
Intercultivar Diversity of Sour Orange (Citrus aurantium L.) Based on Genetic Markers, Phenotypic Characteristics, Aromatic Compounds and Sensorial Analysis

 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Biological Materials
2.2. Phenotypic Description
2.2.1. Data Acquisition
2.2.2. Statistical Analysis
2.3. Essential Oil Analysis
2.3.1. Raw Materials
2.3.2. Hydro Distillation
2.3.3. Essential Oil Analysis Using Gas Chromatography and Gas Chomatrography-Mass Spectrometry
2.3.4. Statistical Analysis
2.4. Sensorial Analysis of Sour Orange Peel Oil
2.4.1. Tests, Panelists and Descriptors
2.4.2. Statistical Analysis
2.5. Genetic Diversity Analysis
2.5.1. DNA Extraction
2.5.2. SSR Genotyping
2.5.3. SNP Genotyping
3. Results
3.1. Genetic Diversity
3.2. Phenotypic Description of the Fruit
3.3. Essential Oil Analysis
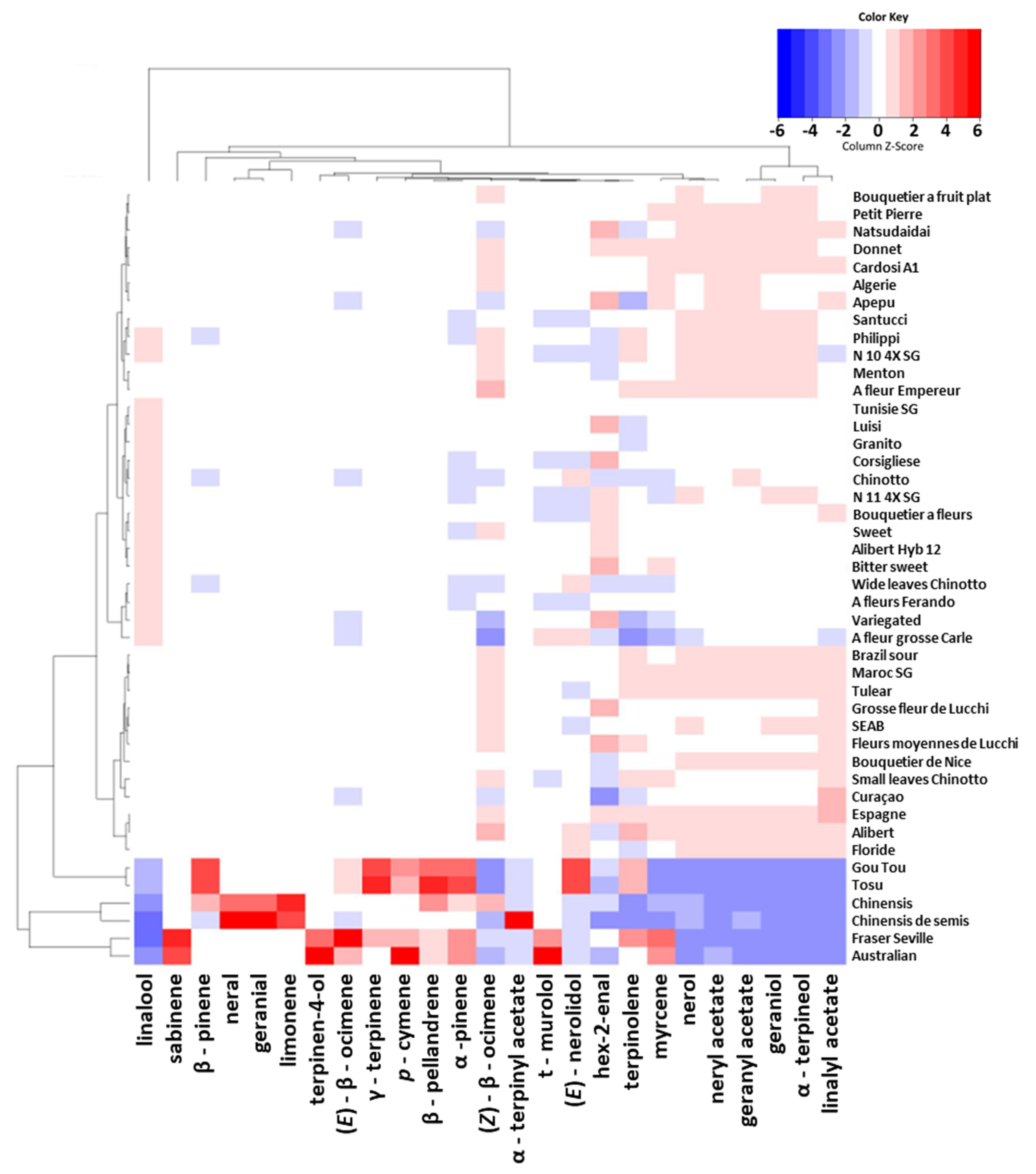
3.3.1. Peel Essential Oil
3.3.2. Peel Essential Oil Sensorial Analysis
3.3.3. Leaf Essential Oil
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Wu, G.A.; Prochnik, S.; Jenkins, J.; Salse, J.; Hellsten, U.; Murat, F.; Perrier, X.; Ruiz, M.; Scalabrin, S.; Terol, J.; et al. Sequencing of diverse mandarin, pummelo and orange genomes reveals complex history of admixture during citrus domestication. Nat. Biotechnol. 2014, 32, 656–662. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.A.; Terol, J.; Ibanez, V.; López-García, A.; Pérez-Román, E.; Borredá, C.; Domingo, C.; Tadeo, F.; Carbonell-Caballero, J.; Alonso, R.; et al. Genomics of the origin and evolution of Citrus. Nat. Cell Biol. 2018, 554, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Yang, X.; Yamamoto, M.; Biswas, M.K. The Genus Citrus; Talon, M., Caruso, M., Gmitter, J.F.G., Eds.; Elsevier WP: Duxford, UK, 2020; pp. 33–56. ISBN 978-0-12-812217-4. [Google Scholar]
- Castle, W.S. A Career Perspective on Citrus Rootstocks, Their Development, and Commercialization. Hort. Sci. 2010, 45, 11–15. [Google Scholar] [CrossRef]
- Moreno, P.; Ambrós, S.; Albiach-Martí, M.R.; Guerri, J.; Peña, L. Citrus tristeza virus: A pathogen that changed the course of the citrus industry. Mol. Plant Pathol. 2008, 9, 251–268. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Muñoz, N.; Velázquez, K.; Vives, M.C.; Ruiz-Ruiz, S.; Pina, J.A.; Flores, R.; Moreno, P.; Guerri, J. The resistance of sour orange to Citrus tristeza virus is mediated by both the salicylic acid and RNA silencing defence pathways. Mol. Plant Pathol. 2016, 18, 1253–1266. [Google Scholar] [CrossRef]
- Anwar, S.; Ahmed, N.; Speciale, A.; Cimino, F.; Saija, A. Bitter Orange (Citrus aurantium L.) Oils. In Essential Oils in Food Preservation, Flavor and Safety, 1st ed.; Preedy, V.R., Ed.; Academic Press: Cambridge, MA, USA; Elsevier BV: Amsterdam, The Netherlands, 2016; pp. 259–268. [Google Scholar]
- Lota, M.-L.; Serra, D.D.R.; Jacquemond, C.; Casanova, J. Chemical variability of peel and leaf essential oils of sour orange. Flavour Fragr. J. 2001, 16, 89–96. [Google Scholar] [CrossRef]
- Peyron, L. Citrus: The Genus Citrus; Dugo, G., Di Giacomo, A., Eds.; Taylor & Francis, CRC Press: London, UK, 2002; Volume 26, pp. 148–152. ISBN 978-0-415-28491-2. [Google Scholar]
- Dugo, G.; Bonaccorsi, I.L.; Sciarrone, D.; Costa, R.; Dugo, P.; Mondello, L.; Santi, L.; Fakhry, H.A. Characterization of Oils from the Fruits, Leaves and Flowers of the Bitter Orange Tree. J. Essent. Oil Res. 2011, 23, 45–59. [Google Scholar] [CrossRef]
- Swingle, W.T.; Reece, P.C.; Reuther, W.; Webber, H.J.; Batchelor, L.D. The Citrus Industry; University of Calofornia Press: Berkeley, CA, USA, 1967; pp. 190–430. [Google Scholar]
- Tanaka, T. Fundamental Discussion of Citrus Classification; Studia Citrologica: Osaka, Japan, 1977; Volume 14, pp. 1–6. [Google Scholar]
- Mabberley, D. A classification for edible Citrus (Rutaceae). Telopea 1997, 167–172. [Google Scholar] [CrossRef]
- Wooldridge, T.J.S. Cross-Compatibility, Graft-Compatibility, and Phylogenetic Relationships in the Auranti-oideae: New Data from the Balsamocitrinae. Ph.D. Thesis, University of Calfornia, Riverside, CA, USA, 2016. [Google Scholar]
- Bernet, G.P.; Asíns, M.J. Identification and genomic distribution of gypsy like retrotransposons in Citrus and Poncirus. Theor. Appl. Genet. 2003, 108, 121–130. [Google Scholar] [CrossRef]
- Ollitrault, P.; Curk, F.; Krueger, R. Citrus taxonomy. In The Genus Citrus; Talon, M., Caruso, M., Gmitter, J.F.G., Eds.; Elsevier WP: Duxford, UK, 2020; pp. 57–81. ISBN 978-0-12-812217-4. [Google Scholar]
- Gogorcena, Y.; Ortiz, J.M. Characterisation of sour orange (Citrus aurantium) cultivars. J. Sci. Food Agric. 1989, 48, 275–284. [Google Scholar] [CrossRef]
- Gogorcena, Y.; Ortiz, J. Use of multivariate analysis in the taxonomy of Citrus aurantium L. and relatives. Sci. Hortic. 1993, 53, 301–310. [Google Scholar] [CrossRef]
- Luro, F.; Gatto, J.; Costantino, G.; Pailly, O. Analysis of genetic diversity in Citrus. Plant Genet. Resour. 2011, 9, 218–221. [Google Scholar] [CrossRef]
- Luro, F.; Rist, D.; Ollitrault, P. Evaluation of genetic relationships in citrus genus by means of sequence tagged microsatellites. Acta Hortic. 2001, 237–242. [Google Scholar] [CrossRef]
- Barkley, N.A.; Roose, M.L.; Krueger, R.R.; Federici, C.T. Assessing genetic diversity and population structure in a citrus germplasm collection utilizing simple sequence repeat markers (SSRs). Theor. Appl. Genet. 2006, 112, 1519–1531. [Google Scholar] [CrossRef]
- Polat, I.; Kaçar, Y.; Yesiloglu, T.; Uzun, A.; Tuzcu, O.; Gulsen, O.; Incesu, M.; Kafa, G.; Turgutoglu, E.; Anil, S. Molecular characterization of sour orange (Citrus aurantium) accessions and their relatives using SSR and SRAP markers. Genet. Mol. Res. 2012, 11, 3267–3276. [Google Scholar] [CrossRef]
- Luro, F.; Viglietti, G.; Marchi, E.; Costantino, G.; Scarpa, G.M.; Tomi, F.; Paoli, M.; Curk, F.; Ollitrault, P. Genetic, morphological and chemical investigations reveal the genetic origin of Pompia (C. medica tuberosa Risso & Poiteau)—An old endemic Sardinian citrus fruit. Phytochemistry 2019, 168, 112083. [Google Scholar] [CrossRef]
- Siragusa, M.; De Pasquale, F.; Abbate, L.; Tusa, N. Identification of Sour Orange Accessions and Evaluation of Their Genetic Variability by Molecular Marker Analyses. HortScience 2006, 41, 84–89. [Google Scholar] [CrossRef]
- Bernet, G.P.; Gorris, M.T.; Carbonell, E.A.; Cambra, M.; Asins, M.J. Citrus tristeza virusresistance in a core collection of sour orange based on a diversity study of three germplasm collections using QTL-linked markers. Plant Breed. 2008, 127, 398–406. [Google Scholar] [CrossRef]
- Shimizu, T.; Kitajima, A.; Nonaka, K.; Yoshioka, T.; Ohta, S.; Goto, S.; Toyoda, A.; Fujiyama, A.; Mochizuki, T.; Nagasaki, H.; et al. Hybrid Origins of Citrus Varieties Inferred from DNA Marker Analysis of Nuclear and Organelle Genomes. PLoS ONE 2016, 11, e0166969. [Google Scholar] [CrossRef]
- Maekawa, K.; Kodama, M.; Kushii, M.; Mitamura, M. Essential Oils of Some Orange Peels. Agric. Biol. Chem. 1967, 31, 373–377. [Google Scholar] [CrossRef]
- Shaw, P.E. Review of quantitative analyses of citrus essential oils. J. Agric. Food Chem. 1979, 27, 246–257. [Google Scholar] [CrossRef]
- Dugo, G.; Cotroneo, A.; Bonaccorsi, I.; Trozzi, A. Citrus Oils: Composition, Advanced Analytical Techniques, Contaminants, and Biological Activity, 1st ed.; Dugo, G., Modello, L., Eds.; CRC Press: Boca Raton, FL, USA, 2010; pp. 10–20. ISBN 978-1-4398-0029-4. [Google Scholar]
- González-Mas, M.C.; Rambla, J.L.; López-Gresa, M.P.; Blázquez, M.A.; Granell, A. Volatile Compounds in Citrus Essential Oils: A Comprehensive Review. Front. Plant Sci. 2019, 10, 12. [Google Scholar] [CrossRef]
- Gaff, M.; Esteban-Decloux, M.; Giampaoli, P. Bitter orange peel essential oil: A review of the different factors and chemical reactions influencing its composition. Flavour Fragr. J. 2020, 35, 247–269. [Google Scholar] [CrossRef]
- Boussaada, O.; Skoula, M.; Kokkalou, E.; Chemli, R. Chemical Variability of Flowers, Leaves, and Peels Oils of Four Sour Orange Provenances. J. Essent. Oil Bear. Plants 2007, 10, 453–464. [Google Scholar] [CrossRef]
- Boussaada, O.; Chemli, R. Seasonal Variation of Essential Oil Composition of Citrus Aurantium, L. var. amara. J. Essent. Oil Bear. Plants 2007, 10, 109–120. [Google Scholar] [CrossRef]
- Ellouze, I.; Abderrabba, M.; Sabaou, N.; Mathieu, F.; Lebrihi, A.; Bouajila, J. Season’s Variation Impact onCitrus aurantiumLeaves Essential Oil: Chemical Composition and Biological Activities. J. Food Sci. 2012, 77, 173–180. [Google Scholar] [CrossRef]
- Song, H.S.; Sawamura, M.; Ito, T.; Ido, A.; Ukeda, H. Quantitative determination and characteristic flavour of daidai (Citrus aurantium L. var.cyathifera Y. Tanaka) peel oil. Flavour Fragr. J. 2000, 15, 323–328. [Google Scholar] [CrossRef]
- Deterre, S.; Rega, B.; Delarue, J.; Decloux, M.; Lebrun, M.; Giampaoli, P. Identification of key aroma compounds from bitter orange (Citrus aurantium L.) products: Essential oil and macerate-distillate extract. Flavour Fragr. J. 2011, 27, 77–88. [Google Scholar] [CrossRef]
- Luro, F.; Bloquel, E.; Tomu, B.; Costantino, G.; Tur, I.; Riolacci, S.; Varamo, F.; Ollitrault, P.; Froelicher, Y.; Curk, F.; et al. The INRA-CIRAD Citrus Germplasm Collection of San Giuliano, Corsica; Publications du Centre Jean Bérard: Naples, Italy, 2017; pp. 243–261. [Google Scholar] [CrossRef]
- Jimenez-Cuesta, M.; Cuquerella, J.; Martinez-Javaga, J.M. Determination of a color index for citrus fruit de-greening. In Proceedings of the International Society of Citriculture/International Citrus Congres, Tokyo, Japan, 9–12 November 1982. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis, 2nd ed.; Springer International Publishing: New York, NY, USA, 2016; pp. 3–253. ISBN 978-3-319-24275-0. [Google Scholar]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research, 2nd ed.; John Wiley & Sons: New York, NY, USA, 1984; pp. 3–700. ISBN 978-0-471-87092-0. [Google Scholar]
- Thioulouse, J.; Dray, S.; Dufour, A.; Siberchicot, A.; Jombart, T.; Pavoine, S. Multivariate Analysis of Ecological Data with ade4, 1st ed.; Springer: New York, NY, USA, 2018; pp. 3–329. ISBN 978-1-4939-8850-1. [Google Scholar]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: AnRPackage for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. R Package Version 1.0.7. 2020. Available online: https://CRAN.R-project.org/package=factoextra (accessed on 25 January 2021).
- Warnes, G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Huber, W.; Liaw, A.; Lumley, T.; Maechler, M.; Magnusson, A.; Moeller, S.; et al. Gplots: Various R Programming Tools for Plotting Data. Available online: https://CRAN.R-project.org/package=gplots (accessed on 25 January 2021).
- Hervé, M. RVAideMemoire: Testing and Plotting Procedures for Biostatistics. R Package Version 0.9-79. 2020. Available online: https://CRAN.R-project.org/package=RVAideMemoire (accessed on 25 January 2021).
- Luro, F.L.; Costantino, G.; Terol, J.; Argout, X.; Allario, T.; Wincker, P.; Talon, M.; Ollitrault, P.; Morillon, R. Transferability of the EST-SSRs developed on Nules clementine (Citrus clementina Hort ex Tan) to other Citrus species and their effectiveness for genetic mapping. BMC Genom. 2008, 9, 287. [Google Scholar] [CrossRef]
- Ligges, U.; Mächler, M. Scatterplot3d—An R Package for Visualizing Multivariate Data. J. Stat. Softw. 2003, 8, 1–20. [Google Scholar] [CrossRef]
- Curk, F.; Ancillo, G.; Ollitrault, F.; Perrier, X.; Jacquemoud-Collet, J.-P.; Garcia-Lor, A.; Navarro, L.; Ollitrault, P. Nuclear Species-Diagnostic SNP Markers Mined from 454 Amplicon Sequencing Reveal Admixture Genomic Structure of Modern Citrus Varieties. PLoS ONE 2015, 10, e0125628. [Google Scholar] [CrossRef]
- Lombardo, G.; Schicchi, R.; Marino, P.; Palla, F. Genetic analysis ofCitrus aurantiumL. (Rutaceae) cultivars by ISSR molecular markers. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2011, 146, 19–26. [Google Scholar] [CrossRef]
- Bretó, M.; Ruiz, C.; Pina, J.; Asíns, M. The Diversification of Citrus clementina Hort. ex Tan., a Vegetatively Propagated Crop Species. Mol. Phylogenet. Evol. 2001, 21, 285–293. [Google Scholar] [CrossRef]
- Garcia-Lor, A.; Luro, F.; Ollitrault, P.; Navarro, L. Genetic diversity and population structure analysis of mandarin germplasm by nuclear, chloroplastic and mitochondrial markers. Tree Genet. Genomes 2015, 11, 1–15. [Google Scholar] [CrossRef]
- Barrett, H.C.; Hutchison, D.J. Spontaneous Tetraploidy in Apomietie Seedlings of Citrus. Econ. Bot. 1978, 32, 27–45. [Google Scholar] [CrossRef]
- Cameron, J.W.; Scora, R.W. A Comparison of rind oil components of diploid and tetraploid citrus by gas-liquid-chromatography. Taxon 1968, 17, 128–135. [Google Scholar] [CrossRef]
- Rouseff, R.L.; Perez-Cacho, P.R.; Jabalpurwala, F. Historical Review of Citrus Flavor Research during the Past 100 Years. J. Agric. Food Chem. 2009, 57, 8115–8124. [Google Scholar] [CrossRef] [PubMed]
- Kohzaki, K.; Gomi, K.; Yamasaki-Kokudo, Y.; Ozawa, R.; Takabayashi, J.; Akimitsu, K. Characterization of a sabinene synthase gene from rough lemon (Citrus jambhiri). J. Plant Physiol. 2009, 166, 1700–1704. [Google Scholar] [CrossRef] [PubMed]
- Alquézar, B.; Rodríguez, A.; De La Peña, M.; Peña, L. Genomic Analysis of Terpene Synthase Family and Functional Characterization of Seven Sesquiterpene Synthases from Citrus sinensis. Front. Plant Sci. 2017, 8, 1481. [Google Scholar] [CrossRef]
- Lota, M.-L.; de Rocca Serra, D.; Tomi, F.; Bessiere, J.-M.; Casanova, J. Chemical Composition of Peel and Leaf Essential Oils of Citrus Medica, L. and C. Limonimedica Lush. Flavour Fragr. J. 1999, 14, 161–166. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrer, V.; Costantino, G.; Paoli, M.; Paymal, N.; Quinton, C.; Ollitrault, P.; Tomi, F.; Luro, F. Intercultivar Diversity of Sour Orange (Citrus aurantium L.) Based on Genetic Markers, Phenotypic Characteristics, Aromatic Compounds and Sensorial Analysis. Agronomy 2021, 11, 1084. https://doi.org/10.3390/agronomy11061084
Ferrer V, Costantino G, Paoli M, Paymal N, Quinton C, Ollitrault P, Tomi F, Luro F. Intercultivar Diversity of Sour Orange (Citrus aurantium L.) Based on Genetic Markers, Phenotypic Characteristics, Aromatic Compounds and Sensorial Analysis. Agronomy. 2021; 11(6):1084. https://doi.org/10.3390/agronomy11061084
Chicago/Turabian StyleFerrer, Vincent, Gilles Costantino, Mathieu Paoli, Noémie Paymal, Carole Quinton, Patrick Ollitrault, Félix Tomi, and François Luro. 2021. "Intercultivar Diversity of Sour Orange (Citrus aurantium L.) Based on Genetic Markers, Phenotypic Characteristics, Aromatic Compounds and Sensorial Analysis" Agronomy 11, no. 6: 1084. https://doi.org/10.3390/agronomy11061084
APA StyleFerrer, V., Costantino, G., Paoli, M., Paymal, N., Quinton, C., Ollitrault, P., Tomi, F., & Luro, F. (2021). Intercultivar Diversity of Sour Orange (Citrus aurantium L.) Based on Genetic Markers, Phenotypic Characteristics, Aromatic Compounds and Sensorial Analysis. Agronomy, 11(6), 1084. https://doi.org/10.3390/agronomy11061084