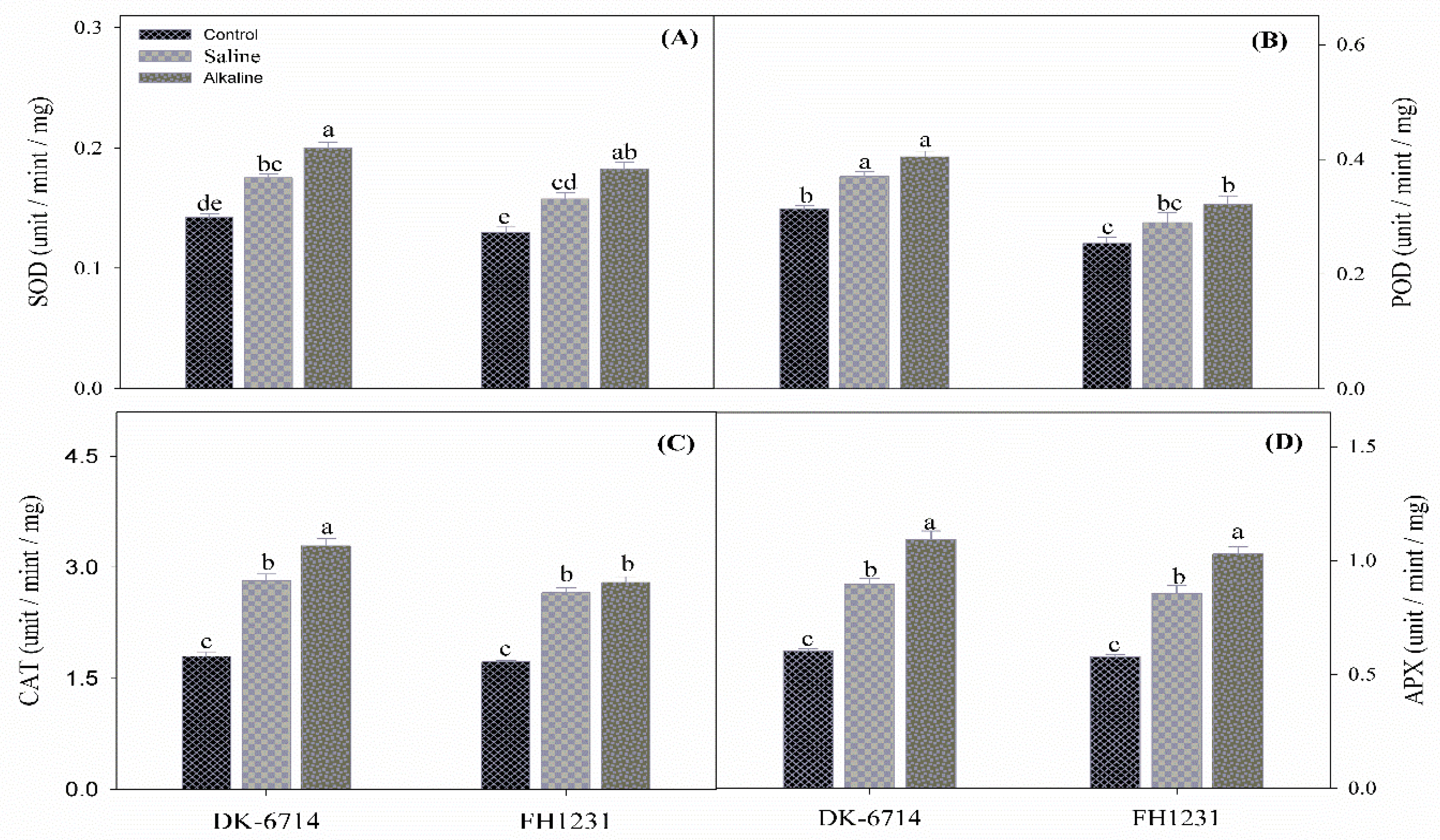
3.6. Antioxidant Enzymatic Activity
3.7. Gene Expression Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Honarjoo, N.; Mojiri, A.; Jalalian, A.; Karimzadeh, H.R. The effects of salinity and alkalinity of soil on growth of Haloxylon sp. In Proceedings of the 2010 International Conference on Chemistry and Chemical Engineering, Segzi, Iran, 1–3 Augest 2010; pp. 285–288. [Google Scholar]
- Vysotskaya, L.; Hedley, P.E.; Sharipova, G.; Veselov, D.; Kudoyarova, G.; Morris, J.; Jones, H.G. Effect of salinity on water relations of wild barley plants differing in salt tolerance. AoB Plants 2010. [Google Scholar] [CrossRef]
- Dass, S.; Singh, I.; Chikkappa, G.; Parihar, C.; Kaul, J.; Singode, A.; Manivannan, A.; Singh, D. Abiotic stresses in Maize: Some issues and solutions. Dir. Maize Res. Indian Counc. Agric. Res. Pusa Campus New Delhi 2010, 110012. Available online: https://www.researchgate.net/profile/Jyoti_Kaul/publication/290908407_Abiotic_Stresses_in_Maize_Some_Issues_and_Solutions/links/569c8a8208aea14769547d65.pdf (accessed on 27 April 2021).
- Pakistan, M.O.F.G.O. Pakistan Economic Survey 2016–2017. 2017. Available online: http://www.finance.gov.pk/survey/chapters_17/02-agriculture.pdf (accessed on 27 April 2021).
- Gong, F.; Yang, L.; Tai, F.; Hu, X.; Wang, W. “Omics” of maize stress response for sustainable food production: Opportunities and challenges. Omics A J. Integr. Biol. 2014, 18, 714–732. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, I.; de Vos, R.C.; Bones, A.M.; Hall, R.D. Plant molecular stress responses face climate change. Trends Plant Sci. 2010, 15, 664–674. [Google Scholar] [CrossRef]
- Carpici, E.B.; Celik, N.; Bayram, G. The effects of salt stress on the growth, biochemical parameter and mineral element content of some maize (Zea mays L.) cultivars. Afr. J. Biotechnol. 2010, 9, 6937–6942. [Google Scholar]
- Hu, X.; Zhang, Y.; Shi, Y.; Zhang, Z.; Zou, Z.; Zhang, H.; Zhao, J. Effect of exogenous spermidine on polyamine content and metabolism in tomato exposed to salinity–alkalinity mixed stress. Plant Physiol. Biochem. 2012, 57, 200–209. [Google Scholar] [CrossRef]
- Shi, D.; Wang, D. Effects of various salt-alkaline mixed stresses on Aneurolepidium chinense (Trin.) Kitag. Plant Soil 2005, 271, 15–26. [Google Scholar] [CrossRef]
- Rao, P.S.; Mishra, B.; Gupta, S.; Rathore, A. Reproductive stage tolerance to salinity and alkalinity stresses in rice genotypes. Plant Breed. 2008, 127, 256–261. [Google Scholar] [CrossRef]
- Yan, X.; Tan, K. Screening rice varieties for SALT tolerance in the greenhouse. IRRN 1991, 16, 16–17. [Google Scholar]
- Alam, S.; Ansari, R.; Khan, M. Saline Agriculture and Pakistan. Industry and Economy. Pak. Load. Bus. Mag. Last 2000, 25. Available online: http://www.pakistaneconomist.com (accessed on 27 April 2021).
- Niaz, A.; Ranjha, A.; Rahmatullah, A.H.; Waqas, M. Boron status of soils as affected by different soil characteristics–pH, CaCO3, organic matter and clay contents. Pak. J. Agric. Sci. 2007, 44, 428–435. [Google Scholar]
- Azooz, M.; Shaddad, M.; Abdel-Latef, A. Leaf growth and K+/Na+ ratio as an indication of the salt tolerance of three sorghum cultivars grown under salinity stress and IAA treatment. Acta Agron. Hung. 2004, 52, 287–296. [Google Scholar] [CrossRef]
- Sultana, N.; Ikeda, T.; Kashem, M. Effect of seawater on photosynthesis and dry matter accumulation in developing rice grains. Photosynthetica 2002, 40, 115–119. [Google Scholar] [CrossRef]
- Rengasamy, P. Soil processes affecting crop production in salt-affected soils. Funct. Plant Biol. 2010, 37, 613–620. [Google Scholar] [CrossRef]
- Saslis-Lagoudakis, C.H.; Hua, X.; Bui, E.; Moray, C.; Bromham, L. Predicting species’ tolerance to salinity and alkalinity using distribution data and geochemical modelling: A case study using Australian grasses. Ann. Bot. 2015, 115, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Shi, D. Effects of NaCl and Na2CO3 on growth of Puccinellia tenuiflora and on present state of mineral elements in nutrient solution. Acta Pratacu. Sin. 1997, 6, 51–61. [Google Scholar]
- Yang, C.; Wang, P.; Li, C.; Shi, D.; Wang, D. Comparison of effects of salt and alkali stresses on the growth and photosynthesis of wheat. Photosynthetica 2008, 46, 107–114. [Google Scholar] [CrossRef]
- Zhanwu, G.; Jiayu, H.; Chunsheng, M.; Jixiang, L.; Xiaoyu, L.; Lidong, L.; Shengnan, S. Effects of saline and alkaline stresses on growth and physiological changes in oat (Avena sativa L.) seedlings. Not. Bot. Horti Agrobot. Cluj Napoca 2014, 42, 357–362. [Google Scholar]
- Guo, R.; Shi, L.; Yan, C.; Zhong, X.; Gu, F.; Liu, Q.; Xia, X.; Li, H. Ionomic and metabolic responses to neutral salt or alkaline salt stresses in maize (Zea mays L.) seedlings. BMC Plant Biol. 2017, 17, 41. [Google Scholar] [CrossRef]
- Pardo, J.M.; Quintero, F.J. Plants and sodium ions: Keeping company with the enemy. Genome Biol. 2002, 3, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Zhang, S.; Xu, X.; Korpelainen, H.; Li, C. Effect of increased alkalinity on Na+ and K+ contents, lipid peroxidation and antioxidative enzymes in two populations of Populus cathayana. Biol. Plant 2009, 53, 597. [Google Scholar] [CrossRef]
- Bazzaz, M.M.; Hossain, M.A. Plant water relations and proline accumulations in soybean under salt and water stress environment. J. Plant Sci. 2015, 3, 272–278. [Google Scholar]
- Cramer, G.R.; Nowak, R.S. Supplemental manganese improves the relative growth, net assimilation and photosynthetic rates of salt-stressed barley. Physiol. Plant 1992, 84, 600–605. [Google Scholar] [CrossRef]
- Sali, A.; Rusinovci, I.; Fetahu, S.; Gashi, B.; Simeonovska, E.; Rozman, L. The effect of salt stress on the germination of maize (Zea mays L.) seeds and photosynthetic pigments. Acta Agric. Slov. 2015, 105, 85–94. [Google Scholar]
- Nishiuchi, S.; Fujihara, K.; Liu, S.; Takano, T. Analysis of expressed sequence tags from a NaHCO3-treated alkali-tolerant plant, Chloris virgata. Plant Physiol. Biochem. 2010, 48, 247–255. [Google Scholar] [CrossRef]
- Zhang, J.-T.; Mu, C.-S. Effects of saline and alkaline stresses on the germination, growth, photosynthesis, ionic balance and anti-oxidant system in an alkali-tolerant leguminous forage Lathyrus quinquenervius. Soil Sci. Plant Nutr. 2009, 55, 685–697. [Google Scholar] [CrossRef]
- Trchounian, A.; Petrosyan, M.; Sahakyan, N. Plant cell redox homeostasis and reactive oxygen species. In Redox State as a Central Regulator of Plant-Cell Stress Responses; Springer: Cham, Switzerland, 2016; pp. 25–50. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Neill, S.; Desikan, R.; Hancock, J. Hydrogen peroxide signalling. Curr. Opin. Plant Biol. 2002, 5, 388–395. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef]
- Van Breusegem, F.; Vranová, E.; Dat, J.F.; Inzé, D. The role of active oxygen species in plant signal transduction. Plant Sci. 2001, 161, 405–414. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Xu, Y.; Kim, S.O.; Li, Y.; Han, J. Autophagy contributes to caspase-independent macrophage cell death. J. Biol. Chem. 2006, 281, 19179–19187. [Google Scholar] [CrossRef]
- Williams, V.; Twine, S. Flame photometric method for sodium, potassium and calcium. Mod. Methods Plant Anal. 1960, 5, 3–5. [Google Scholar]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A. Assay of catalases and peroxidases. Methods Enzym. 1955, 2, 764–775. [Google Scholar]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Vwioko, E.; Adinkwu, O.; El-Esawi, M.A. Comparative Physiological, Biochemical and Genetic Responses to Prolonged Waterlogging Stress in Okra and Maize Given Exogenous Ethylene Priming. Front. Physiol. 2017, 8, 632. [Google Scholar] [CrossRef]
- Neta, I.C.S.; Von Pinho, É.V.D.R.; de Abreu, V.M.; dos Santos, H.O.; Vilela, D.R.; Von Pinho, R.G.; de Carvalho, M.L.M. Enzyme activities and gene expression in dry maize seeds and seeds submitted to low germination temperature. Afr. J. Agric. Res. 2016, 11, 3097–3103. [Google Scholar]
- Huang, H.; Lv, L.; Wang, D.; Guo, B.; Lv, J.; Luo, L.; Wen, B.; Kang, Y. Biochemical and molecular responses of maize (Zea mays L.) to 1, 2-dibromo-4- (1, 2 dibromoethyl) cyclohexane (TBECH) diastereomers: Oxidative stress, DNA damage, antioxidant enzyme gene expression and diversity of root exudates. Sci. Total Environ. 2021, 753, 141872. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2 (-Delta Delta C (T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Lv, B.S.; Li, X.W.; Ma, H.Y.; Sun, Y.; Wei, L.X.; Jiang, C.J.; Liang, Z.W. Differences in Growth and Physiology of Rice in Response to Different Saline-Alkaline Stress Factors. Agron. J. 2013, 105, 1119–1128. [Google Scholar] [CrossRef]
- Guo, M.; Wang, R.; Wang, J.; Hua, K.; Wang, Y.; Liu, X.; Yao, S. ALT1, a Snf2 family chromatin remodeling ATPase, negatively regulates alkaline tolerance through enhanced defense against oxidative stress in rice. PLoS ONE 2014, 9, e112515. [Google Scholar] [CrossRef]
- Wei, L.-X.; Lv, B.-S.; Wang, M.-M.; Ma, H.-Y.; Yang, H.-Y.; Liu, X.-L.; Jiang, C.-J.; Liang, Z.-W. Priming effect of abscisic acid on alkaline stress tolerance in rice (Oryza sativa L.) seedlings. Plant Physiol. Biochem. 2015, 90, 50–57. [Google Scholar] [CrossRef] [PubMed]
- AbdElgawad, H.; Zinta, G.; Hegab, M.M.; Pandey, R.; Asard, H.; Abuelsoud, W. High salinity induces different oxidative stress and antioxidant responses in maize seedlings organs. Front. Plant Sci. 2016, 7, 276. [Google Scholar] [CrossRef]
- Wang, L.; Seki, K.; Miyazaki, T.; Ishihama, Y. The causes of soil alkalinization in the Songnen Plain of Northeast China. Paddy Water Environ. 2009, 7, 259–270. [Google Scholar] [CrossRef]
- Amini, S.; Ghadiri, H.; Chen, C.; Marschner, P. Salt-affected soils, reclamation, carbon dynamics, and biochar: A review. J. Soils Sediments 2016, 16, 939–953. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology, 4th ed.; Sinauer Associates, Inc., Publishers: Sunderland, MA, USA, 2006; p. 764. [Google Scholar]
- Roychoudhury, A.; Basu, S. Overexpression of an abiotic-stress inducible plant protein in the bacteria Escherichia coli. Afr. J. Biotechnol. 2008, 7, 3231–3234. Available online: https://www.ajol.info/index.php/ajb/article/view/59263 (accessed on 27 April 2021).
- Rao, G.; Rao, G. Pigment composition and chlorophyllase activity in pigeon pea (Cajanus indicus Spreng) and Gingelley (Sesamum indicum L.) under NaCl salinity (1981). Indian J. Exp. Biol. 2013, 19, 768–770. [Google Scholar]
- Noreen, Z.; Ashraf, M. Assessment of variation in antioxidative defense system in salt-treated pea (Pisum sativum) cultivars and its putative use as salinity tolerance markers. J. Plant Physiol. 2009, 166, 1764–1774. [Google Scholar] [CrossRef]
- Kariola, T.; Brader, G.; Li, J.; Palva, E.T. Chlorophyllase 1, a damage control enzyme, affects the balance between defense pathways in plants. Plant Cell 2005, 17, 282–294. [Google Scholar] [CrossRef]
- Latef, A.A.H.A.; Chaoxing, H. Effect of arbuscular mycorrhizal fungi on growth, mineral nutrition, antioxidant enzymes activity and fruit yield of tomato grown under salinity stress. Sci. Hortic. 2011, 127, 228–233. [Google Scholar] [CrossRef]
- Abdel Latef, A.; Abu Alhmad, M.; Sallam, M. Comparative study of the physiological response of two wheat cultivars to salinity stress. Assiut Univ. J. Bot. 2014, 43, 55–69. [Google Scholar]
- James, R.A.; Davenport, R.J.; Munns, R. Physiological characterization of two genes for Na+ exclusion in durum wheat, Nax1 and Nax2. Plant Physiol. 2006, 142, 1537–1547. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Xing, J.; Lan, H. Comparative effects of neutral salt and alkaline salt stress on seed germination, early seedling growth and physiological response of a halophyte species Chenopodium glaucum. Afr. J. Biotechnol. 2012, 11, 9572–9581. [Google Scholar]
- Zhu, J.-K. Regulation of ion homeostasis under salt stress. Curr. Opin. Plant Biol. 2003, 6, 441–445. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Shi, D.; Sheng, Y. Effect of various salt–alkaline mixed stress conditions on sunflower seedlings and analysis of their stress factors. Environ. Exp. Bot. 2005, 54, 8–21. [Google Scholar] [CrossRef]
- Mandhania, S.; Madan, S.; Sawhney, V. Antioxidant defense mechanism under salt stress in wheat seedlings. Biol. Plant 2006, 50, 227–231. [Google Scholar] [CrossRef]
- Ahmad, P.; Kumar, A.; Ashraf, M.; Akram, N.A. Salt-induced changes in photosynthetic activity and oxidative defense system of three cultivars of mustard (Brassica juncea L.). Afr. J. Biotechnol. 2012, 11, 2694–2703. [Google Scholar]
- Ashraf, M.; Athar, H.; Harris, P.; Kwon, T. Some prospective strategies for improving crop salt tolerance. Adv. Agron. 2008, 97, 45–110. [Google Scholar]
- Caverzan, A.; Casassola, A.; Brammer, S.P. Antioxidant responses of wheat plants under stress. Genet. Mol. Biol. 2016, 39, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Scandalios, J.G. Oxidative Stress and the Molecular Biology of Antioxidant Defenses; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1997. [Google Scholar]
- Scandalios, J. Oxidative stress: Molecular perception and transduction of signals triggering antioxidant gene defenses. Braz. J. Med. Biol. Res. 2005, 38, 995–1014. [Google Scholar] [CrossRef]
- Menezes-Benavente, L.; Teixeira, F.K.; Kamei, C.L.A.; Margis-Pinheiro, M. Salt stress induces altered expression of genes encoding antioxidant enzymes in seedlings of a Brazilian indica rice (Oryza sativa L.). Plant Sci. 2004, 166, 323–331. [Google Scholar] [CrossRef]
- Heidari, M.; Jamshidi, P. Effects of salinity and potassium application on antioxidant enzyme activities and physiological parameters in pearl millet. Agric. Sci. China 2011, 10, 228–237. [Google Scholar] [CrossRef]
- Bor, M.; Özdemir, F.; Türkan, I. The effect of salt stress on lipid peroxidation and antioxidants in leaves of sugar beet Beta vulgaris L. and wild beet Beta maritima L. Plant Sci. 2003, 164, 77–84. [Google Scholar] [CrossRef]
- Shigeoka, S.; Nakano, Y.; Kitaoka, S. Metabolism of hydrogen peroxide in Euglena gracilis Z by L-ascorbic acid peroxidase. Biochem. J. 1980, 186, 377–380. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, S.R.; Obado, S.O.; Mauricio, I.L.; Kelly, J.M. Trypanosoma cruzi expresses a plant-like ascorbate-dependent hemoperoxidase localized to the endoplasmic reticulum. Proc. Natl. Acad. Sci. USA 2002, 99, 13453–13458. [Google Scholar] [CrossRef]
- Sano, S.; Ueda, M.; Kitajima, S.; Takeda, T.; Shigeoka, S.; Kurano, N.; Miyachi, S.; Miyake, C.; Yokota, A. Characterization of ascorbate peroxidases from unicellular red alga Galdieria partita. Plant Cell Physiol. 2001, 42, 433–440. [Google Scholar] [CrossRef]
- Takeda, T.; Yoshimura, K.; Ishikawa, T.; Shigeoka, S. Purification and characterization of ascorbate peroxidase in Chlorella vulgaris. Biochimie 1998, 80, 295–301. [Google Scholar] [CrossRef]
- Takeda, T.; Yoshimura, K.; Yoshii, M.; Kanahoshi, H.; Miyasaka, H.; Shigeoka, S. Molecular characterization and physiological role of ascorbate peroxidase from halotolerant Chlamydomonas sp. W80 strain. Arch. Biochem. Biophys. 2000, 376, 82–90. [Google Scholar] [CrossRef]
- Caverzan, A.; Passaia, G.; Rosa, S.B.; Ribeiro, C.W.; Lazzarotto, F.; Margis-Pinheiro, M. Plant responses to stresses: Role of ascorbate peroxidase in the antioxidant protection. Genet. Mol. Biol. 2012, 35, 1011–1019. [Google Scholar] [CrossRef]
- Caverzan, A.; Bonifacio, A.; Carvalho, F.E.; Andrade, C.M.; Passaia, G.; Schünemann, M.; dos Santos Maraschin, F.; Martins, M.O.; Teixeira, F.K.; Rauber, R. The knockdown of chloroplastic ascorbate peroxidases reveals its regulatory role in the photosynthesis and protection under photo-oxidative stress in rice. Plant Sci. 2014, 214, 74–87. [Google Scholar] [CrossRef] [PubMed]
- El-Esawi, M.A.; Alaraidh, I.A.; Alsahli, A.A.; Alzahrani, S.M.; Ali, H.M.; Alayafi, A.A.; Ahmad, M. Serratia liquefaciens KM4 Improves Salt Stress Tolerance in Maize by Regulating Redox Potential, Ion Homeostasis, Leaf Gas Exchange and Stress-Related Gene Expression. Int. J. Mol. Sci. 2018, 19, 3310. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.; Pu, S. Tissue-and genotype-specific ascorbate peroxidase expression in sweet potato in response to salt stress. Biol. Plant 2010, 54, 664–670. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Gene ID (Accession Number) | Primer Sequences (5′–3′) | References |
|---|---|---|---|
| SOD (superoxide dismutase 4) | NM_001112234 | F: TGGAGCACCAGAAGATGA R: CTCGTGTCC ACCCTTTCC | Vwioko et al. [42]; Neta et al. [43] |
| APX (ascorbate peroxidase 2) | NM_001112030 | F: TGAGCGACCAGGACATTG R: GAGGGCTTTGTCACTTGGT | Vwioko et al. [42]; Neta et al. [43] |
| CAT (catalase isomer 3) | NM_001152957 | F: AATCGCCTCACGAATCGGTCCT R: CGCACCACAACAACCACTACGA | Huang et al. [44] |
| POD (peroxidase 39 isoform X1) | NM_001112570 | F: TCGGCGTCATCGTCAGTGCT R: GGCGAGGTTGAGGCTCTTGTTC | Huang et al. [44] |
| Actin2 | EU958048 | F: TGAAACCTTCGAATGCCCAG R: GATTGGAACCGTGTGGCTCA | Huang et al. [44] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fatima, A.; Hussain, S.; Hussain, S.; Ali, B.; Ashraf, U.; Zulfiqar, U.; Aslam, Z.; Al-Robai, S.A.; Alzahrani, F.O.; Hano, C.; et al. Differential Morphophysiological, Biochemical, and Molecular Responses of Maize Hybrids to Salinity and Alkalinity Stresses. Agronomy 2021, 11, 1150. https://doi.org/10.3390/agronomy11061150
Fatima A, Hussain S, Hussain S, Ali B, Ashraf U, Zulfiqar U, Aslam Z, Al-Robai SA, Alzahrani FO, Hano C, et al. Differential Morphophysiological, Biochemical, and Molecular Responses of Maize Hybrids to Salinity and Alkalinity Stresses. Agronomy. 2021; 11(6):1150. https://doi.org/10.3390/agronomy11061150
Chicago/Turabian StyleFatima, Arooj, Saddam Hussain, Sadam Hussain, Basharat Ali, Umair Ashraf, Usman Zulfiqar, Zubair Aslam, Sami Asir Al-Robai, Fatima Omari Alzahrani, Christophe Hano, and et al. 2021. "Differential Morphophysiological, Biochemical, and Molecular Responses of Maize Hybrids to Salinity and Alkalinity Stresses" Agronomy 11, no. 6: 1150. https://doi.org/10.3390/agronomy11061150
APA StyleFatima, A., Hussain, S., Hussain, S., Ali, B., Ashraf, U., Zulfiqar, U., Aslam, Z., Al-Robai, S. A., Alzahrani, F. O., Hano, C., & El-Esawi, M. A. (2021). Differential Morphophysiological, Biochemical, and Molecular Responses of Maize Hybrids to Salinity and Alkalinity Stresses. Agronomy, 11(6), 1150. https://doi.org/10.3390/agronomy11061150