1. Introduction
Drought, one of the major types of environmental abiotic stresses, restricts plant performance throughout its life cycle. It limits the productivity of agricultural crops, mostly in dry areas (e.g., semi-arid and arid regions), including Egypt and Saudi Arabia, thus causing an increasing problem over time due to the ongoing climate change. Deficit irrigation water (DIw) adversely affects plant growth, development, and production through decreasing water flux, closure of stomata, and reduced CO
2 fixation [
1,
2,
3]. DIw strongly affects plant antioxidant system, metabolism, and physio-biochemistry [
4,
5,
6,
7,
8]. It causes photooxidation damage to cell components by over-generating reactive species of oxygen (ROS), including radicals of O
2•− and H
2O
2 and eventually photo-inhibition [
9], which cause significant oxidative damage to proteins, lipids, and DNA, thus discouraging plant growth [
10]. To mitigate the oxidative stress damage, complex ROS-scavenging antioxidant system (enzymes (SOD, POD, CAT, etc.) and low-molecular mass compounds (GSH, AsA, carotenoids, phenols, etc.)) are developed/adopted in plants [
11]. The enzymatic antioxidants can directly scavenge ROS or produce one or more of the non-enzymatic antioxidants. SOD is specifically implicated in converting O
2•− to H
2O
2 in cell cytoplasm, chloroplasts, mitochondria, and peroxisomes. In addition, POD acts as a scavenger of H
2O
2 and CAT eliminates the H
2O
2 in mitochondria and micro-bodies [
12], helping mitigate the adverse effects of oxidative stress.
The effective reduction of cell osmotic potential maintains plant cell metabolism and avoids dehydration damage induced by DIw stress via the accumulation of more levels of glycinebetaine, soluble sugars, and proline [
13]. These osmotic compounds maintain an osmotic balance, efficacy, and normality of membrane functions, and protect macromolecules and plasma membranes, providing cellular resistance against DIw stress and dehydration [
10,
14,
15].
In most cases, endogenous components of the developed/adopted antioxidant system in plants are not sufficient to defend against environmental stress. Consequently, plant extract and microbial bio-stimulants may be applied exogenously as an adjuvant to plants to increase their tolerance to stress. Plant extracts can induce plant resistance under various stresses, and microbial biostimulants can increase gene regulation in plants related to drought stress and can also produce biofilm, which helps plants survive under drought stress [
16,
17,
18,
19,
20,
21].
At present, extracts containing natural bio-stimulants obtained from maize grains have been applied for seed priming and have been reported to enhance plant performances under different stress conditions; salinity [
22,
23], nutrient deficiencies [
24], cadmium [
25], and salinity plus drought [
26]. MGE is an important organic bio-stimulant for different stressed plants. It is rich in various phytohormones (e.g., cytokinins, auxins, and gibberellins), antioxidants, and essential nutrients. It has been used to enhance plant morphology, physiology, biochemistry, and induce plant tolerance against stress damages [
22,
25]. To date, no work has been done using MGE enriched with antioxidants such as polyamines for wheat grown under DIw stress. Thus, this is the first report that used MGE to stimulate the growth of pretreated wheat plants under DIw stress due to its high content of antioxidants, plant hormones, vitamins, polyamines, and nutrients, making it an effective biological stimulant [
22,
24,
25,
26,
27] (
Table 1). These reports stated that MGE confers seed and their emerged seedlings the ability to resist the unfavorable effects of stress through elevating wheat performance (growth and productivity), physio-biochemical parameters, and different antioxidants. Thus, MGE is an innovative means to provide seeds and developing seedlings that are resistant to damages caused by environmental stresses such as DIw.
Wheat (
Triticum aestivum L.) grain and straw are major sources of nourishing the diets of humans and animals, respectively in urban and rural communities due to their high protein and calorie content (approximately 82–85%). However, environmental stresses severely affect wheat productivity; if the available soil water reaches 45–50% of field capacity in the root zone, the wheat plants must be watered [
2,
26]. The duration and severity of stress, growth physio-biochemical processes, and patterns of differential gene expression are some of the factors that can influence the wheat plant’s response to DIw stress [
23,
26,
28,
29].
Although MGE has been used in some investigations for some crop plants to boost their antioxidant defense system against the stress impacts of certain abiotic stresses [
22,
24,
25,
26,
27], there is no information available on seed priming using MGE (as a bio-stimulant) to encourage plant growth and production by mitigating the damage caused by drought stress. However, by analyzing MGE, we found that it is rich in phytohormones, especially zeatins, but was poor in PAs (
Table 1), so enriching this extract with PAs could increase its effectiveness. Therefore, this work is based on the hypothesis that seed soaking in MGE enriched in PAs will mitigate drought stress by enhancing the growth, yield, and physio-biochemical systems of wheat plants. Thus, in this study, the effect of wheat seed priming in MGE (without or with enrichment by PAs) on growth and yield, physio-biochemical characteristics, including the up-regulation of AsA–GSH cycle, glyoxalase system, and PAs gene expression of wheat plants was studied under DIw stress conditions.
4. Discussion
No information is available on soaking wheat seeds in maize grain extract enriched with anti-stress stimulants such as polyamines [PAs (MGE
PA)] to alleviate the adverse impacts of drought stress. In this study, improvements in wheat plant growth and yield under the effects of drought stress were obtained through improvements in physiological and biochemical attributes due to seed priming in MGE or MGE
PA with the advantage of MGE
PA. Some articles reported on significant changes in antioxidant defense system including enzymatic and non-enzymatic antioxidants after exposing different plants to drought stress [
7,
8,
16,
63] In the current study, this finding corresponds to the measured up-regulation of the ascorbate–glutathione (AsA-GSH) cycle, glyoxalase system, trans- and cis-zeatin, and PAs and their biosynthetic genes under drought stress in response to soaking wheat seeds in MGE or MGE
PA. In this regard, the results acquired with MGE
PA exceeded those gained with MGE due to enrichment of MGE
PA with PAs that increased the efficiency of the extract. Nevertheless, previous studies indicated the importance of MGE (the extract without enrichment with PAs) in mitigating the adverse effects of some stresses (salinity, cadmium, sandy state, and salinity+drought) in some crop plants [
22,
24,
25,
26,
27], but this study reported a higher benefit of MGE
PA than MGE.
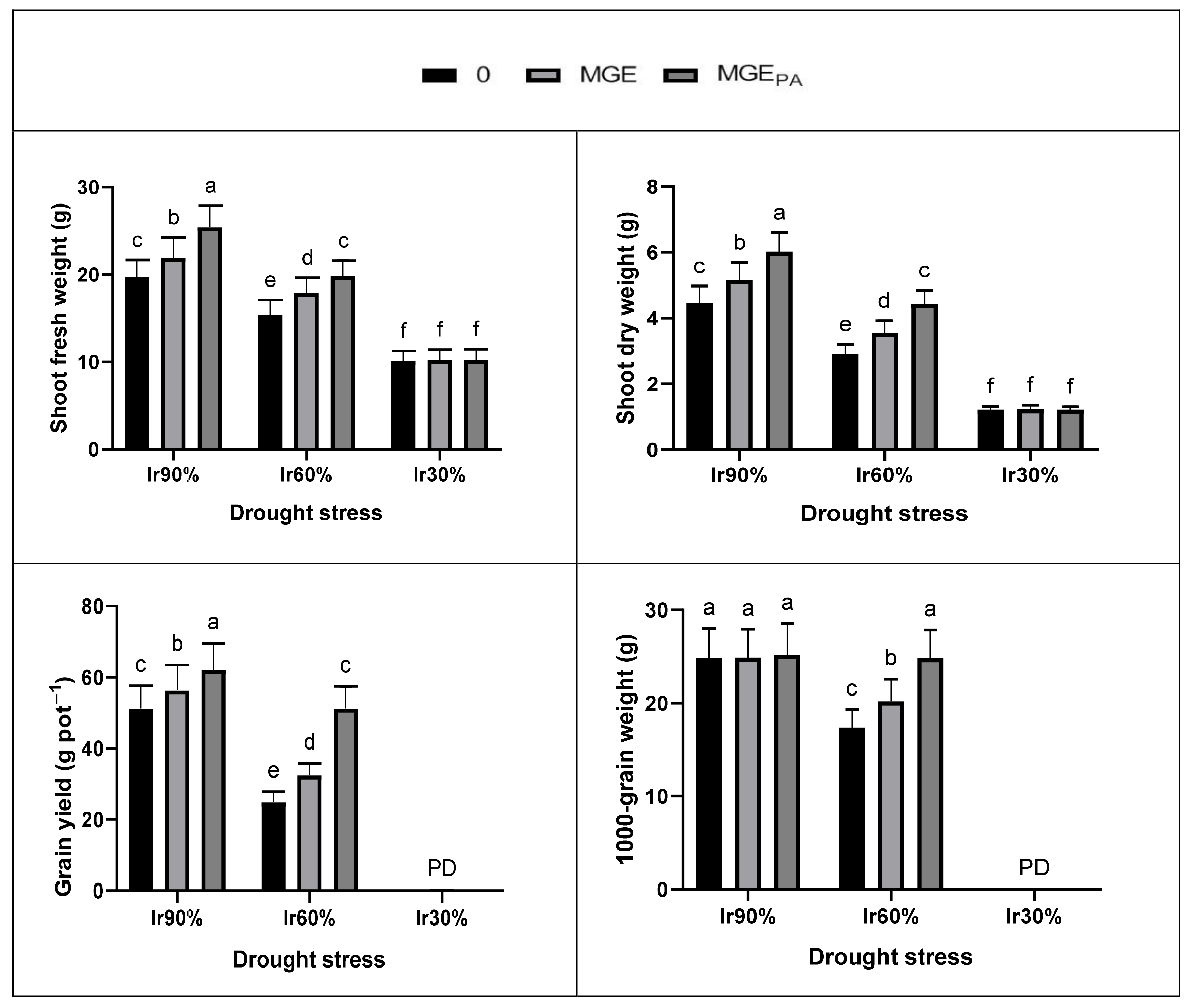
In this study, drought stress in terms of deficit irrigation water (DIw) was detrimental to the growth and yield of wheat plants. Damage to wheat growth and yield due to Ir30% (severe drought; 30% of soil relative water content; SRWC) was more severe than that from Ir60% (moderate drought; 60% of SRWC) (
Figure 1 and
Figure 2,
Table 9). The plants did not survive under severe drought even with pretreatment with both extracts (plants died during the flowering stage;
Figure 1). This negative finding can be attributed to the drought affecting plant physiology and biochemistry (
Figure 1 and
Figure 2,
Table 2,
Table 3,
Table 4,
Table 5,
Table 6,
Table 7,
Table 8 and
Table 9). In this study, the decreased growth and yield of drought-stressed wheat plants associated with the overproduction of reactive species of oxygen (ROS), especially H
2O
2 and O
2•− (
Table 2,
Table 3 and
Table 9). This finding is consistent with that obtained by Noctor et al. [
64] and Wang et al. [
65]. The plant’s first response to drought is closed leaf stomata to reduce loss of water through transpiration (
Table 2,
Table 3 and
Table 9). As a result of continuous photosynthesis in light, the depletion of intercellular CO
2 concentration is facilitated by an increased gas diffusion barrier. Therefore, the low CO
2 availability catalyzes the oxygenation of ribulose-1,5-bisphosphate, thereby increasing the production of photorespiratory H
2O
2 in cell peroxisomes [
64]. Drought treatments negatively affected leaf photosynthetic pigments, photosynthetic efficiency, gas exchange parameters, cell membranes (increase in lipid peroxidation; MDA and ionic leakage; EL, and decrease in stability index), RWC, zeatins, osmoprotectant and PAs contents, antioxidant enzyme, AsA-GSH cycle and glyoxalase system activities, and transcription of PAs biosynthetic genes (
Figure 1 and
Figure 2,
Table 2,
Table 3,
Table 4,
Table 5,
Table 6,
Table 7,
Table 8 and
Table 9). Drought-stimulated oxidative stress (increased production of H
2O
2 and O
2•−;
Table 2,
Table 3 and
Table 9) caused these negative findings. Over Ir60%, Ir30% caused exacerbation of MDA and EL and severely affected PSII function efficiency (
Figure 2 and
Table 9), causing stomatal closure (
Table 2 and
Table 9), which disrupted the plant antioxidant system (
Table 4,
Table 5,
Table 6,
Table 7,
Table 8 and
Table 9), and eventually, the plant did not survive.
However, the advantage of seed soaking in both extracts (MGE or MGE
PA) was reported under moderate drought (Ir60%). The higher benefit of MGE
PA over MGE can be attributed to the additional benefits of PAs granted through pretreatment with MGE
PA, where PAs play important roles in mitigating the damage caused by different stresses on crop plants [
29,
66,
67,
68]. Yildiztugay et al. [
69] reported a relationship between the plant’s biomass (growth) and its water content. Similarly, our results displayed this relationship (
Figure 1 and
Figure 2,
Table 2,
Table 3 and
Table 9). Despite the benefit of pretreatment with MGE, MGE
PA enabled wheat plants to function normally under Ir60% and revealed growth (biomass) and leaf relative water content (RWC), and thus yield similar to that gained by the optimum irrigated plants (Ir90%; control). Wheat plants pretreated with MGE
PA and subjected to Ir60% had leaf photosynthetic pigments, photosynthesis efficiency (Fv/Fm, PI, and hill reaction activity; HRA), and gas exchange parameters identical to those obtained with well-watered plants (
Figure 1 and
Figure 2,
Table 2,
Table 3 and
Table 9). The damage caused by Ir60% was repaired with MGE
PA. During the soaking process, the seeds can absorb the bioactive ingredients from the extract such as antioxidants (AsA and GSH) and hormones (salicylic acid and cytokinins (CKs), especially
t-Z and
c-Z), in addition to the extract PAs (
Table 1). These bioactive ingredients enrich the seeds and accelerate metabolism, giving them robust germination and strong seedlings to withstand drought stress.
Under Ir60%, MGE
PA preserved leaf photosynthetic pigments and photosynthetic efficiency along with gas exchange parameters, which contributed to the maintenance of wheat yield (
Figure 1 and
Figure 2,
Table 2,
Table 3 and
Table 9). In this regard, Farooq et al. [
16] reported a positive correlation between chlorophyll contents and grain yield in wheat plants. Moreover, the maintenance of PSII function (HRA), antioxidant system, and biosynthesis genes of PAs (
Table 4,
Table 5,
Table 6,
Table 7,
Table 8 and
Table 9) along with the beneficial impacts of the bioactive stimulants present in MGE
PA (
Table 1) contributed well to the plant’s performance under drought stress. Under Ir60%, some antioxidant compounds supported MGE
PA-pretreated plants to cause cell membrane stabilization by suppressing H
2O
2 and O
2•− levels, thus reducing MDA and EL levels (
Table 3 and
Table 9). In this study, as drought stress caused a reduction in chlorophyll content, it caused leaf yellowing and senescence (data not shown). This finding can be attributed to a degradation of chlorophyll and a dysregulation of the photosynthetic apparatus, including decreased HRA, and thus a reduction in wheat yield. Nevertheless, MGE
PA enabled wheat plants to stay green by maintaining the chlorophyll content for as long as possible, thus producing better under drought stress (
Figure 2,
Table 9) due to the bioactive stimulants present in MGE
PA. This finding is consistent with that obtained by [
22,
24,
25,
26,
27] under different abiotic stress conditions. Drought-stressed wheat plant becomes better performer (in terms of growth and yield characteristics) when pretreated with MGE, which has a high ROS-scavenging activity (88.6 ± 1.6%) due to the high contents of different antioxidants, plant hormones, and PAs present in MGE (
Table 1). The performance of stressed plant becomes higher with MGE
PA due to the PAs added to MGE. Ghassemi et al. [
70] reported that PAs improved root growth and development and adjusted cellular water potential after emergence, leading to increased nutrient uptake and translocation in drought-stressed plants. In addition, the enhanced translocation of assimilates, which are photosynthesized in plant leaves, to the edible portion (the grains) is attributed to the longer duration of photosynthesis due to the green leaf survival merit, resulting in a longer period of grain filling under stress [
16]. This finding is in line with the finding of this study, which is likely due to the ability of MGE
PA-pretreated wheat plants to stimulate the activity of the antioxidant system to withstand DIw stress (
Table 4,
Table 5,
Table 6,
Table 7,
Table 8 and
Table 9) and enable the plants to perform better with regard to the meristematic tissue activities, including the stimulation of cell division and expansion due to maintaining adequate water (
Figure 1 and
Figure 2,
Table 2,
Table 3 and
Table 9). As a result, it becomes possible to obtain a satisfactory improvement in the growth of drought-stressed plants due to the richness of MGE, especially after its enrichment with PAs, in phytohormones, auxins, CKs including zeatins, gibberellins (GAs), and PAs (
Table 1). Therefore, hormonal homeostasis could be induced with MGE
PA as a potential mechanism for boosting DIw stress tolerance in plants. This hormonal homeostasis mechanism could function through a complex cross-talk among auxins, CKs and GAs, as well as among plant hormones, PAs, and other biostimulants as an anti-stress network in favor of the plant’s response to DIw stress.
In this study, under low availability of water, MGE
PA pretreatment replaced the detrimental effects of DIw to marked increases in wheat growth and yield due to the increase in gas exchange parameters, photosynthetic efficiency including HRA, and the biosynthesis of chlorophyll catalyzed by the biostimulants present in MGE
PA, which contributed to elimination of the biomarkers of oxidative stress (H
2O
2 and O
2•−) and preservation of cell membranes and cell water content due to reduced levels of EL and MDA accumulation (
Figure 1 and
Figure 2,
Table 2,
Table 3 and
Table 9). Different stresses cause a detrimental effect on EL and membrane lipids due to overproduced ROS, which disrupt chlorophyll biosynthesis due to disruption of the pigment protein complex functioning, de novo protein biosynthesis, and chlorophyll-associated components [
8,
16,
71,
72]. However, MGE
PA bioactive promoters eliminated these harmful effects and mediated improvement in chlorophyll biosynthesis (
Figure 1 and
Figure 2,
Table 2,
Table 3 and
Table 9). MGE
PA pretreatment can stimulate the accumulation of different plant hormones in germinated seeds and thus seedlings, especially CKs, to improve the biosynthesis of chlorophylls as shown in Cortleven et al. [
73]. In this study, MGE
PA pretreatment improved photosynthesis efficiency and PSII activity (Fv/Fm, PI, and HRA), which could be attributed to suppressing the formation of ROS (
1O
2, H
2O
2, and O
2•‒), thus protecting the chloroplast structure from DIw-induced oxidative stress damage (
Figure 1 and
Figure 2,
Table 2,
Table 3 and
Table 9). In this regard, Tikkanen et al. [
74] attributed a similar result to the safeguarded light reaction functioning in photosynthesis process, which is functionally synchronized with PSI and PSII enzymes. The functioning of both photosystems is connected to the generation of trans-thylakoid proton, which creates a pH difference for regulating the transfer of electrons to PSI under direct monitoring of the PGR5 protein. As a result of its richness in bioactive stimuli, especially plant hormones, MGE
PA can enhance the biosynthesis of polysaccharides and proteins implicated in photosynthesis regulation, in addition to mediating the synthesis of components of redox that may have contributed to photo-protection. In addition, MGE
PA can increase carotenoids synthesis that can protect the photosynthesis system from the overproduction of ROS by up-regulating pigment-synthesizing enzyme activities associated with reducing the degradation of enzymes [
25,
26,
75]. This result is consistent with Aldesuquy [
76], who reported a positive synergistic impact of plant hormones on pigment and fine chloroplast structure in the flag leaf of stressed wheat plant.
As previously obtained in some works [
22,
24,
25,
26,
27], reduced levels of H
2O
2 and O
2•− and thus reduced lipid peroxidation and EL were obtained with the MGE
PA-pretreated plant under DIw stress (
Table 3 and
Table 9). The MGE-mediated enhanced membrane integrity (due to reduced membrane EL and MDA) could be attributed to the bioactive components in present MGE
PA (
Table 1), which contributed to the maintenance of the components of the antioxidant system, in addition to the low peroxidation levels, which were considerably influenced by DIw stress. MGE
PA pretreatment considerably enhanced the activity of antioxidant enzymes and raised the contents of AsA and glutathione forms (e.g., GSH and GSSG), thus protecting the wheat plants from the oxidative stress (H
2O
2 and O
2•−) stimulated by DIw. Similar results are previously gained [
22,
24,
25,
26,
27] under different stresses. Integrally, after dismutating O
2•− to H
2O
2 by SOD, APX and CAT convert H
2O
2 to H
2O and O
2. This mechanism reduces the formation of hydroxyl; OH
‒ radicals [
77]. Since MGE
PA is a rich source of bioactive stimulants, it may stimulate SOD up-regulation to further dismutate O
2•− to H
2O
2. Plants pretreated with MGE
PA possessed stimulated AsA and GSH accumulations, which could protect stressed plants from ROS damage stimulated by DIw. All antioxidants (GR, APX, MDHAR, DHAR, GSH, and AsA) are components of the ROS-scavenging pathway (the AsA-GSH cycle), which can be stimulated and up-regulated by MGE
PA to elevate tolerance strategies against any potential damage from oxidative stress in the wheat plant (
Table 6,
Table 7,
Table 8 and
Table 9). This robust enhancement in the antioxidant system in the DIw-stressed wheat plants pretreated with MGE
PA resulted in a marked decrease in ROS accumulation (e.g., H
2O
2 and O
2•−) through the AsA‒GSH cycle and thus increased the protection of pathways of photosynthesis, resulting in better performance (e.g., growth and yield productivity) of wheat plants (
Table 6,
Table 7,
Table 8 and
Table 9). The AsA‒GSH cycle includes a sequence of redox reactions, which includes bioactive participation of NADPH, GSH, and AsA [
78]. Through this cycle, H
2O
2 molecules in cell cytosol and chloroplasts are scavenged by APX and CAT, thus preventing H
2O
2 diffusion to other organelles to avoid any damage. The improved functionality of the ASA–GSH cycle pathway due to pretreatment with MGE
PA effectively maintained redox components, including GSH and AsA. These redox components reduced the effects of oxidative stress (H
2O
2 and O
2•−) stimulated by DIw. In this study, the increased antioxidant system activity was accompanied by an enhanced tolerance to DIw stress in the wheat plant. Similar results were gained by [
16,
25,
79]. DHAR and MDHAR activities, which may be up-regulated after pretreatment with MGE
PA, improved AsA and GSH to integrate the functions of other enzymes such as GR and APX with non-enzymatic antioxidant components. As a result, a comprehensive impact occurred on H
2O
2 neutralization and the availability of NADP to effectively protect the electron transport chain [
80]. In this study, pretreatment with MGE
PA bioactive stimulants protected the electron transport chain implicated in photosynthesis, which may be attributed to the up-regulation of the NADP
+/NADPH ratio. This mechanism prevents the flow of electrons into O
2 to restrict O
2•− production [
81]. This study provides a preventive role of MGE
PA, given its richness in bioactive stimuli including plant hormones, for the photosynthesis system through the up-regulation of the antioxidant system components.
High levels of GSH can contribute significantly to MGE
PA-pretreated wheat plants to maintain the glycoxylase system for methylglyoxal elimination, which may reduce the chances of any dangerous genotoxic influence [
82]. Glyoxylase I and II (Gly I and Gly II) constitute the main enzymatic components of the ‘glyoxylase system’. MGE
PA bioactive stimuli-stimulated up-regulation in the enzymatic activities of glyoxylase system could be associated with increased levels of GSH. This may have exploited the improved influences of methylglyoxal such as crosstalk with important signalling molecules such as Ca
2+, ABA, and ROS [
83]. This is the first report conducted with MGE
PA for DIw-stressed wheat plants, in which the MGE
PA bioactive stimuli can optimize methylglyoxal. Gly I utilizes GSH as a co-factor to convert the methylglyoxal to
S-D-lactoylglutathione. In addition, Gly II produces GSH to contribute to redox homeostasis and protection against toxic species [
84]. It can be concluded that improvement in endogenous GSH by pretreatment using MGE
PA bioactive stimuli and the functioning of methylglyoxal scavenging system could enhance stress tolerance in wheat plants. Ahanger et al. [
85] explained up-regulation and the improvement of glyoxylase system activity due to exogenous treatment with kinetin. This result is consistent with our results through the application of CKs-containing MGE
PA, which improved activity of glyoxylase system (
Table 6 and
Table 9). This enhanced glycosylase system activity may protect the system of electron transport by inhibiting any damage to chloroplasts and mitochondrial ultra-structures [
82].
In this study, DIw encouraged osmoprotectant accumulations (e.g., GB, S. sugars, and proline), and these accumulations were further enhanced due to pretreatment with MGE
PA bioactive stimulants. This accumulation of osmoprotectants may occur due to their absorption from MGE
PA and/or their increased biosynthesis catalyzed by MGE
PA in soaked seeds then seedlings. These increased osmoprotectants may give the seeds the driving force for strong germination and thus vigorous seedling growth under stress. Improvements of these osmolytes may be effective mechanisms to increase plant tolerance to DIw stress to maintain adequate cell water for healthy metabolism (
Table 2,
Table 3,
Table 4,
Table 5,
Table 6,
Table 7,
Table 8 and
Table 9). The up-regulation of proline-synthesizing enzymes along with the down-regulation of proline-catabolizing enzymes and/or low incorporation into proteins leads to proline accumulation [
86,
87]. The improvement in GB, S. sugars, and proline accumulations as a result of MGE
PA pretreatment resulted in the maintenance of water balance in DIw-stressed plants to help them withstand DIw stress and avoid increased EL and MDA in plant tissues for healthy metabolic processes, protection of protein turnover, enzymatic activities, and expression of stress protective proteins. These results indicate the protective role of MGE
PA for healthy metabolic pathways and effective osmoregulation. Similar results were obtained by Ahanger and Agarwal [
85] and Thakur and Sharma [
88].
Bypassing MGE, MGE
PA attenuated DIw stress and helped the photosynthetic system to perform efficiently (
Figure 2,
Table 9) in favor of plant metabolism [
27], resulting in a lot of savings including hormonal contents (e.g., CKs such as
t-Z and
c-Z) in DIw-stressed plants (
Table 6 and
Table 9). Therefore, zeatins play a fundamental role in wheat plant response to DIw stress. Mediating plant response to stress, specifically CKs and their signaling components, predominantly regulate plant defense reactions [
89].
T-Z and
c-Z have been reported to modulate the wheat plant’s defense response to stress through several mechanisms including defense genes and hormonal regulation [
26]. This result confirms the results of this study with
t-Z and
c-Z-containing MGE
PA (
Table 6,
Table 7,
Table 8 and
Table 9). This study reported significant increases in trans-zeatin and cis-zeatin contents with MGE
PA pretreatment and supported wheat plant’s tolerance to DIw stress (
Figure 1 and
Figure 2,
Table 2,
Table 3,
Table 4,
Table 5,
Table 6,
Table 7,
Table 8 and
Table 9). Previous reports indicated increased
t-Z activity compared to cis-zeatin, which was demonstrated in this study with a higher trans-zeatin content compared to
c-Z, which may lead to an increase in the physiological role of
t-Z to stimulate a higher increase in PAs contents and their gene expressions under DIw stress due to pretreatment of the plant with MGE
PA (
Table 6,
Table 7,
Table 8 and
Table 9). This finding may be attributed to the transport, degradation, and conjugation processes of these hormones [
90,
91].
In this study, the MGE
PA-conferred increase in PAs contents and their gene expressions contributed to distinct defense mechanisms, which improved the tolerance of the wheat plant to DIw stress (
Figure 1 and
Figure 2,
Table 2,
Table 3,
Table 4,
Table 5,
Table 6,
Table 7,
Table 8 and
Table 9). The integrated defense mechanisms of antioxidant and osmoregulation systems along with hormonal content and gene expressions of PAs, regulated by MGE
PA not only evaluate efficacy in limiting the effects of hazardous stress but also modulate physiological status to terminate the trade-off of plant integrity related to defense responses (
Figure 1 and
Figure 2,
Table 2,
Table 3,
Table 4,
Table 5,
Table 6,
Table 7,
Table 8 and
Table 9). Therefore, wheat plants could survive better with MGE
PA pretreatment in areas with a shortage of water such as dry ones. MGE
PA pretreatment stimulated more accumulation in PAs (PUT, SPD, and SPM) that act as signaling molecules to focus primarily on metabolism in relation to conservation against DIw stress in wheat plants (
Table 7 and
Table 9). This finding, together with other cumulative zeatins and antioxidants (
Table 2,
Table 3,
Table 4,
Table 5,
Table 6,
Table 7,
Table 8 and
Table 9), is likely due to the attenuation of the adverse effects of DIw stress through its powerful antioxidant roles [
29,
67] and their gene expressions [
26]. Cumulative levels of endogenous PAs under DIw stress conditions have been linked to effective up-regulation of SPDS, ADC, DHS, SPMS, and SAMDC gene expression but not to the ODC gene (
Table 8 and
Table 9). Similar results were reported by Ebeed et al. [
67] and Alharby et al. [
26]. The results of this study indicate that PAs, PUT, SPD, and SPM were synthesized under DIw stress through the SPDS, ADC, DHS, SPMS, and SAMDC pathways (and not from ODC) in wheat plants. Various enzymes were involved in the pathway from PUT to SPM and SPD, including SAMDC, SPDS, and SPMS gene expression. The endogenous levels of SPD, SPM, and PUT that accumulated by MGE
PA in DIw-stressed wheat plants were connected to the up-regulated gene expression levels of the ADC, SPDS, SPMS, SAMDC, and DHS genes, giving wheat plants a robust antioxidant defense to withstand DIw stress (
Figure 1 and
Figure 2,
Table 2,
Table 3,
Table 4,
Table 5,
Table 6,
Table 7,
Table 8 and
Table 9).
MGE
PA efficiently outperformed MGE in promoting wheat plant growth, grain yield, physio-biochemistry, and antioxidant defense system components (
Figure 1 and
Figure 2,
Table 2,
Table 3,
Table 4,
Table 5,
Table 6,
Table 7,
Table 8 and
Table 9). These promoted results can be attributed to MGE
PA’s further enrichment of PAs along with its high antioxidants and phytohormone contents, as well as its higher antioxidant activity (88.6%) (
Table 1). This makes MGE
PA possess pivotal mechanisms for inhibiting or at least minimizing the dangerous impacts of oxidative stress, cell membrane lipid peroxidation, and ionic leakage [
25,
26]. Therefore, MGE
PA, an eco-friendly strategy, is a powerful natural extract that contains many antioxidants and biostimulants for plant growth under stress.

 ,
, 



{kind=link}
{kind=link}