
The Effects of Plant Growth-Promoting Bacteria with Biostimulant Features on the Growth of a Local Onion Cultivar and a Commercial Zucchini Variety


 ,
,  , ,
, ,  , ,
, ,  ,
, 


Abstract
1. Introduction
2. Materials and Methods
2.1. Origins of Bacterial Strains and Seeds
2.2. Assessment of PGPB Physiological Traits
2.3. Onion and Zucchini Cultivation
2.4. Mineral, Vitamin and Sulphur Compound Analysis
2.5. Water Content
2.6. Sugar Concentration
2.7. Mineral Concentration
2.8. Protein Extraction and Quantification
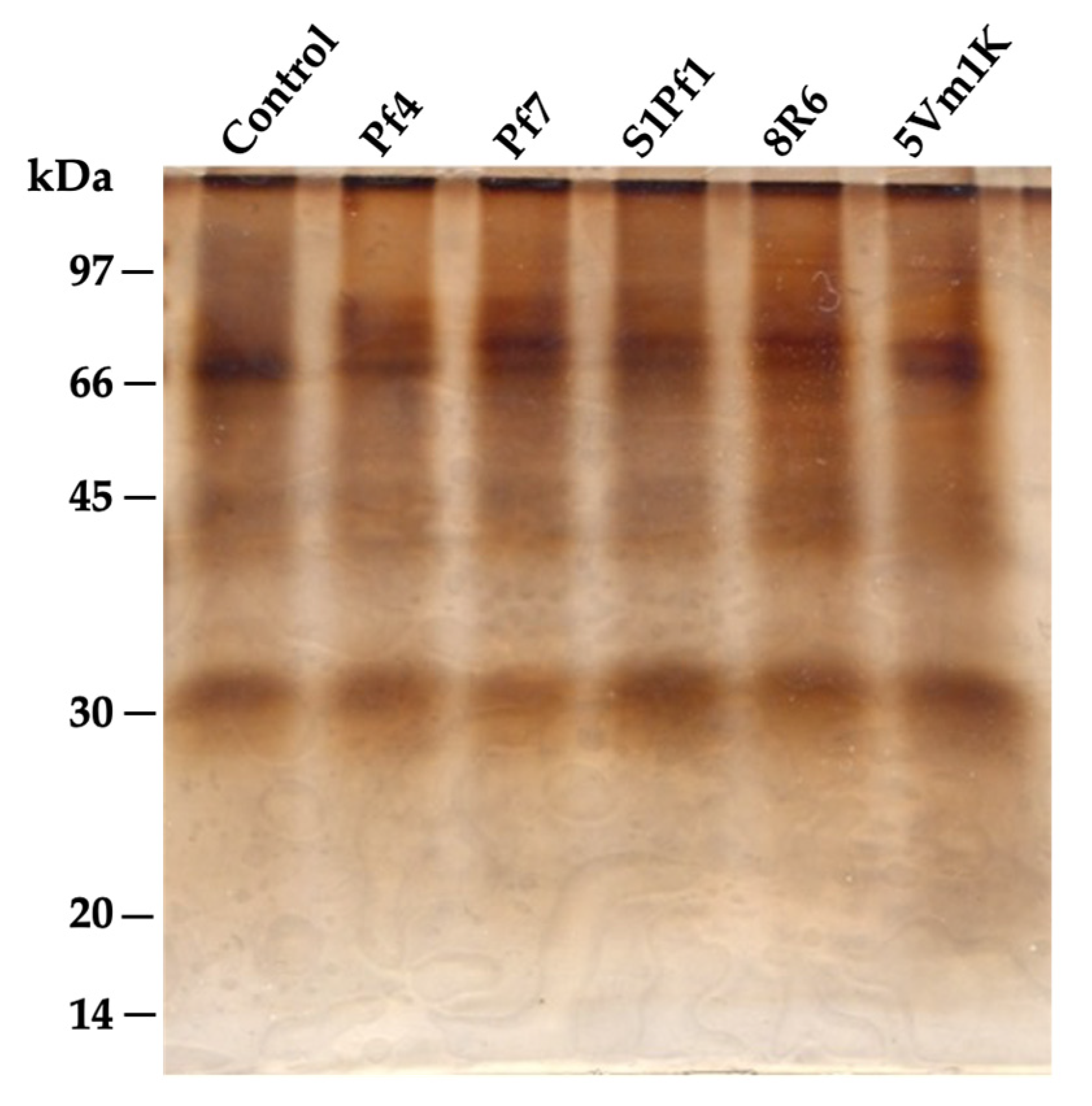
2.9. Protein Electrophoresis
2.10. Statistical Analysis
3. Results
3.1. Plant Beneficial Physiological Traits of PGPB Strains
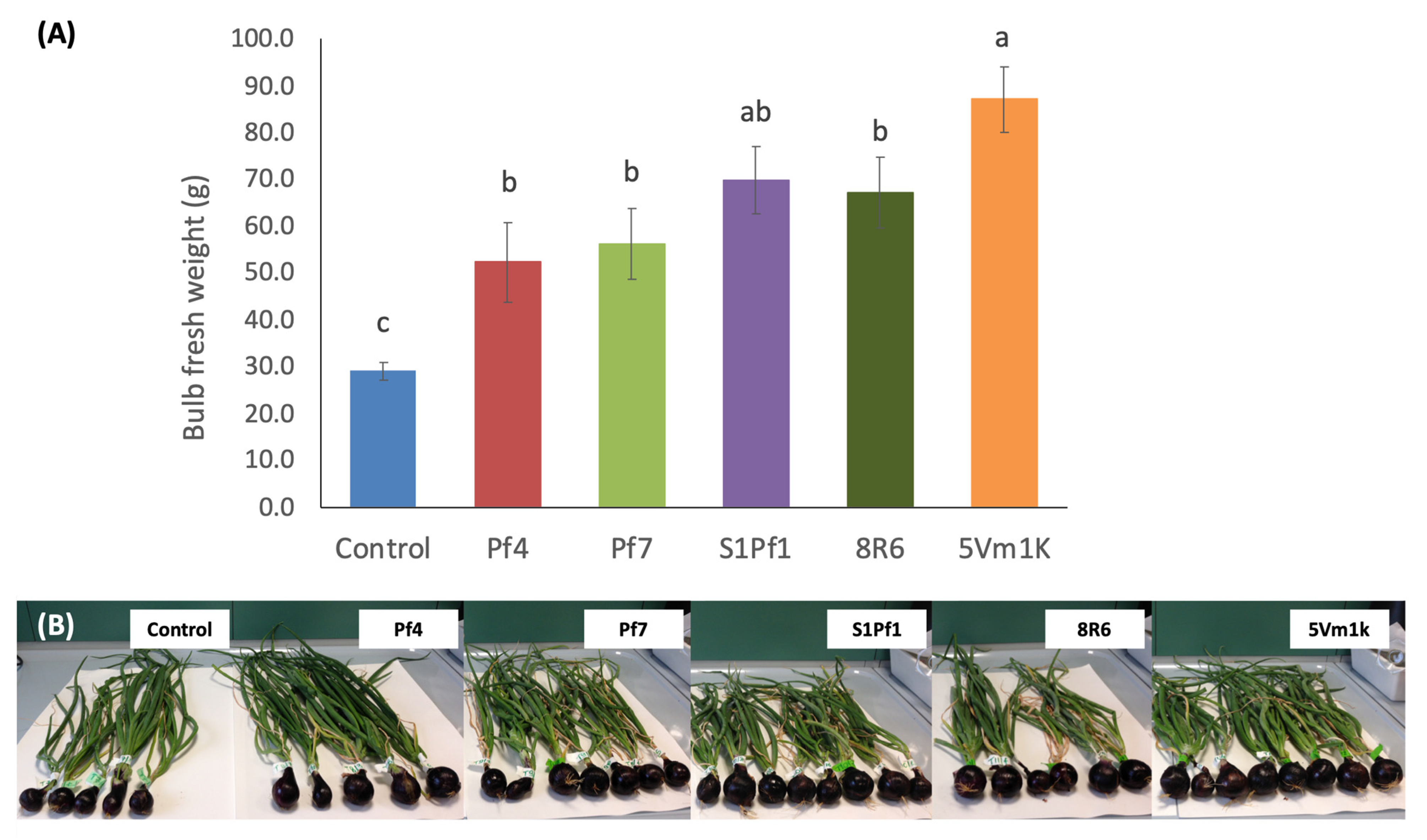
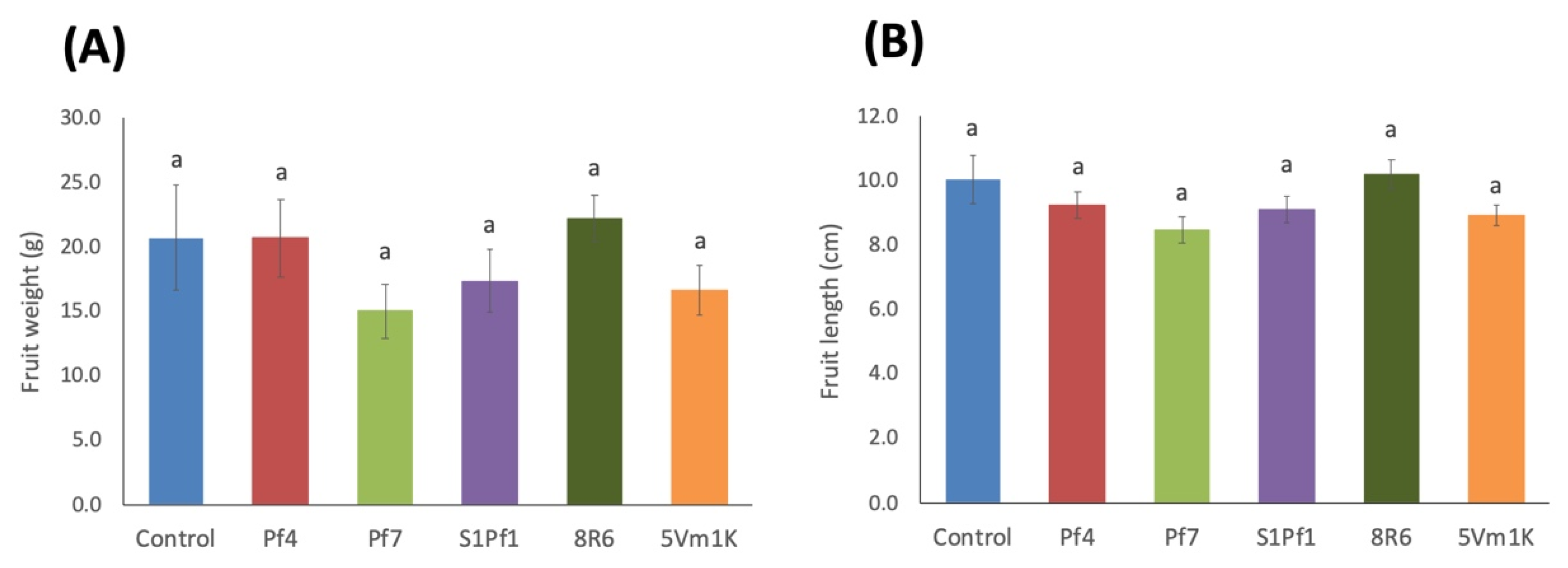
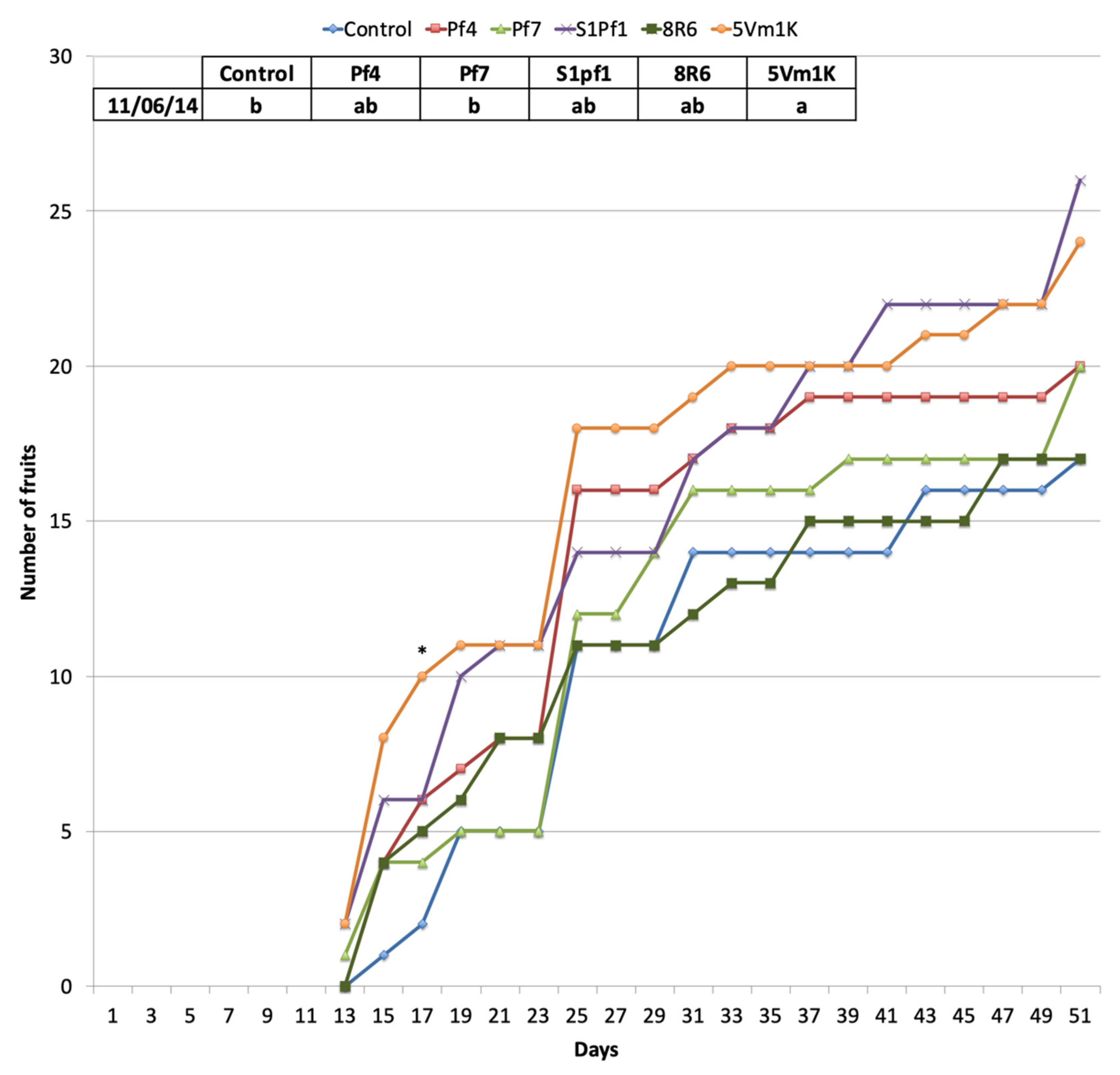
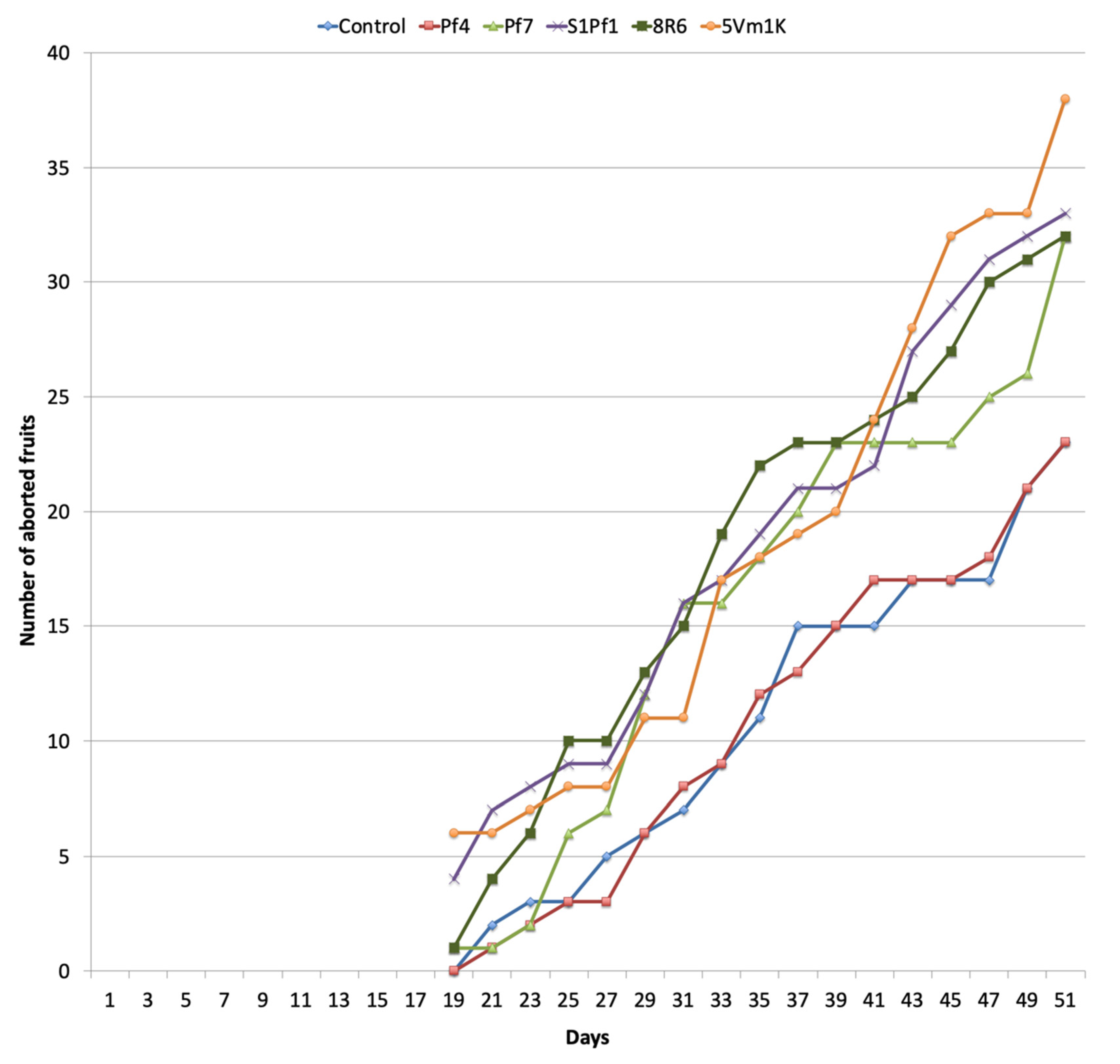
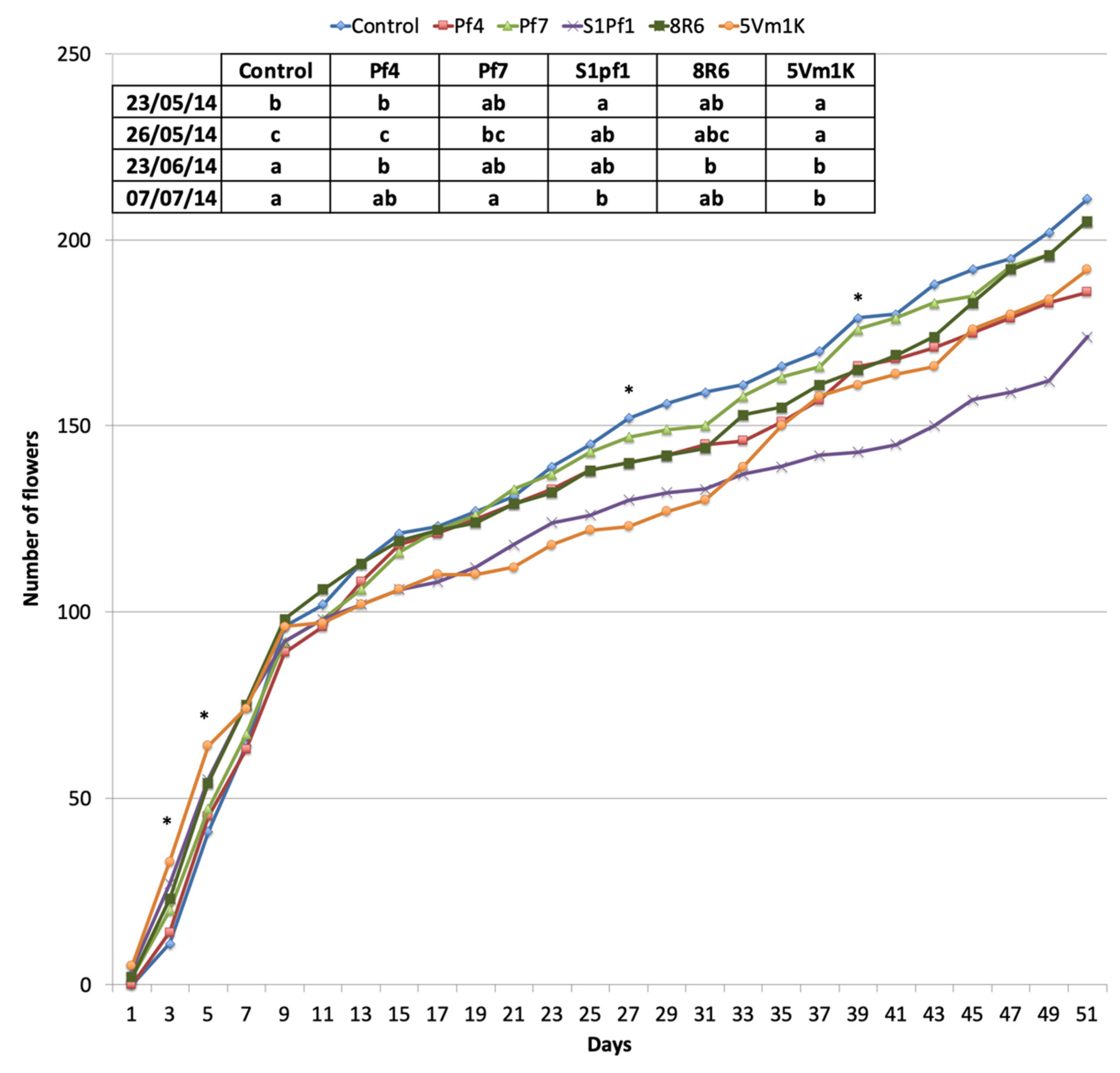
3.2. Effects of PGPBs on the Onion and Zucchini Growth and Yield
3.3. Effects of PGPBs on the Onion Quality
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Scherhaufer, S.; Moates, G.; Hartikainen, H.; Waldron, K.; Obersteiner, G. Environmental Impacts of Food Waste in Europe. Waste Manag. 2018, 77, 98–113. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Bravo, P.; Chambers, V.E.; Noguera-Artiaga, L.; Sendra, E.; Chambers IV, E.; Carbonell-Barrachina, Á.A. Consumer Understanding of Sustainability Concept in Agricultural Products. Food Qual. Prefer. 2021, 89, 104136. [Google Scholar] [CrossRef]
- Darré, E.; Cadenazzi, M.; Mazzilli, S.R.; Rosas, J.F.; Picasso, V.D. Environmental Impacts on Water Resources from Summer Crops in Rainfed and Irrigated Systems. J. Environ. Manag. 2019, 232, 514–522. [Google Scholar] [CrossRef]
- Gamalero, E.; Bona, E.; Todeschini, V.; Lingua, G. Saline and Arid Soils: Impact on Bacteria, Plants, and Their Interaction. Biology 2020, 9, 116. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Geelen, D. Developing Biostimulants from Agro-Food and Industrial By-Products. Front. Plant Sci. 2018, 9, 1567. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef]
- Del Buono, D. Can Biostimulants Be Used to Mitigate the Effect of Anthropogenic Climate Change on Agriculture? It Is Time to Respond. Sci. Total Environ. 2021, 751, 141763. [Google Scholar] [CrossRef]
- Yi, H.-S.; Yang, J.W.; Ryu, C.-M. ISR Meets SAR Outside: Additive Action of the Endophyte Bacillus pumilus INR7 and the Chemical Inducer, Benzothiadiazole, on Induced Resistance against Bacterial Spot in Field-Grown Pepper. Front. Plant Sci. 2013, 4, 122. [Google Scholar] [CrossRef]
- Vinodkumar, S.; Nakkeeran, S.; Renukadevi, P.; Mohankumar, S. Diversity and Antiviral Potential of Rhizospheric and Endophytic Bacillus Species and Phyto-Antiviral Principles against Tobacco Streak Virus in Cotton. Agric. Ecosyst. Environ. 2018, 267, 42–51. [Google Scholar] [CrossRef]
- Ali, S.; Hameed, S.; Shahid, M.; Iqbal, M.; Lazarovits, G.; Imran, A. Functional Characterization of Potential PGPR Exhibiting Broad-Spectrum Antifungal Activity. Microbiol. Res. 2020, 232, 126389. [Google Scholar] [CrossRef]
- Maxton, A.; Singh, P.; Aruna, A.; Prasad, S.M.; Masih, S.A. A Boon in Stress Tolerance and Biocontrol. Res. J. Biotechnol. 2018, 13, 105–111. [Google Scholar]
- Gamalero, E.; Glick, B.R. The Use of Plant Growth-Promoting Bacteria to Prevent Nematode Damage to Plants. Biology 2020, 9, 381. [Google Scholar] [CrossRef]
- Bona, E.; Lingua, G.; Todeschini, V. Effect of Bioinoculants on the Quality of Crops. In Bioformulations: For Sustainable Agriculture; Arora, N.K., Mehnaz, S., Balestrini, R., Eds.; Springer: New Delhi, India, 2016; pp. 93–124. ISBN 978-81-322-2777-9. [Google Scholar]
- Berta, G.; Copetta, A.; Gamalero, E.; Bona, E.; Cesaro, P.; Scarafoni, A.; D’Agostino, G. Maize Development and Grain Quality Are Differentially Affected by Mycorrhizal Fungi and a Growth-Promoting Pseudomonad in the Field. Mycorrhiza 2014, 24, 161–170. [Google Scholar] [CrossRef]
- Bona, E.; Lingua, G.; Manassero, P.; Cantamessa, S.; Marsano, F.; Todeschini, V.; Copetta, A.; D’Agostino, G.; Massa, N.; Avidano, L.; et al. AM Fungi and PGP Pseudomonads Increase Flowering, Fruit Production, and Vitamin Content in Strawberry Grown at Low Nitrogen and Phosphorus Levels. Mycorrhiza 2015, 25, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Bona, E.; Todeschini, V.; Cantamessa, S.; Cesaro, P.; Copetta, A.; Lingua, G.; Gamalero, E.; Berta, G.; Massa, N. Combined Bacterial and Mycorrhizal Inocula Improve Tomato Quality at Reduced Fertilization. Sci. Hortic. 2018, 234, 160–165. [Google Scholar] [CrossRef]
- Arıkan, Ş.; Eşitken, A.; İpek, M.; Aras, S.; Şahin, M.; Pırlak, L.; Dönmez, M.F.; Turan, M. Effect of Plant Growth Promoting Rhizobacteria on Fe Acquisition in Peach (Prunus persica L) Under Calcareous Soil Conditions. J. Plant Nutr. 2018, 41, 2141–2150. [Google Scholar] [CrossRef]
- Todeschini, V.; AitLahmidi, N.; Mazzucco, E.; Marsano, F.; Gosetti, F.; Robotti, E.; Bona, E.; Massa, N.; Bonneau, L.; Marengo, E.; et al. Impact of Beneficial Microorganisms on Strawberry Growth, Fruit Production, Nutritional Quality, and Volatilome. Front. Plant Sci. 2018, 9, 1611. [Google Scholar] [CrossRef] [PubMed]
- Massa, N.; Cesaro, P.; Todeschini, V.; Capraro, J.; Scarafoni, A.; Cantamessa, S.; Copetta, A.; Anastasia, F.; Gamalero, E.; Lingua, G.; et al. Selected Autochthonous Rhizobia, Applied in Combination with AM Fungi, Improve Seed Quality of Common Bean Cultivated in Reduced Fertilization Condition. Appl. Soil Ecol. 2020, 148, 103507. [Google Scholar] [CrossRef]
- Lingua, G.; Bona, E.; Manassero, P.; Marsano, F.; Todeschini, V.; Cantamessa, S.; Copetta, A.; D’Agostino, G.; Gamalero, E.; Berta, G. Arbuscular Mycorrhizal Fungi and Plant Growth-Promoting Pseudomonads Increases Anthocyanin Concentration in Strawberry Fruits (Fragaria x ananassa Var. Selva) in Conditions of Reduced Fertilization. IJMS 2013, 14, 16207–16225. [Google Scholar] [CrossRef]
- Aimo, S.; Gosetti, F.; D’Agostino, G.; Gamalero, E.; Gianotti, V.; Bottaro, M.; Gennaro, M.C.; Berta, G. Use of Arbuscular Mycorrhizal Fungi and Beneficial Soil Bacteria to Imrpove Yield and Quality of Saffron (Crocus sativus L.). Acta Hortic. 2010, 850, 159–164. [Google Scholar] [CrossRef]
- Lingua, G.; Copetta, A.; Musso, D.; Aimo, S.; Ranzenigo, A.; Buico, A.; Gianotti, V.; Osella, D.; Berta, G. Effect of Arbu-scular Mycorrhizal and Bacterial Inocula on Nitrate Concentration in Mesocosms Simulating a Wastewater Treatment System Relying on Phytodepuration. Environ. Sci. Pollut. Res. 2015, 22, 18616–18625. [Google Scholar] [CrossRef] [PubMed]
- D’Amelio, R.; Berta, G.; Gamalero, E.; Massa, N.; Avidano, L.; Cantamessa, S.; D’Agostino, G.; Bosco, D.; Marzachì, C. Increased Plant Tolerance against Chrysanthemum Yellows Phytoplasma (‘Candidatus Phytoplasma Asteris’) Following Double Inoculation with Glomus mosseae BEG12 and Pseudomonas putida S1Pf1Rif: Effects of AM Fungus and a Pseudomonad on Phytoplasmosis. Plant Pathol. 2011, 60, 1014–1022. [Google Scholar] [CrossRef]
- Ravera, M.; Musso, D.; Gosetti, F.; Cassino, C.; Gamalero, E.; Osella, D. Oxidative Degradation of 1,5-Naphthalenedisulfonic Acid in Aqueous Solutions by UV-Photolysis in the Absence and Presence of H2O2. Chemosphere 2010, 79, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Duan, J.; Charles, T.C.; Glick, B.R. A Bioinformatics Approach to the Determination of Genes Involved in Endophytic Behavior in Burkholderia spp. J. Theor. Biol. 2014, 343, 193–198. [Google Scholar] [CrossRef]
- Gamalero, E.; Berta, G.; Massa, N.; Glick, B.R.; Lingua, G. Interactions between Pseudomonas putida UW4 and Gigaspora rosea BEG9 and Their Consequences for the Growth of Cucumber under Salt-Stress Conditions: Salt Affects Bacteria–AM Fungi Interactions. J. Appl. Microbiol. 2010, 108, 236–245. [Google Scholar] [CrossRef]
- Penrose, D.M.; Glick, B.R. Methods for Isolating and Characterizing ACC Deaminase-Containing Plant Growth-Promo-ting Rhizobacteria. Physiol. Plant. 2003, 118, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Schwyn, B.; Neilands, J. Universal Chemical Assay for the Detection and Determination of Siderophore. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Goldstein, A.H. Bacterial Solubilization of Mineral Phosphates: Historical Perspective and Future Prospects. Am. J. Alt. Ag. 1986, 1, 51–57. [Google Scholar] [CrossRef]
- De Brito, A.M.; Gagne, S.; Antoun, H. Effect of Compost on Rhizosphere Microflora of the Tomato and on the Incidence of Plant Growth-Promoting Rhizobacteria. Appl. Environ. Microbiol. 1995, 61, 194–199. [Google Scholar] [CrossRef]
- Forni, C.; Riov, J.; Grilli Caiola, M.; Tel-Or, E. Indole-3-Acetic Acid (IAA) Production by Arthrobacter Species Isolated from Azolla. J. Gen. Microbiol. 1992, 138, 377–381. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantification of Micrograms Quantities of Proteins Utilizing the Principle of Protein-Dye Binding. Ann. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, A.; Wilm, M.; Vorm, O.; Mann, M. Mass Spectrometric Sequencing of Proteins from Silver-Stained Polyacrylamide Gels. Anal. Chem. 1996, 68, 850–858. [Google Scholar] [CrossRef] [PubMed]
- R Core R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018.
- Gamalero, E.; D’Amelio, R.; Musso, C.; Cantamessa, S.; Pivato, B.; D’Agostino, G.; Duan, J.; Bosco, D.; Marzachì, C.; Berta, G. Effects of Pseudomonas putida S1Pf1Rif Against Chrysanthemum Yellows Phytoplasma Infection. Phytopathology 2010, 100, 805–813. [Google Scholar] [CrossRef]
- Rashid, S.; Charles, T.; Glick, B. Isolation and Characterization of New Plant Growth-Promoting Bacterial Endophytes. Appl. Soil Ecol. 2012, 61, 217–224. [Google Scholar] [CrossRef]
- Bona, E.; Cantamessa, S.; Massa, N.; Manassero, P.; Marsano, F.; Copetta, A.; Lingua, G.; D’Agostino, G.; Gamalero, E.; Berta, G. Arbuscular Mycorrhizal Fungi and Plant Growth-Promoting Pseudomonads Improve Yield, Quality and Nutritional Value of Tomato: A Field Study. Mycorrhiza 2017, 27, 1–11. [Google Scholar] [CrossRef]
- Drogue, B.; Doré, H.; Borland, S.; Wisniewski-Dyé, F.; Prigent-Combaret, C. Which Specificity in Cooperation between Phytostimulating Rhizobacteria and Plants? Res. Microbiol. 2012, 163, 500–510. [Google Scholar] [CrossRef]
- Dilpreet, T.; Kulbir, S.; Varinder, S.; Jagdish, S. Growth, Yield And Quality Of Onion As Influenced By Integrated Nutrient Management. Int. J. Agric. Sci. 2016, 8, 5. [Google Scholar]
- Tinna, D.; Garg, N.; Sharma, S.; Pandove, G.; Chawla, N. Utilization of Plant Growth Promoting Rhizobacteria as Root Dipping of Seedlings for Improving Bulb Yield and Curtailing Mineral Fertilizer Use in Onion under Field Conditions. Sci. Hortic. 2020, 270, 109432. [Google Scholar] [CrossRef]
- Slimestad, R.; Fossen, T.; Vågen, I.M. Onions: A Source of Unique Dietary Flavonoids. J. Agric. Food Chem. 2007, 55, 10067–10080. [Google Scholar] [CrossRef]
- Malik, M.N. Bulb crops, onion. In Horticulture; National Book Foundation: Islamabad, Pakistan, 1994; pp. 500–501. [Google Scholar]
- Cutler, R.R.; Wilson, P. Antibacterial Activity of a New, Stable, Aqueous Extract of Allicin against Methicillin-Resistant Staphylococcus Aureus. Br. J. Biomed. Sci. 2004, 61, 71–74. [Google Scholar] [CrossRef]
- Borlinghaus, J.; Albrecht, F.; Gruhlke, M.; Nwachukwu, I.; Slusarenko, A. Allicin: Chemistry and Biological Properties. Molecules 2014, 19, 12591–12618. [Google Scholar] [CrossRef] [PubMed]
- Cannas, D.; Loi, E.; Serra, M.; Firinu, D.; Valera, P.; Zavattari, P. Relevance of Essential Trace Elements in Nutrition and Drinking Water for Human Health and Autoimmune Disease Risk. Nutrients 2020, 12, 2074. [Google Scholar] [CrossRef] [PubMed]
- Mehdi, Y.; Hornick, J.-L.; Istasse, L.; Dufrasne, I. Selenium in the Environment, Metabolism and Involvement in Body Functions. Molecules 2013, 18, 3292–3311. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Melough, M.; Vance, T.; Noh, H.; Koo, S.; Chun, O. Dietary Cadmium Intake and Sources in the US. Nutrients 2018, 11, 2. [Google Scholar] [CrossRef] [PubMed]
- Riaz, M.; Kamran, M.; Rizwan, M.; Ali, S.; Parveen, A.; Malik, Z.; Wang, X. Cadmium Uptake and Translocation: Selenium and Silicon Roles in Cd Detoxification for the Production of Low Cd Crops: A Critical Review. Chemosphere 2021, 273, 129690. [Google Scholar] [CrossRef]
- Guo, J.; Lv, X.; Jia, H.; Hua, L.; Ren, X.; Muhammad, H.; Wei, T.; Ding, Y. Effects of EDTA and Plant Growth-Promoting Rhizobacteria on Plant Growth and Heavy Metal Uptake of Hyperaccumulator Sedum alfredii Hance. J. Environ. Sci. 2020, 88, 361–369. [Google Scholar] [CrossRef]
- Danezis, G.P.; Tsagkaris, A.S.; Camin, F.; Brusic, V.; Georgiou, C.A. Food Authentication: Techniques, Trends & Emer-ging Approaches. Trac Trends Anal. Chem. 2016, 85, 123–132. [Google Scholar] [CrossRef]
- Fernandes, S.; Gois, A.; Mendes, F.; Perestrelo, R.; Medina, S.; Câmara, J.S. Typicality Assessment of Onions (Allium cepa) from Different Geographical Regions Based on the Volatile Signature and Chemometric Tools. Foods 2020, 9, 375. [Google Scholar] [CrossRef]
- Kurokura, T.; Hiraide, S.; Shimamura, Y.; Yamane, K. PGPR Improves Yield of Strawberry Species under Less-Fertilized Conditions. ECB 2017, 55, 121–128. [Google Scholar] [CrossRef]
- Bryla, D. Mycorrhizal Response of Two Tomato Genotypes Relates to Their Ability to Acquire and Utilize Phosphorus. Ann. Bot. 1998, 82, 849–857. [Google Scholar] [CrossRef]
- MacMillan, J. Occurrence of Gibberellins in Vascular Plants, Fungi, and Bacteria. J. Plant Growth Regul. 2001, 20, 387–442. [Google Scholar] [CrossRef] [PubMed]
- Gamalero, E.; Glick, B.R. Mechanisms Used by Plant Growth-Promoting Bacteria. In Bacteria in Agrobiology: Plant Nutrient Management; Maheshwari, D.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 17–46. ISBN 978-3-642-21060-0. [Google Scholar]
- Nordstedt, N.P.; Chapin, L.J.; Taylor, C.G.; Jones, M.L. Identification of Pseudomonas Spp. That Increase Ornamental Crop Quality during Abiotic Stress. Front. Plant Sci. 2020, 10, 1754. [Google Scholar] [CrossRef] [PubMed]
- Zaferanchi, S.; Salmasi, S.Z.; Lisar, S.Y.S.; Sarikhani, M.R. Bio-Inoculants and Organics Influence on Mineral Nutrition and Productivity in Calendula Officinalis L. J. Med. Plants By-prod. 2020, 1, 43–50. [Google Scholar]
- Franks, S.J. The Unique and Multifaceted Importance of the Timing of Flowering. Am. J. Bot. 2015, 102, 1401–1402. [Google Scholar] [CrossRef]
- Tabassum, B.; Khan, A.; Tariq, M.; Ramzan, M.; Iqbal Khan, M.S.; Shahid, N.; Aaliya, K. Bottlenecks in Commercialisation and Future Prospects of PGPR. Appl. Soil Ecol. 2017, 121, 102–117. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analyte | Q1 | Q3 | Dwell Time (ms) | DP (V) | EP (V) | CEP (V) | CE (V) | CXP (V) |
|---|---|---|---|---|---|---|---|---|
| Alliin | 178 | 88/74 | 25 | 14.80 | 4.0 | 15.1 | 12.1/28.4 | 2.2/2.2 |
| Vitamin C | 177 | 95/141 | 25 | 23.00 | 4.3 | 15.1 | 16.0/9.3 | 2.2/2.3 |
| Retinol | 269 | 93/91 | 25 | 38.86 | 3.5 | 17.7 | 31.4/63.0 | 2.1/2.1 |
| Strain | ACCd 1 | Siderophore 2 | Phosphate Solubilization (DCP) 3 | Phosphate Solubilization (TCP) 4 | IAA 5 | References |
|---|---|---|---|---|---|---|
| P. fluorescens Pf4 | 0.0 ± 0.0 | 3.8 ± 0.2 | 1.25 | + * | 103 ± 2 | Berta et al., 2014 [14] |
| P. protegens Pf7 | ND | 1.8 ± 0.6 * | 0.00 | + * | 39.0 ± 0.7 * | This work |
| P. putida S1Pf1 | 0.0 ± 0.0 | 5.5 ± 0.2 | 0.00 | + * | 10.6 ± 0.6 | Gamalero et al., 2010 [36] |
| P. migulae 8R6 | 10.90 | 1.9 ± 0.3 * | 0.00 | + * | 39.43 | Rashid et al., 2012 [37] |
| Pseudomonas sp. 5Vm1K | ND | 3.8 ± 0.2 | 0.9 | + * | ++++ 6 | Bona et al., 2015 [15] |
| Control | Pf4 | Pf7 | S1Pf1 | 8R6 | 5Vm1K | |
|---|---|---|---|---|---|---|
| Collar diameter A (cm) | 1.0 ± 0.2 a | 1.3 ± 0.2 ab | 1.0 ± 0.2 a | 1.5 ± 0.1 b | 1.4 ± 0.1 b | 1.6 ± 0.2 b |
| Bulb major diameter B (cm) | 3.5 ± 0.1 a | 4.4 ± 0.4 b | 4.7 ± 0.3 bc | 5.2 ± 0.2 c | 5.2 ± 0.2 c | 5.3 ± 0.2 c |
| Bulb minor diameter C (cm) | 3.3 ± 0.2 a | 4.1 ± 0.4 b | 4.4 ± 0.3 b | 5.0 ± 0.2 bc | 4.8 ± 0.2 bc | 5.5 ± 0.2 c |
| Water content (%) | 90.4 | 82.6 | 90.9 | 90.4 | 90.3 | 90.8 |
| Control | Pf4 | Pf7 | S1Pf1 | 8R6 | 5Vm1K | |
|---|---|---|---|---|---|---|
| Proteins | 1.1 ± 0.1 a | 0.9 ± 0.5 a | 1.1 ± 0.1 a | 1.5 ± 0.1 a | 1.1 ± 0.1 a | 1.2 ± 0.1 a |
| Sucrose | 5.6 ± 0.4 a | 6.0 ± 0.2 a | 5.8 ± 0.3 a | 6.3 ± 0.5 a | 6.2 ± 0.3 a | 5.9± 0.1 a |
| D-glucose | 14.3 ± 0.7 a | 12.9 ± 0.6 a | 14.8 ± 0.5 a | 13.2 ± 1.2 a | 14.4 ± 0.6 a | 12.7 ± 0.9 a |
| D-fructose | 12.5 ± 0.9 a | 12.7 ± 0.7 a | 13.9 ± 0.4 a | 14.4 ± 0.6 a | 12.6 ± 0.5 a | 13.9 ± 0.7 a |
| Alliin (μg g−1) | 1.22 ± 0.16 a | 10.13 ± 9.26 a | 9.13 ± 7.60 a | 7.07 ± 2.63 a | 0.80 ± 0.08 a | 1.07 ± 0.09 a |
| Vitamin C (μg g−1) | 3.2 ± 0.8 a | 2.3 ± 0.2 a | 2.8 ± 0.2 a | 2.9 ± 0.2 a | 2.1 ± 0.2 a | 2.9 ± 0.3 a |
| Retinol (μg g−1) | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Control | Pf4 | Pf7 | S1Pf1 | 8R6 | 5Vm1K | |
|---|---|---|---|---|---|---|
| Na (mg g−1) | 1.89 ± 0.06 a | 1.90 ± 0.24 a | 2.09 ± 0.06 a | 1.66 ± 0.05 a | 1.96 ± 0.08 a | 2.14 ± 0.13 a |
| Mg (mg g−1) | 9.04 ± 0.50 a | 9.70 ± 0.50 a | 8.41 ± 0.71 a | 10.19 ± 0.45 a | 8.87 ± 0.40 a | 9.02 ± 1.16 a |
| K (mg g−1) | 166 ± 7 a | 157 ± 6 a | 160 ± 6 a | 157 ± 5 a | 100 ± 42 a | 159 ± 6 a |
| Ca (mg g−1) | 18 ± 3 ab | 19 ± 3 ab | 12 ± 1 bc | 25 ± 3 a | 12 ± 4 bc | 8 ± 1 c |
| Mn (μg g−1) | 108 ± 7 a | 82 ± 5 b | 80 ± 4 b | 114 ± 8 a | 83 ± 1 b | 85 ± 12 b |
| Fe (μg g−1) | 114 ± 2 a | 120 ± 12 a | 118 ± 15 a | 111 ± 4 a | 97 ± 5 a | 125 ± 21 a |
| Cu (μg g−1) | 39.12 ± 1.95 a | 29.51 ± 1.89 b | 26.22 ± 1.87 bc | 25.43 ± 1.35 bc | 21.67 ± 2.45 c | 22.94 ± 0.42 c |
| Zn (μg g−1) | 261 ± 7 a | 180 ± 15 b | 131 ± 14 cd | 162 ± 13 bc | 103 ± 5 d | 114 ± 11 d |
| Se (μg g−1) | 1.15 ± 0.11 c | 1.36 ± 0.04 ab | 1.22 ± 0.06 b | 1.36 ± 0.08 ab | 1.38 ± 0.05 ab | 1.51 ± 0.03 a |
| Cd (μg g−1) | 2.04 ± 0.21 a | 1.00 ± 0.09 b | 0.79 ± 0.06 bc | 1.1 ± 0.11 b | 0.59 ± 0.02 c | 0.53 ± 0.10 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Novello, G.; Cesaro, P.; Bona, E.; Massa, N.; Gosetti, F.; Scarafoni, A.; Todeschini, V.; Berta, G.; Lingua, G.; Gamalero, E. The Effects of Plant Growth-Promoting Bacteria with Biostimulant Features on the Growth of a Local Onion Cultivar and a Commercial Zucchini Variety. Agronomy 2021, 11, 888. https://doi.org/10.3390/agronomy11050888
Novello G, Cesaro P, Bona E, Massa N, Gosetti F, Scarafoni A, Todeschini V, Berta G, Lingua G, Gamalero E. The Effects of Plant Growth-Promoting Bacteria with Biostimulant Features on the Growth of a Local Onion Cultivar and a Commercial Zucchini Variety. Agronomy. 2021; 11(5):888. https://doi.org/10.3390/agronomy11050888
Chicago/Turabian StyleNovello, Giorgia, Patrizia Cesaro, Elisa Bona, Nadia Massa, Fabio Gosetti, Alessio Scarafoni, Valeria Todeschini, Graziella Berta, Guido Lingua, and Elisa Gamalero. 2021. "The Effects of Plant Growth-Promoting Bacteria with Biostimulant Features on the Growth of a Local Onion Cultivar and a Commercial Zucchini Variety" Agronomy 11, no. 5: 888. https://doi.org/10.3390/agronomy11050888
APA StyleNovello, G., Cesaro, P., Bona, E., Massa, N., Gosetti, F., Scarafoni, A., Todeschini, V., Berta, G., Lingua, G., & Gamalero, E. (2021). The Effects of Plant Growth-Promoting Bacteria with Biostimulant Features on the Growth of a Local Onion Cultivar and a Commercial Zucchini Variety. Agronomy, 11(5), 888. https://doi.org/10.3390/agronomy11050888