
Rhizobacteria Associated with a Native Solanaceae Promote Plant Growth and Decrease the Effects of Fusariumoxysporum in Tomato




Abstract
1. Introduction
2. Materials and Methods
2.1. Rhizosphere soil Sampling and Isolation of Solanum Hindsianum Rhizospheric Bacteria
2.2. In Vitro Antifungal Activity against Fusarium Oxysporum f. sp. Lycopersici
2.3. In Vitro Screening of Plant Growth Promoting Traits
2.3.1. Phosphate Solubilization Assay
2.3.2. Potassium Solubilization Assay
2.3.3. Zinc Solubilization Assay
2.3.4. Indole Acetic Acid Production Assay
2.3.5. Hydrogen Cyanide Production Assay
2.3.6. ACC Deaminase Assay
2.3.7. Microplate Biofilm Formation Assay
2.3.8. Siderophores Production Assay
2.3.9. Chitinase Production Assay
2.4. Effect of Rhizobacteria Inoculation on Tomato Growth
Effect of Rhizobacteria Inoculation on Tomato Fusarium Wilt
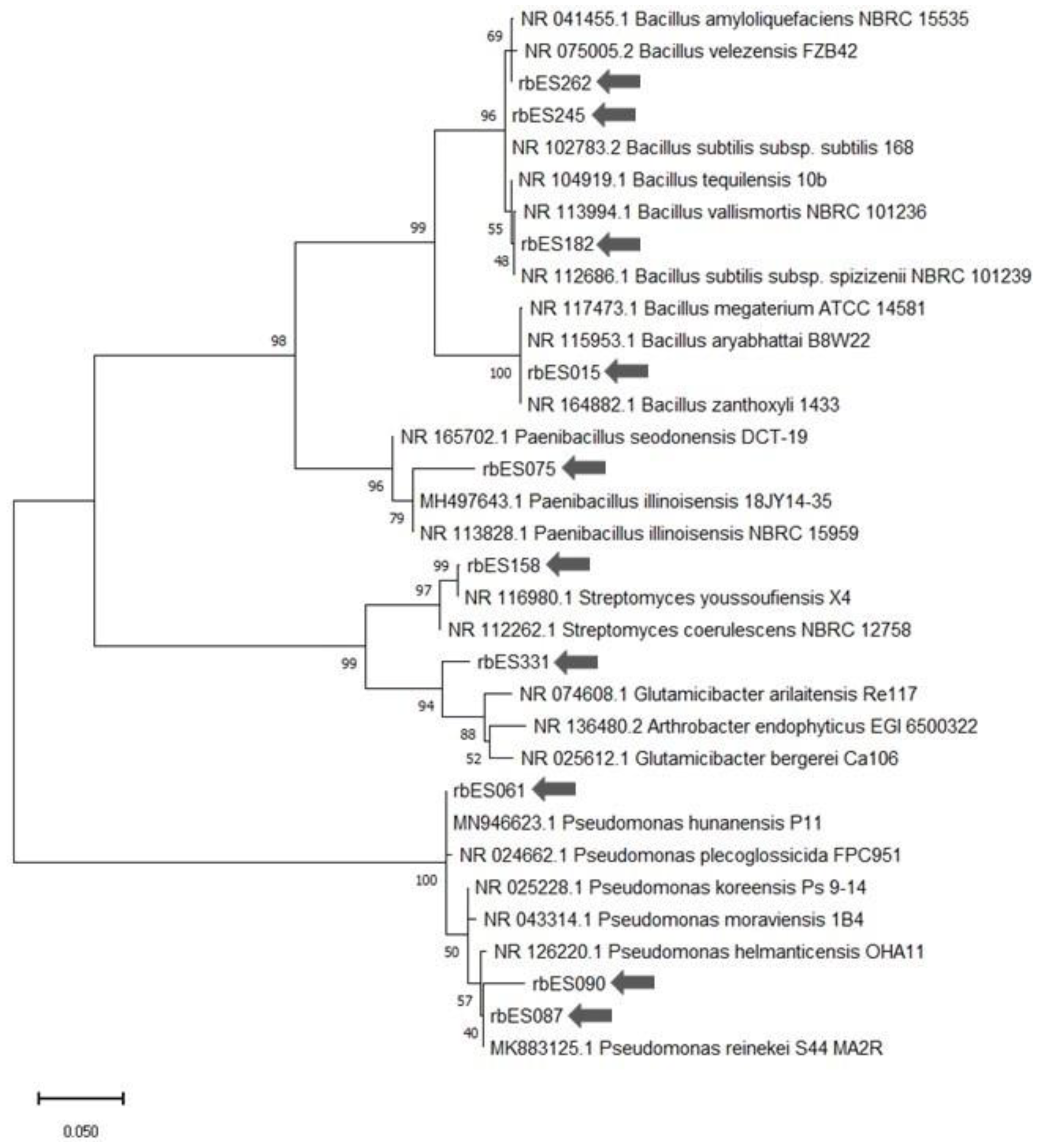
2.5. Molecular Identification of Bacterial Strains
3. Results
3.1. Antagonistic Effect of Mariola Rhizobacteria against F. oxysporum In Vitro
3.2. In Vitro Identification of Mariola Associated Rhizobacteria with Plant Growth-Promoting Traits
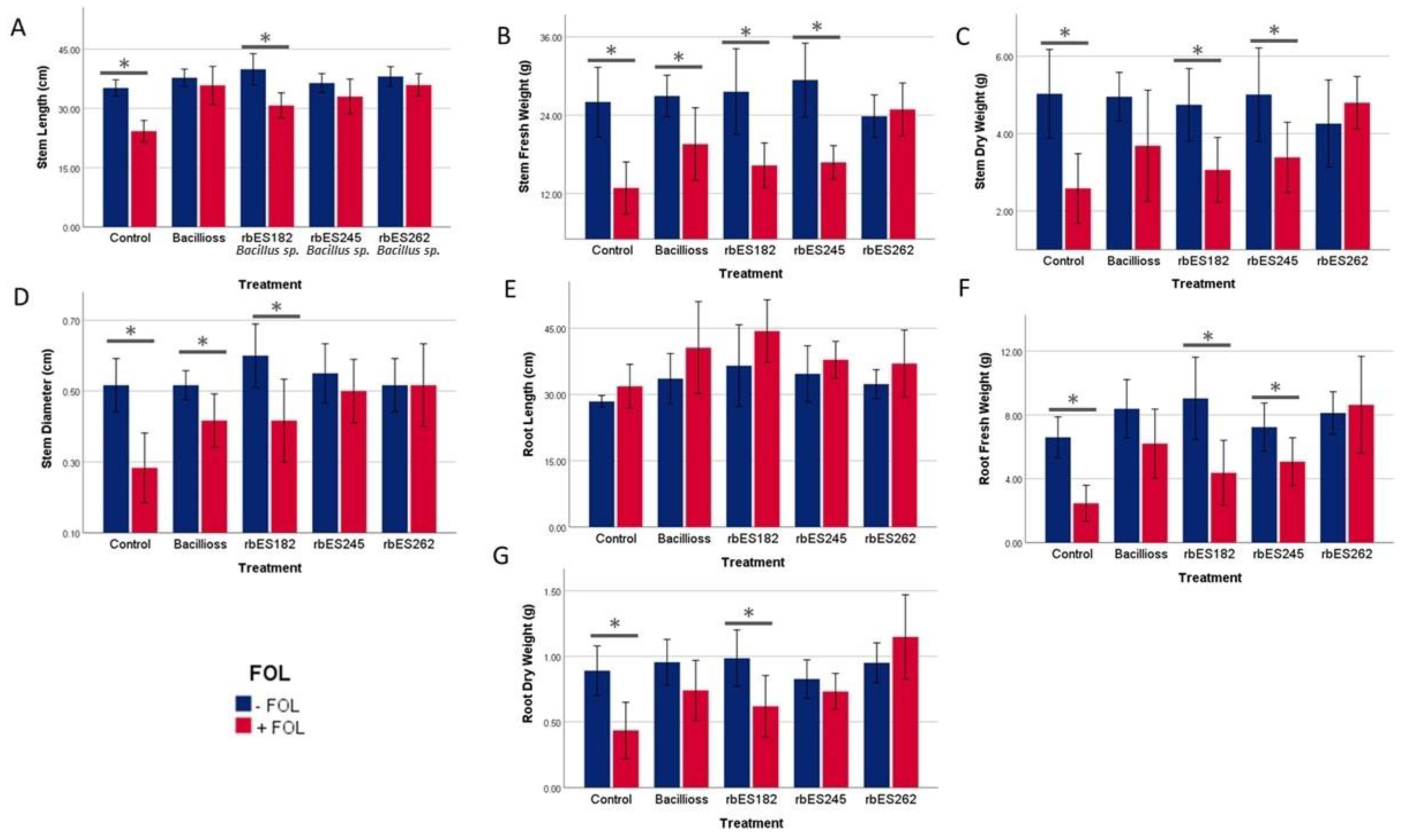
3.3. Inoculation of Tomato with Mariola Associated Rhizobacteria Increases Plant Root and Aerial Biomass
3.4. Mariola Associated Rhizobacteria Decrease the Effects of Fusarium on Tomato Growth
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Hassani, M.A.; Durán, P.; Hacquard, S. Microbial interactions within the plant holobiont. Microbiome 2018, 6, 58. [Google Scholar] [CrossRef]
- Méndez-Bravo, A.; Cortazar-Murillo, E.M.; Guevara-Avendaño, E.; Ceballos-Luna, O.; Rodríguez-Haas, B.; Kiel-Martínez, A.L.; Hernández-Cristóbal, O.; Guerrero-Analco, J.A.; Reverchon, F. Plant growth-promoting rhizobacteria associated with avocado display antagonistic activity against Phytophthora cinnamomi through volatile emissions. PLoS ONE 2018, 13, e0194665. [Google Scholar] [CrossRef] [PubMed]
- Avis, T.J.; Gravel, V.; Antoun, H.; Tweddell, R.J. Multifaceted beneficial effects of rhizosphere microorganisms on plant health and productivity. Soil Biol. Biochem. 2008, 40, 1733–1740. [Google Scholar] [CrossRef]
- Hayat, R.; Ali, S.; Amara, U.; Khalid, R.; Ahmed, I. Soil beneficial bacteria and their role in plant growth promotion: A review. Ann. Microbiol. 2010, 60, 579–598. [Google Scholar] [CrossRef]
- Khatoon, Z.; Huang, S.; Rafique, M.; Fakhar, A.; Kamran, M.A.; Santoyo, G. Unlocking the potential of plant growth-promoting rhizobacteria on soil health and the sustainability of agricultural systems. J. Environ. Manag. 2020, 273, 111118. [Google Scholar] [CrossRef]
- Berg, G. Plant-microbe interactions promoting plant growth and health: Perspectives for controlled use of microorganisms in agriculture. Appl. Microbiol. Biotechnol. 2009, 84, 11–18. [Google Scholar] [CrossRef]
- Glick, B.R. Plant Growth-Promoting Bacteria: Mechanisms and Applications. Scientifica 2012, 2012, 963401. [Google Scholar] [CrossRef]
- Collinge, D.B.; Jørgensen, H.J.; Latz, M.A.; Manzotti, A.; Ntana, F.; Rojas Tayo, E.C.; Jensen, B. Searching for novel fungal biological control agents for plant disease control among endophytes. In Endophytes for a Growing World, 1st ed.; Hodkinson, T., Doohan, F., Saunders, M., Murphy, B., Eds.; Cambridge University Press: Cambridge, UK, 2019; pp. 25–51. [Google Scholar]
- Raaijmakers, J.M.; Paulitz, T.C.; Steinberg, C.; Alabouvette, C.; Moënne-Loccoz, Y. The Rhizosphere: A Playground and Battlefield for Soilborne Pathogens and Beneficial Microorganisms. Plant Soil 2009, 321, 341–361. [Google Scholar] [CrossRef]
- Dutta, S.; Rani, T.S.; Podile, A.R. Root exudate-induced alterations in Bacillus cereus cell wall contribute to root colonization and plant growth promotion. PLoS ONE 2013, 8, e78369. [Google Scholar] [CrossRef]
- Martínez-Viveros, O.; Jorquera, M.A.; Crowley, D.E.; Gajardo, G.; Mora, M.L. Mechanisms and practical considerations involved in plant growth promotion by Rhizobacteria. J. Plant Nutr. 2010, 10, 293–319. [Google Scholar] [CrossRef]
- Requena, N.; Jimenez, I.; Toro, M.; Barea, J.M. Interactions between plant-growth-promoting rhizobacteria (PGPR), arbuscular mycorrhizal fungi and Rhizobium spp. in the rhizosphere of Anthyllis cytisoides, a model legume for revegetation in mediterranean semi-arid ecosystems. New Phytol. 1997, 136, 667–677. [Google Scholar] [CrossRef]
- Zahid, M.; Kaleem Abbasi, M.; Hameed, S.; Rahim, N. Isolation and identification of indigenous plant growth promoting rhizobacteria from Himalayan region of Kashmir and their effect on improving growth and nutrient contents of maize (Zea mays L.). Front. Microbiol. 2015, 6, 207. [Google Scholar] [CrossRef] [PubMed]
- Borja-Bravo, M.; García-Salazar, J.A.; Skaggs, R.K. Mexican fresh tomato exports in the North American market: A case study of the effects of productivity on competitiveness. Can. J. Plant Sci. 2013, 93, 839–850. [Google Scholar] [CrossRef][Green Version]
- Singh, V.K.; Singh, H.B.; Upadhyay, R.S. Role of fusaric acid in the development of “Fusarium wilt” symptoms in tomato: Physiological, biochemical and proteomic perspectives. Plant Physiol. Bioch. 2017, 118, 320–332. [Google Scholar] [CrossRef]
- Blancard, D. Tomato Diseases, 2nd ed.; CRC Press: London, UK, 2013; p. 668. [Google Scholar] [CrossRef]
- Padilla-Bernal, L.E.; Lara-Herrera, A.; Reyes-Rivas, E.; González-Hernández, J. Roberto Assessing Environmental Management of Tomato Production under Protected Agriculture. Int. Food Agribus. Manag. Rev. 2015, 18, 193–210. [Google Scholar] [CrossRef]
- Arellano, E.; Montaño-Soto, T.; Arellano-García, E.; Ojinaga, L.C.; Vonglascoe, C.; Ruiz-Ruiz, B. Genotoxic Biomonitoring and Exposure to Pesticides in Women Laborers at Maneadero Valley in Baja California, Mexico. Int. J. Appl. Nat. Sci. 2014, 3, 89–96. [Google Scholar]
- Espejel, I.; Fischer, D.W.; Hinojosa, A.; García, C.; Leyva, C. Land-use planning for the Guadalupe Valley, Baja California, Mexico. Landsc. Urban Plan. 1999, 45, 219–232. [Google Scholar] [CrossRef]
- Gilabert-Alarcón, C.; Daesslé, L.W.; Salgado-Méndez, S.O.; Pérez-Flores, M.A.; Knöller, K.; Kretzschmar, T.G.; Stumpp, C. Effects of reclaimed water discharge in the Maneadero coastal aquifer, Baja California, Mexico. J. Appl. Geochem. 2018, 92, 121–139. [Google Scholar] [CrossRef]
- Hartmann, A.; Schmid, M.; Van Tuinen, D.; Berg, G. Plant-driven selection of microbes. Plant Soil 2009, 321, 235–257. [Google Scholar] [CrossRef]
- Zhang, N.; Wang, D.; Liu, Y.; Li, S.; Shen, Q.; Zhang, R. Effects of different plant root exudates and their organic acid components on chemotaxis, biofilm formation and colonization by beneficial rhizosphere-associated bacterial strains. Plant Soil 2014, 374, 689–700. [Google Scholar] [CrossRef]
- Santoyo, G.; Hernández-Pacheco, C.; Hernández-Salmerón, J.; Hernández-León, R. The role of abiotic factors modulating the plant-microbe-soil interactions: Toward sustainable agriculture. A review. Span. J. Agric. Res. 2017, 15, 13. [Google Scholar] [CrossRef]
- Rueda-Puente, E.O.; Castellanos-Cervantes, T.; Díaz de León-Álvarez, J.L.; Preciado-Rangel, P.; Almaguer-Vargas, G. Bacterial community of rhizosphere associated to the annual halophite Salicornia bigelovii (Torr). Terra Latinoam. 2010, 28, 345–353. [Google Scholar]
- Aguirre-Garrido, J.F.; Montiel-Lug, D.; Hernández-Rodríguez, C.; Torres-Cortés, G.; Millán, V.; Toro, N.; Martínez-Abarca, F.; Ramírez-Saad, H. Bacterial community structure in the rhizosphere of three cactus species from semi-arid highlands in central Mexico. Antonie Leeuwenhoek 2012, 101, 891–904. [Google Scholar] [CrossRef] [PubMed]
- Aydi Ben Abdallah, R.; Jabnoun-Khiareddine, H.; Nefzi, A.; Mokni-Tlili, S.; Daami-Remadi, M. Biocontrol of Fusarium Wilt and Growth Promotion of Tomato Plants Using Endophytic Bacteria Isolated from Solanum elaeagnifolium Stems. J. Phytopathol. 2016, 164, 811–824. [Google Scholar] [CrossRef]
- Knapp, S.; Sagona, E.; Carbonell, A.K.Z.; Chiarini, F. A revision of the Solanum elaeagnifolium clade (Elaeagnifolium clade; subgenus Leptostemonum, Solanaceae). PhytoKeys 2017, 2017, 1–104. [Google Scholar] [CrossRef] [PubMed]
- Rebman, J.; ROberts, N.C. Baja California Plant Field Guide, 3rd ed.; San Diego Natural History Museum: San Diego, CA, USA, 2012; p. 480. ISBN 9780916251185. [Google Scholar]
- Noel, K.D.; Sanchez, A.; Fernandez, L.; Leemans, J.; Cevallos, M.A. Rhizobium phaseoli symbiotic mutants with transposon Tn5 insertions. J. Bacteriol. 1984, 158, 148–155. [Google Scholar] [CrossRef]
- Guevara-Avendaño, E.; Carrillo, J.D.; Ndinga-Muniania, C.; Moreno, K.; Méndez-Bravo, A.; Guerrero-Analco, J.A.; Eskalen, A.; Reverchon, F. Antifungal activity of avocado rhizobacteria against Fusarium euwallaceae and Graphium spp., associated with Euwallacea spp. nr. fornicatus, and Phytophthora cinnamomi. Antonie Leeuwenhoek 2018, 111, 563–572. [Google Scholar] [CrossRef]
- Gupta, R.; Singal, R.; Shankar, A.; Kuhad, R.C.; Saxena, R.K. A modified plate assay for screening phosphate solubilizing microorganisms. J. Gen. Appl. Microbiol. 1994, 40, 255–260. [Google Scholar] [CrossRef]
- Zheng, B.X.; Ibrahim, M.; Zhang, D.P.; Bi, Q.F.; Li, H.Z.; Zhou, G.W.; Ding, K.; Peñuelas, J.; Zhu, Y.G.; Yang, X.R. Identification and characterization of inorganic-phosphate-solubilizing bacteria from agricultural fields with a rapid isolation method. AMB Express 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Pikovskaya, R.I. Mobilization of Phosphorus in Soil Connection with the Vital Activity of Some Microbial Species. Microbiologiya 1948, 17, 362–370. [Google Scholar]
- Bapiri, A.; Asgharzadeh, A.; Mujallali, H.; Khavazi, K.; Pazira, E. Evaluation of Zinc solubilization potential by different strains of Fluorescent Pseudomonads. J. Appl. Sci. Environ. Manag. 2012, 16, 295–298. [Google Scholar]
- Ahmad, F.; Ahmad, I.; Khan, M.S. Screening of free-living rhizospheric bacteria for their multiple plant growth promoting activities. Microbiol. Res. 2008, 163, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Penrose, D.M.; Glick, B.R. Methods for isolating and characterizing ACC deaminase-containing plant growth-promoting rhizobacteria. Physiol. Plant 2003, 118, 10–15. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, G.A.; Pratt, L.A.; Watnick, P.I.; Newman, D.K.; Weaver, V.B.; Kolter, R. Genetic approaches to study of biofilms. Meth. Enzymol. 1999, 310, 91–109. [Google Scholar] [CrossRef]
- Hamon, M.A.; Lazazzera, B.A. The sporulation transcription factor Spo0A is required for biofilm development in Bacillus subtilis. Mol. Microbiol. 2001, 42, 1199–1209. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Lynne, A.M.; Haarmann, D.; Louden, B.C. Use of Blue Agar CAS Assay for Siderophore Detection. J. Microbiol. Biol. Edu. 2011, 12, 51–53. [Google Scholar] [CrossRef]
- Agrawal, T.; Kotasthane, A.S. Chitinolytic assay of indigenous Trichoderma isolates collected from different geographical locations of Chhattisgarh in central India. SpringerPlus 2012, 1, 1–10. [Google Scholar] [CrossRef]
- Hanson, P.; Lu, S.F.; Wang, J.F.; Chen, W.; Kenyon, L.; Tan, C.W. Conventional and molecular marker-assisted selection and pyramiding of genes for multiple disease resistance in tomato. Sci. Hortic. 2016, 201, 346–354. [Google Scholar] [CrossRef]
- Frank, J.A.; Reich, C.I.; Sharma, S.; Weisbaum, J.S.; Wilson, B.A.; Olsen, G.J. Critical evaluation of two primers commonly used for amplification of bacterial 16S rRNA genes. Appl. Environ. Microbiol. 2008, 74, 2461–2470. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. R Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Habibi, S.; Djedidi, S.; Prongjunthuek, K.; Mortuza, M.F.; Ohkama-Ohtsu, N.; Sekimoto, H.; Yokoyoma, T. Physiological and genetic characterization of rice nitrogen fixer PGPR isolated from rhizosphere soils of different crops. Plant Soil 2014, 379, 51–66. [Google Scholar] [CrossRef]
- Liu, D.; Yang, Q.; Ge, K.; Hu, X.; Qi, G.; Du, B.; Liu, K.; Ding, Y. Promotion of iron nutrition and growth on peanut by Paenibacillus illinoisensis and Bacillus sp. strains in calcareous soil. Braz. J. Microbiol. 2017, 48, 656–670. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.-G.; Mun, B.-G.; Kang, S.-M.; Hussain, A.; Shahzad, R.; Seo, C.-W.; Kim, A.-Y.; Lee, S.-U.; Oh, K.Y.; Lee, D.Y.; et al. Bacillus aryabhattai SRB02 tolerates oxidative and nitrosative stress and promotes the growth of soybean by modulating the production of phytohormones. PLoS ONE 2017, 12, e0173203. [Google Scholar] [CrossRef]
- Kang, S.M.; Khan, A.L.; Waqas, M.; Asaf, S.; Lee, K.E.; Park, Y.G.; Kim, A.Y.; Khan, M.A.; You, Y.H.; Lee, I.J. Integrated phytohormone production by the plant growth-promoting rhizobacterium Bacillus tequilensis SSB07 induced thermotolerance in soybean. J. Plant Interact. 2019, 14, 416–423. [Google Scholar] [CrossRef]
- Barraza, A.; Caamal-Chan, M.G.; Castellanos, T.; Loera-Muro, A. Bacterial community characterization of the rhizobiome of plants belonging to Solanaceae family cultivated in desert soils. Ann. Microbiol. 2020, 70, 34. [Google Scholar] [CrossRef]
- Hernández-León, R.; Rojas-Solís, D.; Contreras-Pérez, M.; Orozco-Mosqueda, M.C.; Macías-Rodríguez, L.I.; Cruz, H.R.-D.L.; Valencia-Cantero, E.; Santoyo, G. Characterization of the antifungal and plant growth-promoting effects of diffusible and volatile organic compounds produced by Pseudomonas fluorescens strains. Biol. Contr. 2015, 81, 83–92. [Google Scholar] [CrossRef]
- Schlatter, D.; Kinkel, L.; Thomashow, L.; Weller, D.; Paulitz, T. Disease Suppressive Soils: New Insights from the Soil Microbiome. Phytopathology 2017, 107, 1284–1297. [Google Scholar] [CrossRef]
- Ongena, M.; Jacques, P. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol. Trends Microbiol. 2008, 16, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Asaka, O.; Shoda, M. Biocontrol of Rhizoctonia solani Damping-Off of Tomato with Bacillus subtilis RB14. Appl. Environ. Microbiol. 1996, 62, 4081–4085. [Google Scholar] [CrossRef] [PubMed]
- Spaepen, S.; Vanderleyden, J.; Remans, R. Indole-3-acetic acid in microbial and microorganism-plant signaling. FEMS Microbiol. Rev. 2007, 31, 425–448. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.F.; Wei, J.Y.; Chen, H.W.; Liu, Y.Y.; Lu, H.Y.; Chou, J.Y. Indole-3-acetic acid: A widespread physiological code in interactions of fungi with other organisms. Plant. Signal. Behav. 2015, 10, e1048052. [Google Scholar] [CrossRef] [PubMed]
- Smyth, E.M.; McCarthy, J.; Nevin, R.; Khan, M.R.; Dow, J.M.; O’Gara, F.; Doohan, F.M. In vitro analyses are not reliable predictors of the plant growth promotion capability of bacteria; a Pseudomonas fluorescens strain that promotes the growth and yield of wheat. J. Appl. Microbiol. 2011, 111, 683–692. [Google Scholar] [CrossRef]
- Ghyselinck, J.; Velivelli, S.L.; Heylen, K.; O’Herlihy, E.; Franco, J.; Rojas, M.; Prestwich, B.D. Bioprospecting in potato fields in the Central Andean Highlands: Screening of rhizobacteria for plant growth-promoting properties. Syst. Appl. Microbiol. 2013, 36, 116–127. [Google Scholar] [CrossRef]
- Kearl, J.; McNary, C.; Lowman, J.S.; Mei, C.; Aanderud, Z.T.; Smith, S.T.; West, J.; Colton, E.; Hamson, M.; Nielsen, B.L. Salt-Tolerant Halophyte Rhizosphere Bacteria Stimulate Growth of Alfalfa in Salty Soil. Front. Microbiol. 2019, 10, 1849. [Google Scholar] [CrossRef]
- Sezen, A.; Ozdal, M.; Koc, K.; Algur, O.F. Isolation and Characterization of Plant Growth Promoting Rhizobacteria (PGPR) and Their Effects on Improving Growth of Wheat. J. Appl. Biol. Sci. 2019, 10, 41–46. Available online: http://jabsonline.org/index.php/jabs/article/view/485 (accessed on 17 March 2021).
- Aydi-Ben-Abdallah, R.; Jabnoun-Khiareddine, H.; Daami-Remadi, M. Fusarium wilt biocontrol and tomato growth stimulation, using endophytic bacteria naturally associated with Solanum sodomaeum and S. bonariense plants. Egypt. J. Biol. Pest Cont. 2020, 30, 113. [Google Scholar] [CrossRef]
- Akram, M.S.; Shahid, M.; Tariq, M.; Azeem, M.; Javed, M.T.; Saleem, S.; Riaz, S. Deciphering Staphylococcus sciuri SAT-17 Mediated Anti-oxidative Defense Mechanisms and Growth Modulations in Salt Stressed Maize (Zea mays L.). Front. Microbiol. 2016, 7, 867. [Google Scholar] [CrossRef]
- Liqiao, S.; Zhaoyuan, W.; Yani, Z.; Zhigang, Z.; Wei, F.; Yueying, W.; Zhongyi, W.; Kaimei, W.; Shaoyong, K. Herbicidal Secondary Metabolites from Actinomycetes: Structure Diversity, Modes of Action, and Their Roles in the Development of Herbicides. J. Agric. Food Chem. 2020, 68, 17–32. [Google Scholar] [CrossRef]
- Yoshida, H.; Arai, N.; Sugoh, M.; Iwabuchi, J.; Shiomi, K.; Shinose, M.; Tanaka, Y.; Omura, S. 4-chlorothreonine, a herbicidal antimetabolite produced by Streptomyces sp. OH-5093. J. Antibiot. 1994, 47, 1165–1166. [Google Scholar] [CrossRef][Green Version]
- Duke, S.O.; Dayan, F.E. Modes of Action of Microbially-Produced Phytotoxins. Toxins 2011, 3, 1038–1064. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Sugoh, M.; Ji, W.; Iwabuchi, J.; Yoshida, H.; Omura, S. Screening method for cellulose biosynthesis inhibitors with herbicidal activity. J. Antibiot. 1995, 48, 720–724. [Google Scholar] [CrossRef] [PubMed][Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PGP Associated Traits | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Activity | Strain | Sample | Genera | GeneBank | IAA (µg/mL) | K sol. | P sol. | Zn sol. | Chit | Sid | HCN | Bio-film | ACCd | Path. Inhi. | Inhi. Type |
| PGPT | rbES001 | M1 | Staphylococcus | + | + | + | ND | − | + | − | ND | ND | − | ||
| PGPT | rbES005 | M4 | Staphylococcus | + | + | + | ND | − | − | − | ND | ND | − | − | |
| PGPT | rbES015 | M1 | Bacillus | MW391472 | 3.29 | + | + | − | − | + | − | + | − | − | − |
| PGPT | rbES031 | M4 | Microbacterium | 14.54 | + | + | − | − | + | − | − | − | − | − | |
| PGPT | rbES041 | M6 | Pseudomonas | 52.87 | + | − | − | − | + | − | − | − | − | − | |
| PGPT | rbES050 | M7 | Staphylococcus | + | + | + | ND | + | + | − | ND | ND | − | − | |
| PGPT | rbES053 | M7 | Staphylococcus | + | + | + | ND | + | + | − | ND | ND | − | − | |
| PGPT | rbES057 | M7 | Staphylococcus | + | + | + | ND | − | + | − | ND | ND | − | − | |
| PGPT | rbES061 | M2 | Pseudomonas | MW391473 | 12.09 | + | + | + | − | + | − | − | − | − | − |
| PGPT | rbES062 | M2 | Bacillus | 9.57 | + | + | − | − | + | − | + | − | − | − | |
| PGPT | rbES075 | M4 | Paenibacillus | MW391474 | 16.08 | + | + | − | − | + | − | − | − | − | − |
| PGPT | rbES086 | M5 | Pseudomonas | 54.66 | + | + | − | − | + | − | − | − | − | − | |
| PGPT | rbES087 | M5 | Pseudomonas | MW391476 | 32.57 | + | + | + | − | + | − | + | − | − | − |
| PGPT | rbES090 | M5 | Pseudomonas | MW391477 | 56.12 | + | + | − | + | + | − | − | + | − | − |
| PGPT | rbES098 | M6 | Pseudomonas | 72.71 | + | + | − | + | − | − | + | − | − | − | |
| PGPT | rbES120 | M7 | Lysinibacillus | 4.52 | + | + | − | − | − | − | − | − | − | − | |
| PGPT | rbES145 | M10 | Chryseobacterium | 18.38 | + | + | + | + | − | − | − | − | − | − | |
| AA | rbES158 | M4 | Streptomyces | MW391478 | − | + | + | − | − | + | − | − | − | 24% | SS |
| PGPT | rbES169 | M1 | Staphylococcus | + | + | + | ND | − | + | − | ND | ND | − | − | |
| PPGT+AA | rbES182 | M9 | Bacillus | MW391475 | 6.55 | + | + | + | − | + | − | + | + | 58% | MS |
| AA | rbES232 | M4 | Bacillus | 11.55 | + | − | + | − | − | − | + | + | 66% | MS | |
| AA | rbES245 | M11 | Bacillus | MW391479 | 8.81 | + | + | − | − | − | − | + | − | 69% | MS |
| AA | rbES254 | M11 | Bacillus | 10.37 | + | − | − | − | − | − | + | − | 57% | MS | |
| AA | rbES256 | M11 | Bacillus | 8.24 | + | + | − | − | − | − | + | − | 55% | MS | |
| AA | rbES262 | M12 | Bacillus | MW391480 | 11.12 | − | − | − | − | − | − | + | − | 68% | MS |
| AA | rbES331 | M11 | Glutamicibacter | MW391481 | 8.6 | − | − | − | − | − | − | − | − | 23% | SS |
| Treatment | Symptoms | |||
|---|---|---|---|---|
| Leaf Clorosis | Root Necrosis | Crown Necrosis | Plant Death | |
| −Control | − | − | − | − |
| rbES158 | − | − | − | + |
| rbES182 | − | − | − | − |
| rbES245 | − | − | − | − |
| rbES262 | − | − | − | − |
| rbES331 | − | − | − | + |
| Bacillioss | − | − | − | − |
| −Control + FOL | + | + | + | − |
| rbES158 + FOL | − | + | − | + |
| rbES182 + FOL | − | − | + | − |
| rbES245 + FOL | − | + | − | − |
| rbES262 + FOL | − | − | − | − |
| rbES331 + FOL | − | + | − | + |
| Bacillioss + FOL | − | + | − | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delgado-Ramírez, C.S.; Hernández-Martínez, R.; Sepúlveda, E. Rhizobacteria Associated with a Native Solanaceae Promote Plant Growth and Decrease the Effects of Fusariumoxysporum in Tomato. Agronomy 2021, 11, 579. https://doi.org/10.3390/agronomy11030579
Delgado-Ramírez CS, Hernández-Martínez R, Sepúlveda E. Rhizobacteria Associated with a Native Solanaceae Promote Plant Growth and Decrease the Effects of Fusariumoxysporum in Tomato. Agronomy. 2021; 11(3):579. https://doi.org/10.3390/agronomy11030579
Chicago/Turabian StyleDelgado-Ramírez, Carmen Sanjuana, Rufina Hernández-Martínez, and Edgardo Sepúlveda. 2021. "Rhizobacteria Associated with a Native Solanaceae Promote Plant Growth and Decrease the Effects of Fusariumoxysporum in Tomato" Agronomy 11, no. 3: 579. https://doi.org/10.3390/agronomy11030579
APA StyleDelgado-Ramírez, C. S., Hernández-Martínez, R., & Sepúlveda, E. (2021). Rhizobacteria Associated with a Native Solanaceae Promote Plant Growth and Decrease the Effects of Fusariumoxysporum in Tomato. Agronomy, 11(3), 579. https://doi.org/10.3390/agronomy11030579