
Hyperspectral Detection and Monitoring of Salt Stress in Pomegranate Cultivars


 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Collection of Leaf Spectra
2.3. Standard Measurements
2.4. Analyses of Spectral Signatures
2.5. PLSR-Model Calibration and Validation
2.6. Estimation of Leaf Traits by PLSR-Models and Vegetation Spectral Indices
2.7. Statistical Analysis of Leaf Traits Estimated from Spectra, Vegetation Spectral Indices, and Ψw
3. Results
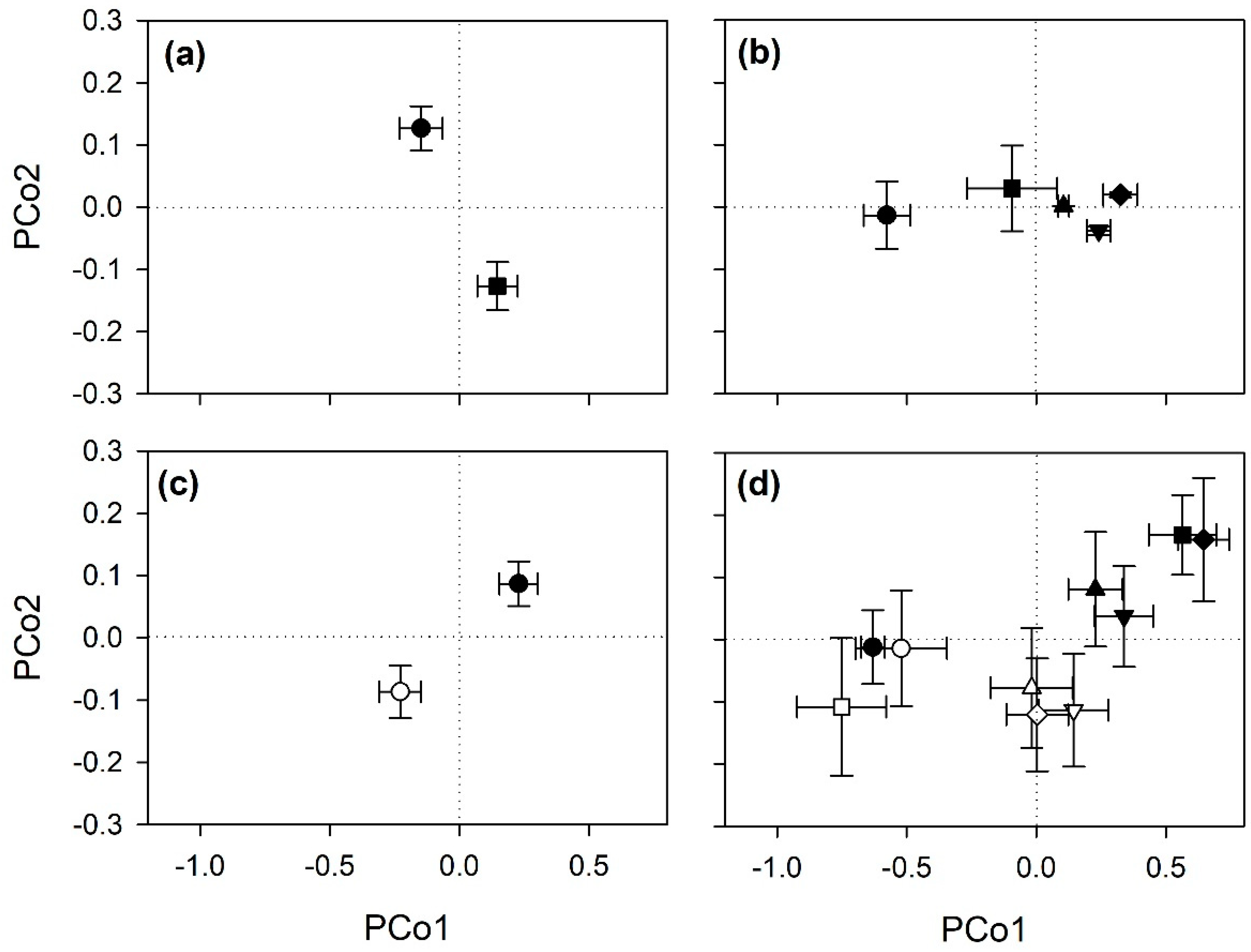
3.1. Analyses of Spectral Singatures
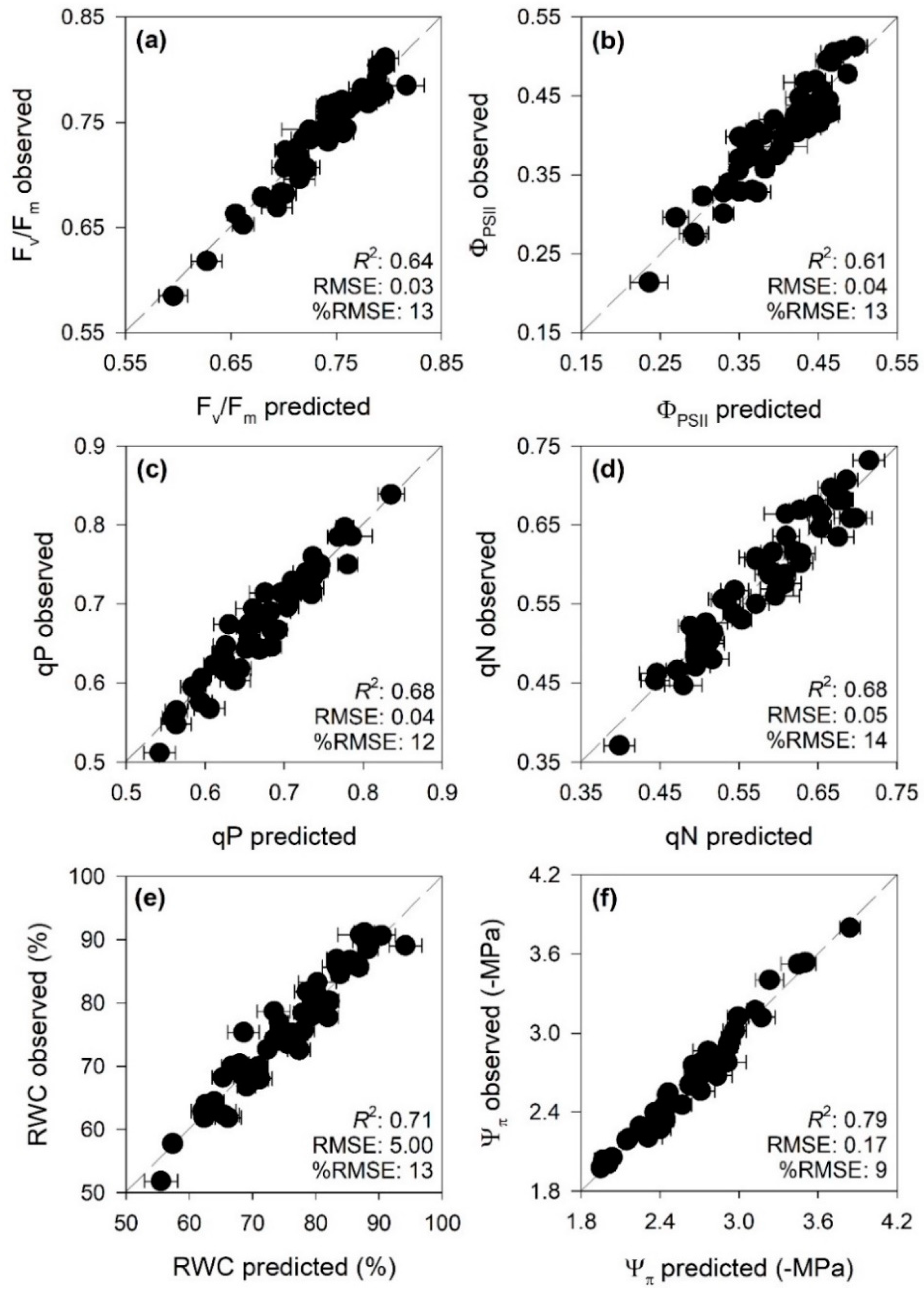
3.2. PLSR Prediction Models
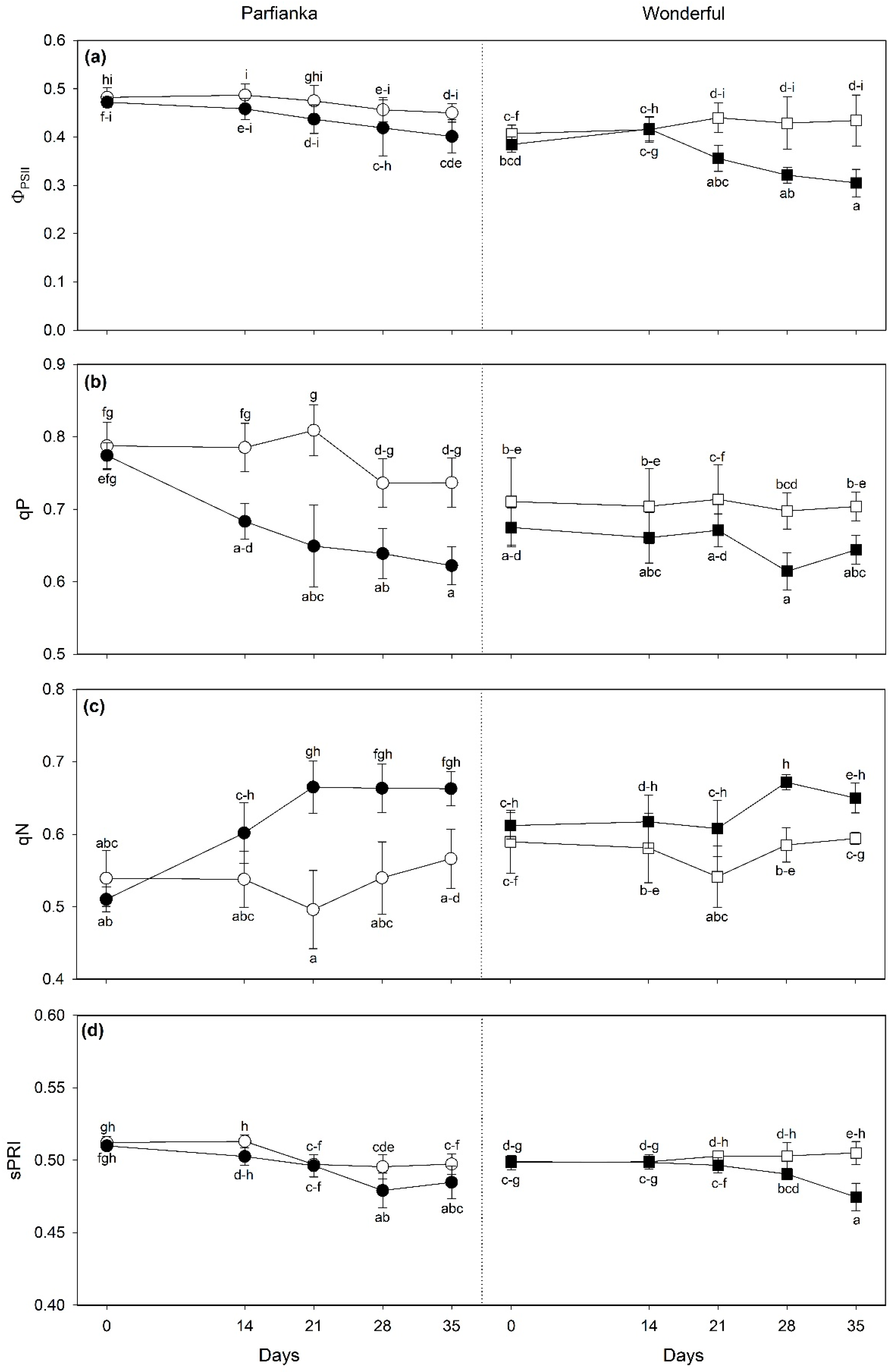
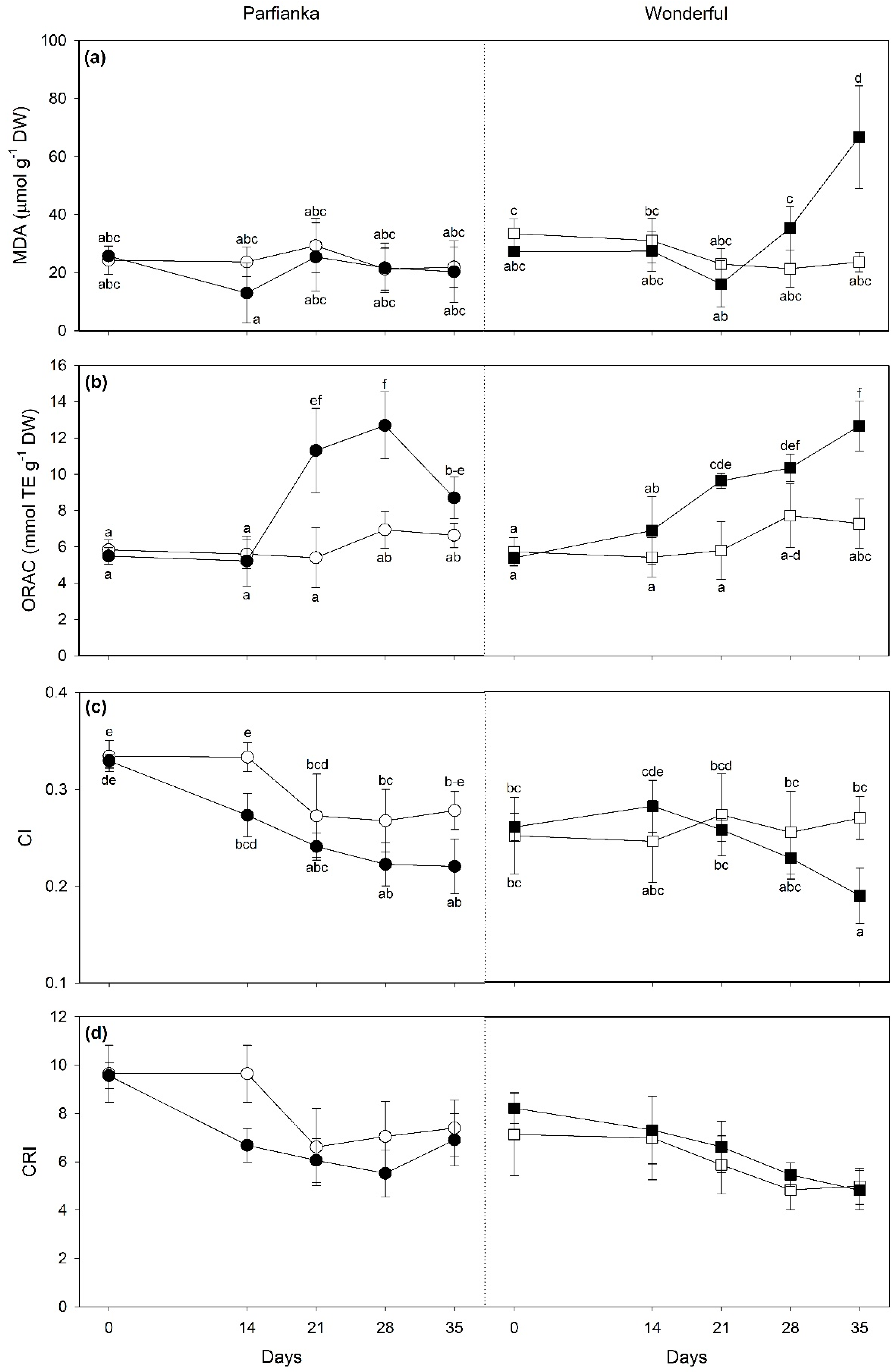
3.3. Variations of Spectra-Estimated Parameters and Vegetation Spectral Indices
4. Discussion
4.1. Hyperspectral Discrimination of Cultivars and Salinity Conditions
4.2. Spectroscopic Estimation of Photosynthetic Performance and Water Status Parameters
4.3. Variations of Spectra-Estimated Parameters and Vegetation Spectral Indices
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Isayenkov, S.V.; Maathuis, F.J.M. Plant salinity stress: Many unanswered questions remain. Front. Plant Sci. 2019, 10, 80. [Google Scholar] [CrossRef]
- Olesen, J.E.; Trnka, M.; Kersebaum, K.C.; Skjelvåg, A.O.; Seguin, B.; Peltonen-Sainio, P.; Rossi, F.; Kozyra, J.; Micale, F. Impacts and adaptation of European crop production systems to climate change. Eur. J. Agron. 2011, 34, 96–112. [Google Scholar] [CrossRef]
- Olsson, L.; Barbosa, S.; Bhadwal, S.; Cowie, A.; Delusca, D.; Flores-Renteira, D.; Hermans, K.; Jobbagy, E.; Kurz, W.; Li, D.; et al. Land Degradation. In Climate Change and Land: An IPCC Special Report on Climate Change, Desertification, Land degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems; Shukla, P.R., Skea, J., Calvo Buendia, E., Masson-Delmotte, V., Pörtner, H.-H., Roberts, D.C., Zhai, P., Slade, R., Connors, S., van Diemen, R., et al., Eds.; IPCC: Geneva, Switzerland, 2019; pp. 345–436. [Google Scholar]
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi. J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef]
- Cotrozzi, L.; Couture, J.J. Hyperspectral assessment of plant responses to multi-stress environments: Prospects for managing protected agrosystems. Plants People Planet 2020, 2, 244–258. [Google Scholar] [CrossRef]
- Cotrozzi, L.; Townsend, P.A.; Pellegrini, E.; Nali, C.; Couture, J.J. Reflectance spectroscopy: A novel approach to better understand and monitor the impact of air pollution on Mediterranean plants. Environ. Sci. Pollut. Res. 2018, 25, 8249–8267. [Google Scholar] [CrossRef] [PubMed]
- Gamon, J.A.; Field, C.B.; Goulden, M.L.; Griffin, K.L.; Hartley, A.E.; Joel, G.; Peñuelas, J.; Valentini, R. Relationships between NDVI, canopy structure, and photosynthesis in three Californian vegetation types. Ecol. Appl. 1995, 5, 28–41. [Google Scholar] [CrossRef]
- Gamon, J.; Serrano, L.; Surfus, J. The photochemical reflectance index: An optical indicator of photosynthetic radiation use efficiency across species, functional types, and nutrient levels. Oecologia 1997, 112, 492–501. [Google Scholar] [CrossRef]
- Serbin, S.P.; Singh, A.; Desai, A.R.; Dubois, S.G.; Jablosnki, A.D.; Kingdon, C.C.; Kruger, E.L.; Townsend, P.A. Remotely estimating photosynthetic capacity, and its response to temperature, in vegetation canopies using imaging spectroscopy. Remote Sens. Environ. 2015, 167, 78–87. [Google Scholar] [CrossRef]
- Couture, J.J.; Singh, A.; Rubert-Nason, K.F.; Serbin, S.P.; Lindroth, R.L.; Townsend, P.A. Spectroscopic determination of ecologically relevant plant secondary metabolites. Methods Ecol. Evol. 2016, 7, 1402–1412. [Google Scholar] [CrossRef]
- Cotrozzi, L.; Couture, J.J.; Cavender-Bares, J.; Kingdon, C.C.; Fallon, B.; Pilz, G.; Pellegrini, E.; Nali, C.; Townsend, P.A. Using foliar spectral properties to assess the effects of drought on plant water potential. Tree Physiol. 2017, 37, 1582–1591. [Google Scholar] [CrossRef]
- Ely, K.S.; Burnett, A.C.; Lieberman-Cribbin, W.; Serbin, S.P.; Rogers, A. Spectroscopy can predict key leaf traits associated with source-sink balance and carbon-nitrogen status. J. Exp. Bot. 2019, 70, 1789–1799. [Google Scholar] [CrossRef] [PubMed]
- Marchica, A.; Loré, S.; Cotrozzi, L.; Lorenzini, G.; Nali, C.; Pellegrini, E.; Remorini, D. Early detection of sage (Salvia officinalis L.) responses to ozone using reflectance spectroscopy. Plants 2019, 8, 346. [Google Scholar] [CrossRef] [PubMed]
- Wold, S.; Sjӧstrӧm, M.; Eriksson, L. PLS-regression: A basic tool of chemometrics. Chemom. Intell. Lab. Syst. 2001, 58, 109–130. [Google Scholar] [CrossRef]
- Mahlein, A.-K.; Kuska, M.T.; Behmann, J.; Polder, G.; Walter, A. Hyperspectral sensors and imaging technologies in phytopathology: State of the art. Annu. Rev. Phytopathol. 2018, 56, 535–558. [Google Scholar] [CrossRef]
- Weber, V.S.; Araus, J.L.; Cairns, J.E.; Sanchez, C.; Melchinger, A.E.; Orsini, E. Prediction of grain yield using reflectance spectra of canopy and leaves in maize plants grown under different water regimes. Field Crops Res. 2012, 128, 82–90. [Google Scholar] [CrossRef]
- Araus, J.L.; Cairns, J.E. Field high-throughput phenotyping: The new crop breeding frontier. Trends Plant Sci. 2014, 19, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Couture, J.J.; Singh, A.; Charkowski, A.O.; Groves, R.L.; Gray, S.M.; Bethke, P.C.; Townsend, P.A. Integrating spectroscopy with potato disease management. Plant Dis. 2018, 102, 2233–2240. [Google Scholar] [CrossRef]
- Moghimi, A.; Yang, C.; Miller, M.E.; Kianian, S.F.; Marchetto, P.M. A novel approach to assess salt stress tolerance in wheat using hyperspectral imaging. Front. Plant Sci. 2018, 9, 1182. [Google Scholar] [CrossRef] [PubMed]
- El-Hendawy, S.; Al-Suhaibani, N.; Alotaibi, M. Estimating growth and photosynthetic properties of wheat grown in simulated saline field conditions using hyperspectral reflectance sensing and multivariate analysis. Sci. Rep. 2019, 9, 16473. [Google Scholar] [CrossRef]
- Zhu, K.; Sun, Z.; Zhao, F.; Yang, T.; Tian, Z.; Lai, J.; Zhu, W.; Long, B. Relating hyperspectral vegetation indices with soil salinity at different depths for the diagnosis of winter wheat salt stress. Remote Sens. 2021, 13, 250. [Google Scholar] [CrossRef]
- Das, B.; Manohara, K.K.; Mahajan, G.R.; Sahoo, R.N. Spectroscopy based novel spectral indices, PCA- and PLSR-coupled machine learning models for salinity stress phenotyping of rice. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2020, 229, 117983. [Google Scholar] [CrossRef]
- Elsayed, S.; Darwish, W.K. Hyperspectral remote sensing to assess the water status, biomass, and yield of maize cultivars under salinity and water stress. Bragantia 2017, 76, 62–72. [Google Scholar] [CrossRef]
- Tirado, S.B.; St Dennis, S.; Enders, T.A.; Springer, N.M. Utilizing top-down hyperspectral imaging for monitoring genotype and growth conditions in maize. BioRxiv 2020. [Google Scholar] [CrossRef]
- Krezhova, D.; Kirova, E. Hyperspectral remote sensing of the impact of environmental stress on nitrogen fixing soybean plants (Glycine max L.). In Proceedings of the 5th International Conference on Recent Advances in Space Technologies, Istanbul, Turkey, 9–11 June 2011; pp. 172–177. [Google Scholar]
- Peñuelas, J.; Isla, R.; Filella, I.; Araus, J.L. Visible and near-infrared reflectance assessment of salinity effects on barley. Crop Sci. 1997, 37, 198–202. [Google Scholar] [CrossRef]
- Brugger, A.; Behmann, J.; Paulus, S.; Luigs, H.G.; Kuska, M.T.; Schramowski, P.; Kersting, K.; Steiner, U.; Mahlein, A.K. Extending hyperspectral imaging for plant phenotyping to the UV-range. Remote Sens. 2019, 11, 1401. [Google Scholar] [CrossRef]
- Hamzeh, S.; Naseri, A.; Alavi Panah, S.; Mojaradi, B.; Bartholomeus, H.M.; Clevers, J.G.; Behzad, M. Estimating salinity stress in sugarcane fields with spaceborne hyperspectral vegetation indices. Int. J. Appl. Earth Obs. Geoinf. 2013, 21, 282–290. [Google Scholar] [CrossRef]
- Lara, M.Á.; Diezma, B.; Lleó, L.; Roger, J.M.; Garrido, Y.; Gil, M.I.; Ruiz-Altisent, M. Hyperspectral imaging to evaluate the effect of irrigation water salinity in lettuce. Appl. Sci. 2016, 6, 412. [Google Scholar] [CrossRef]
- Hernandez, E.I.; Melendez-Pastor, I.; Navarro-Pedreno, J.; Gomez, I. Spectral indices for the detection of salinity effects in melon plants. Sci. agric. 2014, 71, 324–330. [Google Scholar] [CrossRef]
- Feng, X.; Zhan, Y.; Wang, Q.; Yang, X.; Yu, C.; Wang, H.; Tang, Z.; Jiang, D.; Peng, C.; He, Y. Hyperspectral imaging combined with machine learning as a tool to obtain high-throughput plant salt-stress phenotyping. Plant J. 2020, 101, 1448–1461. [Google Scholar] [CrossRef]
- Li, G.; Wanb, S.; Zhoua, J.; Yanga, Z.; Qina, P. Leaf chlorophyll fluorescence, hyperspectral reflectance, pigments content, malondialdehyde and proline accumulation responses of castor bean (Ricinus communis L.) seedlings to salt stress levels. Ind. Crops Prod. 2010, 31, 13–19. [Google Scholar] [CrossRef]
- Singh, B.; Singh, J.P.; Kaur, A.; Singh, N. Phenolic compounds as beneficial phytochemicals in pomegranate (Punica granatum L.) peel: A review. Food Chem. 2018, 261, 75–86. [Google Scholar] [CrossRef]
- Naeini, M.R.; Khoshgoftarmanesh, A.H.; Fallahi, E. Partitioning of chlorine, sodium, and potassium and shoot growth of three pomegranate cultivars under different levels of salinity. J. Plant Nut. 2006, 29, 1835–1843. [Google Scholar] [CrossRef]
- Grieve, C.M.; Grattan, S.R.; Maas, E.V. Plant salt tolerance. In ASCE Manual and Reports on Engineering Practice No. 71 Agricultural Salinity Assessment and Management, 2nd ed.; Wallender, W.W., Tanji, K.K., Eds.; ASCE: Reston, VA, USA, 2012; Volume 13, pp. 405–459. [Google Scholar]
- Calzone, A.; Podda, A.; Lorenzini, G.; Maserti, B.E.; Carrari, E.; Deleanu, E.; Hoshika, Y.; Haworth, M.; Nali, C.; Badea, O.; et al. Cross-talk between physiological and biochemical adjustments by Punica granatum cv. Dente di cavallo mitigates the effects of salinity and ozone stress. Sci. Tot. Environ. 2019, 656, 589–597. [Google Scholar] [CrossRef] [PubMed]
- Karimi, H.R.; Hasanpour, Z. Effects of salinity and water stress on growth and macro nutrients concentration of pomegranate (Punica granatum L.). J. Plant Nutr. 2014, 37, 1937–1951. [Google Scholar] [CrossRef]
- Calzone, A.; Cotrozzi, L.; Pellegrini, E.; Guidi, L.; Lorenzini, G.; Nali, C. Differential response strategies of pomegranate cultivars lead to similar tolerance to increasing salt concentrations. Sci. Hortic. 2020, 271, 109441. [Google Scholar] [CrossRef]
- Khodabakhshian, R.; Emadi, B.; Khojastehpour, M.; Golzarian, M.R. Determining quality and maturity of pomegranates using multispectral imaging. J. Saudi Soc. Agric. Sci. 2017, 16, 322–331. [Google Scholar] [CrossRef]
- Khodabakhshian, R.; Emadi, B.; Khojastehpour, M.; Golzarian, M.R.; Sazgarnia, A. Non-destructive evaluation of maturity and quality parameters of pomegranate fruit by visible/near infrared spectroscopy. Int. J. Food Prop. 2017, 20, 41–52. [Google Scholar] [CrossRef]
- Khodabakhshian, R.; Emadi, B.; Khojastehpour, M.; Golzarian, M.R. A comparative study of reflectance and transmittance modes of Vis/NIR spectroscopy used in determining internal quality attributes in pomegranate fruits. J. Food Meas. Charact. 2019, 13, 3130–3139. [Google Scholar] [CrossRef]
- Arendse, E.; Fawole, O.A.; Magwaza, L.S.; Nieuwoudt, H.; Opara, U.L. Comparing the analytical performance of near and mid infrared spectrometers for evaluating pomegranate juice quality. LWT 2018, 91, 180–190. [Google Scholar] [CrossRef]
- Calzone, A.; Cotrozzi, L.; Remorini, D.; Lorenzini, G.; Nali, C.; Pellegrini, E. Oxidative stress assessment by a spectroscopic approach in pomegranate plants under a gradient of ozone concentrations. Environ. Exp. Bot. 2021, 182, 10439. [Google Scholar] [CrossRef]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 28, 239–250. [Google Scholar] [CrossRef]
- Mujeeb-Kazi, A.; Munns, R.; Rasheed, A.; Ogbonnaya, F.; Ali, N.; Hollington, P. Breeding strategies for structuring salinity tolerance in wheat. Adv. Agron. 2019, 155, 121–187. [Google Scholar] [CrossRef]
- Cotrozzi, L.; Remorini, D.; Pellegrini, E.; Guidi, L.; Nali, C.; Lorenzini, G.; Massai, R.; Landi, M. Living in a Mediterranean city in 2050: Broadleaf or evergreen ‘citizens’? Environ. Sci. Pollut. Res. 2018, 25, 8161–8173. [Google Scholar] [CrossRef] [PubMed]
- Stanton, K.M.; Micklebart, M.V. Maintenance of water uptake and reduced water loss contribute to water stress tolerance of Spiraea alba Du Roi and Spiraea tomentosa L. Hortic. Res. 2014, 1, 14033. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Chevallier, S.; Bertand, D.; Kohler, A.; Courcoux, P. Application of PLS-DA in multivariate image analysis. J. Chemom. 2006, 20, 221–229. [Google Scholar] [CrossRef]
- Kuhn, M. Building predictive models in R using the caret package. J. Stat. Softw. 2008, 28, 5. [Google Scholar] [CrossRef]
- Chen, S.; Hong, X.; Harris, C.J.; Sharkey, P.M. Sparse modeling using orthogonal forest regression with PRESS statistic and regularization. IEEE Trans. Syst. Man Cybern. B 2004, 34, 898–911. [Google Scholar] [CrossRef]
- Chong, I.G.; Jun, C.H. Performance of some variable selection methods when multicollinearity is present. Chemom. Intell. Lab. Syst. 2005, 28, 103–112. [Google Scholar] [CrossRef]
- Yendrek, C.R.; Tomaz, T.; Montes, C.M.; Cao, Y.; Morse, A.M.; Brown, P.J.; McIntyre, L.M.; Leakey, A.D.B.; Ainsworth, E.A. High-throughput phenotyping maize leaf physiological and biochemical traits using hyperspectral reflectance. Plant Physiol. 2017, 173, 614–626. [Google Scholar] [CrossRef]
- Gao, B.C. NDWI-a normalized difference water index for remote sensing of vegetation liquid from space. Remote Sens. Environ. 1996, 58, 257–266. [Google Scholar] [CrossRef]
- Merzlyak, M.N.; Gitelson, A.A.; Chivkunova, O.B.; Rakitin, V.Y. Non-destructive optical detection of pigment changes during leaf senescence and fruit ripening. Physiol. Plant. 1999, 106, 135–141. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Merzlyak, M.N. Quantitative estimation of chlorophyll a using reflectance spectra—Experiments with autumn chestnut and maple leaves. J. Photochem. Photobiol. 1994, 22, 247–252. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Zur, Y.; Chivkunova, O.B.; Merzlyak, M.N. Assessing carotenoid content in plant leaves with reflectance spectroscopy. Photochem. Photobiol. 2002, 75, 272–281. [Google Scholar] [CrossRef]
- Cavender-Bares, J.; Ackerly, D.D.; Hobbie, S.E.; Townsend, P.A. Evolutionary legacy effects on ecosystems: Biogeographic origins, plant traits, and implications for management in the era of global change. Annu. Rev. Ecol. Evol. Syst. 2016, 47, 433–462. [Google Scholar] [CrossRef]
- Campos-Medina, V.A.; Cotrozzi, L.; Stuart, J.J.; Couture, J.J. Spectral characterization of wheat functional trait responses to Hessian fly: Mechanisms for trait-based resistance. PLoS ONE 2019, 14, e0219431. [Google Scholar] [CrossRef]
- Begum, H.; Alam, M.S.; Feng, Y.; Koua, P.; Ashrafuzzman, M.; Shrestha, A. Genetic dissecation of bread wheat diversity and identification of adaptive loci in response to elevated tropospheric ozone. Plant Cell Environ. 2020, 43, 2650–2665. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [PubMed]
- Peñuelas, J.; Garbulsky, M.F.; Filella, I. Photochemical reflectance index (PRI) and remote sensing of plant CO2 uptake. New Phytol. 2011, 191, 596–599. [Google Scholar] [CrossRef] [PubMed]
- Zarco-Tejada, P.J.; Miller, J.R.; Mohammed, G.H.; Noland, T.L.; Sampson, P.H. Estimation of chlorophyll fluorescence under natural illumination from hyperspectral data. Int. J. Appl. Earth Obs. 2001, 3, 321–327. [Google Scholar] [CrossRef]
- Jia, M.; Li, D.; Colombo, R.; Whang, Y.; Whang, X.; Cheng, T.; Zhu, Y.; Yao, X.; Xu, C.; Ouer, G.; et al. Quantifying chlorophyll fluorescence parameters from hyperspectral reflectance at the leaf scale under various nitrogen treatment regimes in winter wheat. Remote Sens. 2019, 11, 2838. [Google Scholar] [CrossRef]
- Serbin, S.P.; Dillaway, D.N.; Kruger, E.L.; Townsend, P.A. Leaf optical properties reflect variation in photosynthetic metabolism and its sensitivity to temperature. J. Exp. Bot. 2012, 63, 489–502. [Google Scholar] [CrossRef]
- Heckmann, D.; Schlüter, U.; Weber, A.P.M. Machine learning techniques for predicting crop photosynthetic capacity from leaf reflectance spectra. Mol. Plant. 2017, 10, 878–890. [Google Scholar] [CrossRef]
- Fu, P.; Meacham-Hensold, K.; Guan, K.; Wu, J.; Bernacchi, C. Estimating photosynthetic traits from reflectance spectra: A synthesis of spectral indices, numerical inversion, and partial least square regression. Plant Cell Environ. 2020, 43, 1241–1258. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.L.; Steven, M.D.; Colls, J.J. Use of hyperspectral derivative tools in red-edge region to identify plant stress response to gas leaks. Remote Sens. Environ. 2004, 92, 207–212. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; Miller, J.R.; Morales, A.; Berjón, A.; Agüera, J. Hyperspectral indices and model simulation for chlorophyll estimation in open-canopy tree crops. Remote Sens. Environ. 2004, 9, 463–476. [Google Scholar] [CrossRef]
- Mutanga, O.; Skidmore, A.K. Red edge shift and biochemical content in grass canopies. ISPRS J. Photogram. Remote Sens. 2007, 62, 34–42. [Google Scholar] [CrossRef]
- Merzlyak, M.N.; Gitelson, A.A.; Chivkunova, O.B.; Solovchenko, A.E.; Pogosyan, S.I. Application of reflectance spectroscopy for analysis of higher plant pigments. Russ. J. Plant Physiol. 2003, 50, 704–710. [Google Scholar] [CrossRef]
- Jones, H.G. Monitoring plant and soil water status: Established and novel methods revisited and their relevance to studies of drought tolerance. J. Exp. Bot. 2007, 58, 119–130. [Google Scholar] [CrossRef]
- Curran, P.J. Remote sensing of foliar chemistry. Remote Sens. Environ. 1989, 30, 271–278. [Google Scholar] [CrossRef]
- Shetty, N.; Gislum, R. Quantification of fructan concentration in grasses using NIR spectroscopy and PLSR. Field Crops Res. 2011, 120, 31–37. [Google Scholar] [CrossRef]
- Rubert-Nason, K.F.; Holeski, L.M.; Couture, J.J.; Gusse, A.; Undersander, D.J.; Lindroth, R.L. Rapid phytochemical analysis of birch (Betula) and poplar (Populus) foliage by near-infrared reflectance spectroscopy. Anal. Bioanal. Chem. 2013, 405, 1333–1344. [Google Scholar] [CrossRef] [PubMed]
- Asner, G.P.; Martin, R.E. Spectroscopic remote sensing of non-structural carbohydrates in forest canopies. Remote Sens. 2015, 7, 3526–3547. [Google Scholar] [CrossRef]
- Ramirez, J.A.; Posada, J.M.; Handa, I.T.; Hoch, G.; Vohland, M.; Messier, C.; Reu, B. Near-infrared spectroscopy (NIRS) predicts non-structural carbohydrate concentrations in different tissue types of a broad range of tree species. Methods Ecol. Evol. 2015, 6, 1018–1025. [Google Scholar] [CrossRef]
- Olmo, A.; García-Sánchez, F.; Simon, I.; Lidon, V.; Alfosea-Simon, M.; Cámara-Zapata, J.M.; Nicolás, J.J.; Simón-Grao, S. Characterization of the ecophysiological responses of three pomegranate cultivars to salinity. Photosynthetica 2019, 4, 1015–1024. [Google Scholar] [CrossRef]
- Borzouei, A.; Kafi, M.; Akbari-Ghogdi, E.; Mousavi-Shalmani, M. Long term salinity stress in relation to lipid peroxidation, super oxide dismutase activity and proline content of salt-sensitive and salt-tolerant wheat cultivars. Chil. J. Agric. Res. 2012, 72, 476–482. [Google Scholar] [CrossRef]
- Liu, C.; Zhao, X.; Yan, J.; Yuan, Z.; Gu, M. Effects of salt stress on growth, photosynthesis, and mineral nutrients of 18 pomegranate (Punica granatum) cultivars. Agronomy 2020, 10, 27. [Google Scholar] [CrossRef]
- Ben-Abdallah, S.; Aung, B.; Amyot, L. Salt stress (NaCl) affects plant growth and branch pathways of carotenoid and flavonoid biosyntheses in Solanum nigrum. Acta Physiol. Plant. 2016, 38, 72. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effect | df | F | p |
|---|---|---|---|
| Cultivar | 1 | 15.24 | *** |
| Time | 4 | 11.22 | *** |
| Salinity | 1 | 23.74 | *** |
| Cultivar × time | 4 | 0.26 | ns |
| Cultivar × salinity | 1 | 0.83 | ns |
| Time × salinity | 4 | 6.36 | *** |
| Cultivar × time × salinity | 4 | 1.88 | ns |
| Effect | Comp | Cal | Val | ||
|---|---|---|---|---|---|
| Accuracy | Kappa | Accuracy | Kappa | ||
| Cultivar | 39 | 1.00 ± 0.00 | 1.00 ± 0.00 | 0.89 ± 0.05 | 0.79 ± 0.11 |
| Time | 26 | 0.86 ± 0.02 | 0.83 ± 0.02 | 0.66 ± 0.07 | 0.58 ± 0.09 |
| Salinity | 44 | 0.90 ± 0.02 | 0.81 ± 0.04 | 0.79 ± 0.07 | 0.58 ± 0.15 |
| Time × salinity | 60 | 0.83 ± 0.02 | 0.81 ± 0.03 | 0.53 ± 0.08 | 0.48 ± 0.08 |
| Parameter | Range (nm) | Comp | Cal | Val | |||||
|---|---|---|---|---|---|---|---|---|---|
| R2 | RMSE | %RMSE | R2 | Bias | RMSE | %RMSE | |||
| Fv/Fm | 400–1200 | 12 | 0.94 ± 0.01 | 0.01 ± 0.00 | 5 | 0.64 ± 0.18 | 0.00 ± 0.01 | 0.03 ± 0.01 | 13 |
| ΦPSII | 400–1200 | 11 | 0.92 ± 0.01 | 0.02 ± 0.00 | 6 | 0.61 ± 0.18 | 0.00 ± 0.01 | 0.04 ± 0.01 | 13 |
| qP | 400–700 | 13 | 0.94 ± 0.01 | 0.02 ± 0.00 | 5 | 0.68 ± 0.16 | 0.00 ± 0.01 | 0.04 ± 0.01 | 12 |
| qN | 400–800 | 12 | 0.93 ± 0.01 | 0.02 ± 0.00 | 6 | 0.68 ± 0.15 | 0.00 ± 0.02 | 0.05 ± 0.01 | 14 |
| RWC | 1400–2400 | 13 | 0.94 ± 0.01 | 2.26 ± 0.19 | 6 | 0.71 ± 0.15 | 0.05 ± 1.76 | 5.00 ± 1.05 | 13 |
| Ψπ | 1400–2400 | 12 | 0.98 ± 0.01 | 0.06 ± 0.01 | 3 | 0.79 ± 0.13 | 0.00 ± 0.06 | 0.17 ± 0.04 | 9 |
| Effect | df | Fv/Fm | ΦPSII | qP | qN | sPRI | NDVI | sPSRI | Ψw | Ψπ | RWC | NDWI | MDA | ORAC | Phen | Ant | CI | CRI |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cultivar | 1 | 28.04 | 121.19 | 43.92 | 16.94 | 2.7 | 30.96 | 31.91 | 0.01 | 30.45 | 3.84 | 57.04 | 28.47 | 1.76 | 8.19 | 0 | 27.04 | 38.48 |
| *** | *** | *** | *** | ns | *** | *** | ns | *** | ns | *** | *** | ns | ** | ns | *** | *** | ||
| Time | 4 | 11.71 | 9.66 | 13.89 | 11.06 | 22.63 | 7.52 | 0.47 | 49.27 | 6.71 | 10.05 | 1.55 | 13.6 | 44.64 | 15.53 | 4.22 | 29.45 | 28.81 |
| *** | *** | *** | *** | *** | *** | ns | *** | *** | *** | ns | *** | *** | *** | ** | *** | *** | ||
| Salinity | 1 | 15.7 | 78.58 | 136.6 | 113.05 | 57 | 4.11 | 2.29 | 221.4 | 64.52 | 11.05 | 29 | 3.1 | 123.02 | 0.06 | 1.25 | 5.79 | 2.12 |
| *** | *** | *** | *** | *** | * | ns | *** | *** | *** | *** | ns | *** | ns | ns | * | ns | ||
| Cultivar × time | 4 | 2.39 | 0.69 | 4.46 | 4.57 | 10.63 | 5.98 | 2.04 | 1.1 | 3.97 | 0.93 | 5.13 | 11.98 | 5.82 | 0.89 | 3.29 | 17.87 | 3.36 |
| ns | ns | ** | ** | *** | *** | ns | ns | ** | ns | *** | *** | *** | ns | * | *** | * | ||
| Cultivar × salinity | 1 | 2.86 | 9.98 | 11.9 | 5.81 | 0.28 | 4.27 | 0.04 | 0.26 | 3.13 | 3.72 | 46.89 | 13.6 | 0 | 1.96 | 0.75 | 8.5 | 16 |
| ns | ** | *** | * | ns | * | ns | ns | ns | ns | *** | *** | ns | ns | ns | *** | *** | ||
| Time × salinity | 4 | 1.32 | 7.12 | 4.38 | 11.03 | 9.96 | 0.65 | 0.15 | 25.75 | 21.75 | 1.16 | 1.27 | 12.18 | 19.97 | 2.86 | 1.95 | 5.61 | 2.19 |
| ns | *** | ** | *** | *** | ns | ns | *** | *** | ns | ns | *** | *** | * | ns | *** | ns | ||
| Cultivar × time × salinity | 4 | 1.86 | 3.13 | 3.31 | 3.55 | 3.92 | 3.71 | 2.3 | 1.33 | 4.63 | 2.84 | 2.03 | 9.72 | 6.48 | 2.28 | 3.43 | 3.66 | 1.49 |
| ns | * | * | ** | ** | ** | ns | ns | ** | * | ns | *** | *** | ns | * | ** | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calzone, A.; Cotrozzi, L.; Lorenzini, G.; Nali, C.; Pellegrini, E. Hyperspectral Detection and Monitoring of Salt Stress in Pomegranate Cultivars. Agronomy 2021, 11, 1038. https://doi.org/10.3390/agronomy11061038
Calzone A, Cotrozzi L, Lorenzini G, Nali C, Pellegrini E. Hyperspectral Detection and Monitoring of Salt Stress in Pomegranate Cultivars. Agronomy. 2021; 11(6):1038. https://doi.org/10.3390/agronomy11061038
Chicago/Turabian StyleCalzone, Antonella, Lorenzo Cotrozzi, Giacomo Lorenzini, Cristina Nali, and Elisa Pellegrini. 2021. "Hyperspectral Detection and Monitoring of Salt Stress in Pomegranate Cultivars" Agronomy 11, no. 6: 1038. https://doi.org/10.3390/agronomy11061038
APA StyleCalzone, A., Cotrozzi, L., Lorenzini, G., Nali, C., & Pellegrini, E. (2021). Hyperspectral Detection and Monitoring of Salt Stress in Pomegranate Cultivars. Agronomy, 11(6), 1038. https://doi.org/10.3390/agronomy11061038