
Diversity of Leaf Cuticular Transpiration and Growth Traits in Field-Grown Wheat and Aegilops Genetic Resources




Abstract
1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. Field Vegetation Experiments
2.3. Analyses of Growth Parameters
2.4. Determination of Cuticular Transpiration
2.5. Statistical Analyses
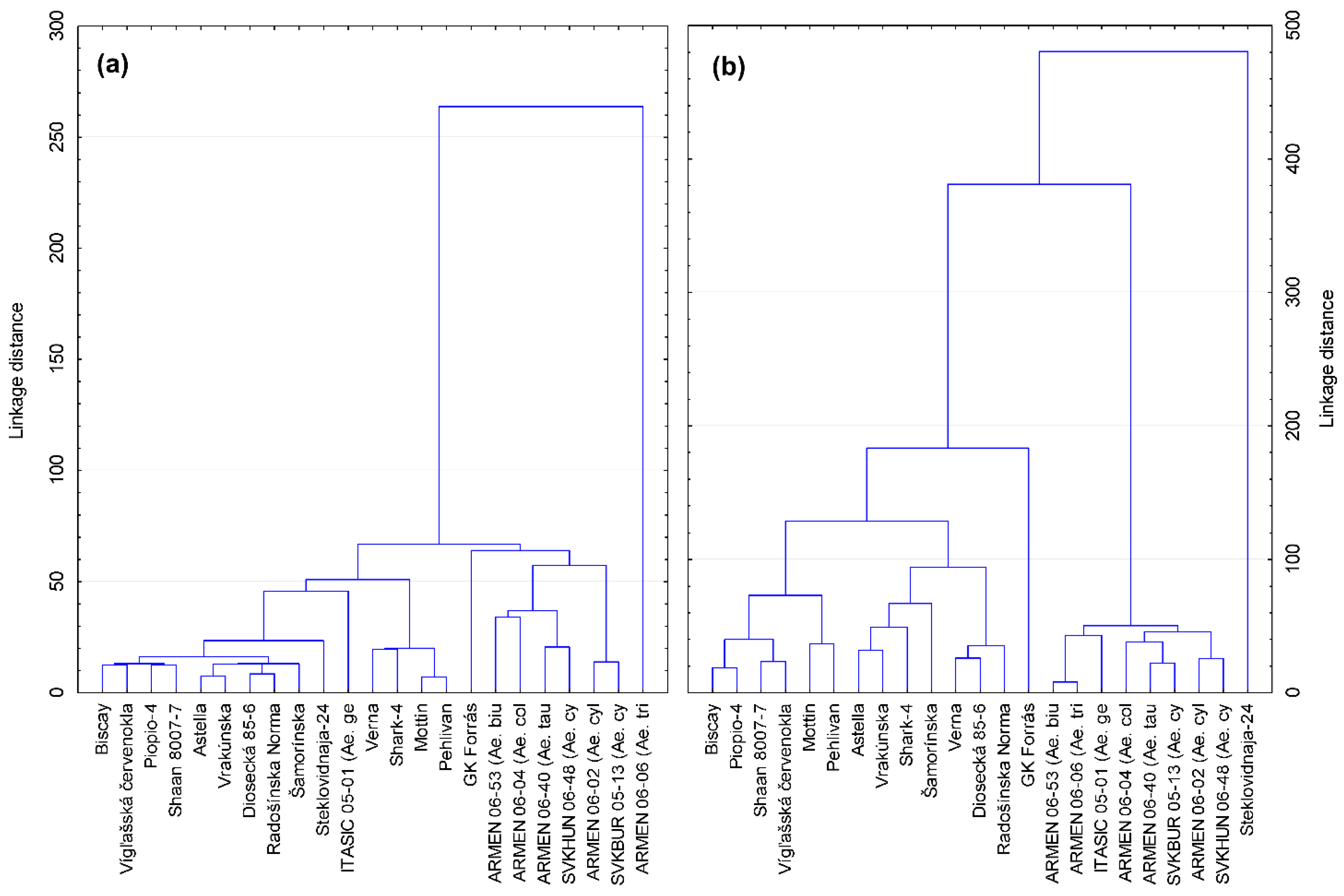
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Riederer, M.; Schreiber, L. Protecting against water loss: Analysis of the barrier properties of plant cuticles. J. Exp. Bot. 2001, 52, 2023–2032. [Google Scholar] [CrossRef] [PubMed]
- Leide, J.; Hildebrandt, U.; Vogg, G.; Riederer, M. The positional sterile (ps) mutation affects cuticular transpiration and wax biosynthesis of tomato fruits. J. Plant Physiol. 2011, 168, 871–877. [Google Scholar] [CrossRef]
- Schönherr, J. Resistance of plant surfaces to water loss: Transport properties of cutin, suberin and associated lipids. In Physiological Plant Ecology II. Encyclopedia of Plant Physiology (New Series); Lange, O.L., Nobel, P.S., Osmond, C.B., Ziegler, H., Eds.; Springer: Berlin/Heidelberg, Germany, 1982; Volume 12B, pp. 153–179. [Google Scholar]
- Goodwin, S.M.; Jenks, M.A. Plant cuticle function as a barrier to water loss. In Plant Abiotic Stress; Jenks, M., Hasegawa, P., Eds.; Blackwell Scientific Publishers: Oxford, UK, 2005; pp. 14–36. [Google Scholar]
- Kosma, D.K.; Jenks, M.A. Eco-physiological and molecular-genetic determinants of plant cuticle function in drought and salt stress tolerance. In Advances in Molecular Breeding towards Drought and Salt Tolerant Crops; Jenks, M.A., Hasegawa, P.M., Jain, S.M., Eds.; Springer: Dortrecht, The Netherlands, 2007; pp. 91–120. [Google Scholar]
- Huihui, B.; Kovalchuk, N.; Langridge, P.; Tricker, P.; Lopato, S.; Borisjuk, N. The impact of drought on wheat leaf cuticle properties. BMC Plant Biol. 2017, 17, 2–13. [Google Scholar] [CrossRef]
- Holmgren, P.; Jarvis, P.G.; Jarvis, M.S. Resistances to Carbon Dioxide and Water Vapor Transfer in Leaves of Different Plant Species. Physiol. Plant. 1965, 18, 557–573. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, X.; Du, Z.; Zhang, W.; Devkota, A.R.; Chen, Z.; Chen, C.; Sun, W.; Chen, M. A Proposed Method for Simultaneous Measurement of Cuticular Transpiration from Different Leaf Surfaces in Camellia sinensis. Front. Plant Sci. 2020, 11, 420. [Google Scholar] [CrossRef] [PubMed]
- Schuster, A.-C.; Burghardt, M.; Riederer, M. The ecophysiology of leaf cuticular transpiration: Are cuticular water permeabilities adapted to ecological conditions? J. Exp. Bot. 2017, 68, 5271–5279. [Google Scholar] [CrossRef]
- Petcu, E.; Maria, S.; Cirstea, V.E. The effect of water stress on cuticular transpiration and its association with alfalfa yield. Rom. Agric. Res. 2009, 26, 53–56. [Google Scholar]
- Jetter, R.; Schäffer, S.; Riederer, M. Leaf cuticular waxes are arranged in chemically and mechanically distinct layers: Evidence from Prunus laurocerasus L. Plant Cell Environ. 2000, 23, 619–628. [Google Scholar] [CrossRef]
- Canny, M.J.; Huang, C.X. Leaf water content and palisade cell size. New Phytol. 2006, 170, 75–85. [Google Scholar] [CrossRef]
- Ali, I.; Abbas, S.Q.; Hameed, M.; Naz, N.; Zafar, S.; Kanwal, S. Leaf anatomical adaptations in some exotic species of Eucalyptus l’hér. (Myrtaceae). Pak. J. Bot. 2009, 41, 2717–2727. [Google Scholar]
- Oteros, J.; García-Mozo, H.; Vázquez, L.; Mestre, A.; Dominguezvilches, E.; Galan, C. Modelling olive phenological response to weather and topography. Agric. Ecosyst. Environ. 2013, 179, 62–68. [Google Scholar] [CrossRef]
- Carignato, A.; Vázquez-Piqué, J.; Tapias, R.; Ruiz, F.; Fernández, M. Variability and Plasticity in Cuticular Transpiration and Leaf Permeability Allow Differentiation of Eucalyptus Clones at an Early Age. Forest 2020, 11, 9. [Google Scholar] [CrossRef]
- Kramer, P.J. Drought, stress and the origin of adaptation. In Adaptation of Plants to Water and High Temperature Stress; Turner, N., Kramer, P.J., Eds.; Academic Press: San Diego, CA, USA, 1980; pp. 7–20. [Google Scholar]
- Boyer, J.S. Plant Productivity and Environment. Science 1982, 218, 443–448. [Google Scholar] [CrossRef] [PubMed]
- David, M. Osmotic adjustment capacity and cuticular transpiration in several wheat cultivars cultivated in Algeria. Rom. Agric. Res. 2009, 26, 29–33. [Google Scholar]
- Dewey, D.R. The genomic system of classification as a guide to intergeneric hybridization with the perennial Triticeae. In Gene Manipulation in Plant Improvement; Gustafson, J.P., Ed.; Columbia University Press: New York, NY, USA, 1984; pp. 209–279. [Google Scholar]
- Chen, N.; Chen, W.-J.; Yan, H.; Wang, Y.; Kang, H.-Y.; Zhang, H.-Q.; Zhou, Y.-H.; Sun, G.-L.; Sha, L.-N.; Fan, X. Evolutionary patterns of plastome uncover diploid-polyploid maternal relationships in Triticeae. Mol. Phylogenet. Evol. 2020, 149, 106838. [Google Scholar] [CrossRef]
- Nazari, M.; Moosavi, S.S.; Maleki, M. Morpho-physiological and proteomic responses of Aegilops tauschii to imposed moisture stress. Plant Physiol. Biochem. 2018, 132, 445–452. [Google Scholar] [CrossRef]
- Asseng, S.; Guarin, J.R.; Raman, M.; Monje, O.; Kiss, G.; Despommier, D.D.; Meggers, F.M.; Gauthier, P.P.G. Wheat yield potential in controlled-environment vertical farms. Proc. Natl. Acad. Sci. USA 2020, 117, 19131–19135. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef]
- Gloser, J.; Barták, M.; Gloser, V. Changes in photosynthetic parameters of evolutionary different species of triticum and Aegilops grown at high and low nitrogen availability. In Photosynthesis: Mechanisms and Effects; Garab, G., Ed.; Kluwer Academic Publishers: Dortrecht, The Nederlands, 1998; Volume V, pp. 3789–3792. [Google Scholar]
- Van Slageren, M.W. Wild Wheats: A Monograph of Aegilops L. and Amblyopyrum (Jaub and Spach) Eig (Poaceae); Wageningen Agricultural University Papers: Aleppo, Syria, 1994; pp. 1–512. [Google Scholar]
- Molnár, I.; Gáspár, L.; Sárvári, E.; Dulai, S. Physiological and morphological responses to water stress in Aegilops biuncialis and Triticum aestivum genotypes with differing tolerance to drought. Funct. Plant Biol. 2004, 31, 1149–1159. [Google Scholar] [CrossRef]
- Zaharieva, M.; Gaulin, E.; Havaux, M.; Acevedo, E. Drought and heat responses in the wild wheat relative Aegilops geniculata Roth: Potential interest for wheat improvement. Crop Sci. 2001, 41. [Google Scholar] [CrossRef]
- Colmer, T.D.; Flowers, T.J.; Munns, R. Use of wild relatives to improve salt tolerance in wheat. J. Exp. Bot. 2006, 57, 1059–1078. [Google Scholar] [CrossRef] [PubMed]
- Brestic, M.; Zivcak, M.; Hauptvogel, P.; Misheva, S.; Kocheva, K.; Yang, X.; Li, X.; Allakhverdiev, S.I. Wheat plant selection for high yields entailed improvement of leaf anatomical and biochemical traits including tolerance to nonoptimal temper-ature conditions. Photosynth. Res. 2018, 136, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Monneveux, P.; Zaharieva, M.; Rekika, D.; Royo, C. The utilization of Triticum and Aegilops species for the improvement of durum wheat. In Durum Wheat Improvement in the Mediterranean Region: New Challenges; Royo, C., Nachit, M., Di Fonzo, N., Araus, J.L., Eds.; CIHEAM: Zaragoza, Spain, 2000; pp. 71–81. [Google Scholar]
- Leśniowska-Nowak, J.; Okoń, S.; Wieremczuk, A. Molecular diversity analysis of genotypes from four Aegilops species based on retrotransposon–microsatellite amplified polymorphism (REMAP) markers. Cereal Res. Commun. 2020, 49, 37–44. [Google Scholar] [CrossRef]
- Bultynck, L.; Fiorani, F.; Van Volkenburgh, E.; Lambers, H. Epidermal cell division and cell elongation in two Aegilops species with contrasting leaf elongation rates. Funct. Plant Biol. 2003, 30, 425–432. [Google Scholar] [CrossRef]
- Brestic, M.; Zivcak, M.; Kunderlikova, K.; Allakhverdiev, S.I. High temperature specifically affects the photoprotective responses of chlorophyll b-deficient wheat mutant lines. Photosynth. Res. 2016, 130, 251–266. [Google Scholar] [CrossRef] [PubMed]
- Chovancek, E.; Zivcak, M.; Botyanszka, L.; Hauptvogel, P.; Yang, X.; Misheva, S.; Hussain, S.; Brestic, M. Transient Heat Waves May Affect the Photosynthetic Capacity of Susceptible Wheat Genotypes Due to Insufficient Photosystem I Photoprotection. Plants 2019, 8, 282. [Google Scholar] [CrossRef] [PubMed]
- Chovancek, E.; Zivcak, M.; Brestic, M.; Hussain, S.; Allakhverdiev, S.I. The different patterns of post-heat stress responses in wheat genotypes: The role of the transthylakoid proton gradient in efficient recovery of leaf photosynthetic capacity. Photosynth. Res. 2021, 1–15. [Google Scholar] [CrossRef]
- Yul Yoo, C.; Pence, H.E.; Hasegawa, P.M.; Mickelbart, M.V. Regulation of Transpiration to Improve Crop Water Use. Crit. Rev. Plant Sci. 2009, 6, 410–431. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Shabala, L.; Zhou, M.; Brodribb, T.J.; Corkrey, R.; Shabala, S. Factors determining stomatal and non-stomatal (residual) transpiration and their contribution towards salinity tolerance in contrasting barley genotypes. Environ. Exp. Bot. 2018, 153, 10–20. [Google Scholar] [CrossRef]
- Andivia, E.; Carevic, F.; Fernandez, M.; Alejano, R.; Vázquez-Piqué, J.; Tapias, R. Seasonal evolution of water status after outplanting of two provenances of Holm oak nursery seedlings. New For. 2012, 43, 815–824. [Google Scholar] [CrossRef]
- David, M. Water loss from excised leaves in collection of Triticum aestivum and Triticum durum cultivars. Rom. Agric. Res. 2010, 27, 27–34. [Google Scholar]
- Burghardt, M.; Riederer, M. Ecophysiological relevance of cuticular transpiration of deciduous and evergreen plants in relation to stomatal closure and leaf water potential. J. Exp. Bot. 2003, 54, 1941–1949. [Google Scholar] [CrossRef] [PubMed]
- Poorter, H.; Niinemets, Ü.; Poorter, L.; Wright, I.J.; Villar, R. Causes and consequences of variation in leaf mass per area (LMA): A meta-analysis. New Phytol. 2009, 182, 565–588. [Google Scholar] [CrossRef] [PubMed]
- Witkowski, E.T.F.; Lamont, B.B. Leaf specific mass confounds leaf density and thickness. Oecologia 1991, 88, 486–493. [Google Scholar] [CrossRef] [PubMed]
- De La Riva, E.G.; Tosto, A.; Pérez-Ramos, I.M.; Navarro-Fernández, C.M.; Olmo, M.; Anten, N.P.R.; Marañón, T.; Villar, R. A plant economics spectrum in Mediterranean forests along environmental gradients: Is there coordination among leaf, stem and root traits? J. Veg. Sci. 2016, 27, 187–199. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Lin, S.; Niklas, K.J.; Wan, Y.; Hölscher, D.; Hui, C.; Ding, Y.; Shi, P. Leaf shape influences the scaling of leaf dry mass vs. area: A test case using bamboos. Ann. For. Sci. 2020, 77, 1–15. [Google Scholar] [CrossRef]
- Oren, R.; Schulze, E.-D.; Matyssek, R.; Zimmermann, R. Estimating photosynthetic rate and annual carbon gain in conifers from specific leaf weight and leaf biomass. Oecologia 1986, 70, 178–193. [Google Scholar] [CrossRef]
- Reich, P.B.; Walters, M.B.; Ellsworth, D.S. From tropics to tundra: Global convergence in plant functioning. Proc. Natl. Acad. Sci. USA 1997, 94, 13730–13734. [Google Scholar] [CrossRef]
- Quero, J.L.; Villar, R.; Marañón, T.; Zamora, R. Interactions of drought and shade effects on seedlings of four Quercus species: Physiological and structural leaf responses. New Phytol. 2006, 170, 819–834. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B.; Walters, M.B.; Ellsworth, D.S. Leaf Life-Span in Relation to Leaf, Plant, and Stand Characteristics among Diverse Ecosystems. Ecol. Monogr. 1992, 62, 365–392. [Google Scholar] [CrossRef]
- Poorter, H.; Van der Werf, A. Is inherent variation in RGR determined by LAR at low irradiance and by NAR at high irradiance? A review of herbaceous species. In Inherent Variation in Plant Growth; Lambers, H., Poorter, H., Van Vuuren, M.M.I., Eds.; Backhuys Publishers: Leiden, The Netherland, 1998; pp. 309–336. [Google Scholar]
- Antúnez, I.; Retamosa, E.C.; Villar, R. Relative growth rate in phylogenetically related deciduous and evergreen woody species. Oecologia 2001, 128, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Robleto, J.; Villar, R. Relative Growth Rate and Biomass Allocation in Ten Woody Species with Different Leaf Longevity Using Phylogenetic Independent Contrasts (PICs). Plant Biol. 2005, 7, 484–494. [Google Scholar] [CrossRef]
- Lopez-Iglesias, B.; Olmo, M.; Gallardo, A.; Villar, R. Short-term effects of litter from 21 woody species on plant growth and root development. Plant Soil 2014, 381, 177–191. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Thompson, K. Functional leaf attributes predict litter decomposition rate in herbaceous plants. New Phytol. 1997, 135, 109–114. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Perez-Harguindeguy, N.; Diaz, S.; Grime, J.P.; Marzano, B.; Cabido, M.; Vendramini, F.; Cerabolini, B. Leaf structure and defence control litter decomposition rate across species and life forms in regional floras on two continents. New Phytol. 1999, 143, 191–200. [Google Scholar] [CrossRef]
- Zhang, B.; Xiaozhen, L.; Jiang, J.; DeAngelis, D.L.; Fu, Z.; Zhang, J. Similarity of plant functional traits and aggregation pattern in a subtropical forest. Ecol. Evol. 2017, 7, 4086–4098. [Google Scholar] [CrossRef]
- McNeal, F.H.; Berg, M.A. Flag leaf area in five spring wheat crosses and the relationship to grain yield. Euphytica 1977, 26, 739–744. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Davies, N.W.; Shabala, L.; Zhou, M.; Brodribb, T.J.; Shabala, S. Residual transpiration as a component of salinity stress tolerance mechanism: A case study for barley. BMC Plant Biol. 2017, 17, 1–12. [Google Scholar] [CrossRef]
- Li, H.; Madden, J.L.; Potts, B.M. Variation in leaf waxes of the Tasmanian Eucalyptus species—I. Subgenus Symphyomyrtus. Biochem. Syst. Ecol. 1997, 25, 631–657. [Google Scholar] [CrossRef]
- Migacz, I.P.; Raeski, P.A.; Almeida, V.P.; Raman, V.; Nisgoski, S.; Muniz, G.I.B.; Farago, P.V.; Khan, I.A.; Budel, J.M. Comparative leaf morpho-anatomy of six species of Eucalyptus cultivated in Brazil. Rev. Bras. Farm. 2018, 28, 273–281. [Google Scholar] [CrossRef]
- Berry, C.Z.; Emery, N.C.; Gotsch, S.G.; Goldsmith, G.R. Foliar water uptake: Processes, pathways, and integration into plant water budgets. Plant Cell Environ. 2018, 42, 410–423. [Google Scholar] [CrossRef] [PubMed]
- Kerstiens, G. Diffusion of water vapour and gases across cuticles and through stomatal pores presumed closed. In Plant Cu-ticles: An Integrated Functional Approach; BIOS Scientific: Oxford, UK, 1996; pp. 121–134. [Google Scholar]


{kind=link}
| Genotype | Cuticular Transpiration (CT) (g m−2 h−1) | Dry Mass Per Leaf Area (MLA) (g m−2) | Flag Leaf Area (FLA) (cm2) | Plant Leaf Area (PLA) (cm2) |
|---|---|---|---|---|
| Group—modern varieties | ||||
| Biscay | 11.7 ± 0.7 a | 44.3 ± 2.0 efg | 28.6 ± 2.5 h | 278 ± 18 efgh |
| GK Forrás | 20.1 ± 1.2 c | 39.6 ± 1.3 ab | 22.4 ± 1.1 f | 346 ± 20 ghi |
| Piopio-4 | 12.9 ± 0.7 ab | 43.7 ± 0.7 ef | 18.0 ± 0.6 de | 307 ± 23 fghi |
| Astella | 16.6 ± 0.4 abc | 49.2 ± 0.8 gh | 29.0 ± 2.1 h | 279 ± 12 efgh |
| Verna | 16.9 ± 1.1 abc | 42.3 ± 1.0 ef | 26.9 ± 1.5 gh | 517 ± 30 j |
| Steklovidnaja-24 | 19.5 ± 2.0 c | 44.3 ± 1.9 efg | 30.8 ± 2.2 hi | 251 ± 11 ef |
| Mottin | 12.9 ± 0.9 ab | 40.8 ± 1.2 cde | 24.3 ± 2.3 fg | 489 ± 14 j |
| Pehlivan | 18.1 ± 0.7 bc | 40.0 ± 1.4 bcde | 34.3 ± 1.9 i | 364 ± 36 hi |
| Shaan 8007-7 | 16.4 ± 0.6 abc | 52.3 ± 1.5 h | 22.6 ± 1.0 f | 269 ± 18 efgh |
| Shark-4 | 20.1 ± 1.8 c | 46.7 ± 0.6 fg | 16.2 ± 1.2 d | 383 ± 4 i |
| Average (modern varieties) | 16.5 | 44.3 | 25.3 | 348 |
| Group—landraces | ||||
| Diosecká 85-6 | 16.6 ± 0.8 abc | 42.7 ± 2.9 ef | 21.6 ± 0.6 ef | 333 ± 32 fghi |
| Vigľašská červenoklasá | 18.0 ± 3.2 bc | 32.1 ± 1.4 a | 21.8 ± 1.8 ef | 349 ± 54 ghi |
| Šamorínska | 21.4 ± 1.4 c | 41.7 ± 1.9 def | 20.8 ± 2.1 ef | 212 ± 36 de |
| Vrakúnska | 17.6 ± 0.8 bc | 38.8 ± 1.8 bcde | 27.2 ± 0.3 gh | 282 ± 19 efgh |
| Radošínska Norma | 21.4 ± 0.5 c | 41.5 ± 1.4 def | 22.8 ± 1.1 f | 265 ± 12 efgh |
| Average (landraces) | 19.0 | 39.3 | 22.8 | 288 |
| Group—Aegilops | ||||
| ARMEN06-53 (Ae. biuncialis) | 40.4 ± 2.8 e | 39.1 ± 1.5 bcde | 3.8 ± 0.1 a | 48 ± 4 a |
| ARMEN06-04 (Ae. columnaris) | 33.8 ± 1.7 d | 44.1 ± 1.7 efg | 14.2 ± 0.1 cd | 90 ± 28 abc |
| ARMEN06-02 (Ae. cylindrica) | 28.9 ± 2.9 d | 34.8 ± 1.1 ab | 10.5 ± 0.6 bc | 85 ± 7 abc |
| ITASIC05-01 (Ae. geniculata) | 22.3 ± 1.6 c | 43.8 ± 1.6 ef | 2.6 ± 0.3 a | 40 ± 0.3 a |
| ARMEN06-40 (Ae. tauschii) | 18.4 ± 2.5 bc | 39.1 ± 0.9 bcde | 7.7 ± 0.3 b | 157 ± 38 bcd |
| ARMEN06-06 (Ae. triuncialis) | 46.1 ± 2.7 f | 36.6 ± 1.1 abcd | 2.6 ± 0.1 a | 82 ± 11 abc |
| SVKBUR05-13 (Ae. cylindrica) | 16.6 ± 1.5 abc | 32.3 ± 2.8 a | 9.8 ± 0.4 b | 136 ± 32 bcd |
| SVKHUN06-48 (Ae. cylindrica) | 20.7 ± 1.1 c | 35.8 ± 1.7 abc | 10.5 ± 0.7 b | 167 ± 35 cd |
| Average (Aegilops) | 28.4 | 38.2 | 7.7 | 101 |
| Genotype | Dry Mass Per Ground Area (DM) (g m−2) | LAI (m2 m−2) | Water Loss by CT Per Ground Area (g m-2) | Water Loss by CT Per Dry Mass Unit (mg g−1) |
|---|---|---|---|---|
| Group—modern varieties | ||||
| Biscay | 978 ± 67 | 5.0 ± 0.3 efgh | 58.6 ± 5.2 cdef | 92.0 ± 8.1 a |
| GK Forrás | 795 ± 45 | 6.2 ± 0.4 ghi | 125.2 ± 10.4 kl | 191.7 ± 20.2 g |
| Piopio-4 | 994 ± 77 | 5.5 ± 0.4 fghi | 71.3 ± 6.8 efgh | 110.2 ± 10.5 bcde |
| Astella | 1280 ± 53 | 5.0 ± 0.2 efgh | 83.7 ± 4.1 ghi | 100.4 ± 4.9 a |
| Verna | 1553 ± 90 | 9.3 ± 0.5 j | 157.8 ± 14.0 m | 156.0 ± 13.8 ef |
| Steklovidnaja-24 | 2059 ± 98 | 4.5 ± 0.2 ef | 88.1 ± 10.0 hi | 65.7 ± 7.5 a |
| Mottin | 1117 ± 20 | 8.8 ± 0.2 j | 113.7 ± 8.5 jk | 156.5 ± 11.7 bcd |
| Pehlivan | 1152 ± 111 | 6.6 ± 0.6 hi | 118.5 ± 12.4 jkl | 158.0 ± 16.5 def |
| Shaan 8007-7 | 1032 ± 92 | 4.8 ± 0.4 efg | 79.7 ± 7.8 fghi | 118.7 ± 11.5 abc |
| Shark-4 | 1358 ± 16 | 6.9 ± 0.1 i | 138.5 ± 12.4 lm | 156.6 ± 14.0 def |
| Average—modern varieties | 1232 | 6.27 | 103.5 | 135.6 |
| Group—landraces | ||||
| Diosecká 85-6 | 1579 ± 137 | 6.0 ± 0.5 fghi | 99.7 ± 9.9 ij | 97.0 ± 9.7 a |
| Vigľašská červenoklasá | 1044 ± 186 | 6. 3 ± 1.1 ghi | 67.1 ± 7.8 defgh | 98.8 ± 11.5 a |
| Šamorínska | 1424 ± 256 | 3.8 ± 0.7 de | 81.8 ± 5.4 fghi | 88.2 ± 5.8 a |
| Vrakúnska | 1311 ± 100 | 5.1 ± 0.4 efgh | 89.5 ± 8.0 hi | 104.9 ± 9.4 a |
| Radošínska Norma | 1519 ± 76 | 4.8 ± 0.2 efg | 102. ± 5.5 ijk | 103.6 ± 5.6 ab |
| Average—landraces | 1375 | 5.2 | 88.1 | 98.5 |
| Group—Aegilops | ||||
| ARMEN06-53 (Ae. biuncialis) | 193 ± 18 | 0.86 ± 0.08 a | 34.1 ± 2.8 ab | 271.6 ± 22.1 gh |
| ARMEN06-04 (Ae. columnaris) | 285 ± 90 | 1.63 ± 0.52 abc | 55.1 ± 2.8 bcde | 297.6 ± 15.2 h |
| ARMEN06-02 (Ae. cylindrica) | 389 ± 31 | 1.52 ± 0.12 abc | 44.0 ± 4.3 bcd | 174.0 ± 17.2 f |
| ITASIC05-01 (Ae. geniculata) | 236 ± 24 | 0.72 ± 0.07 a | 16.2 ± 2.1 a | 105.3 ± 13.9 a |
| ARMEN06-40 (Ae. tauschii) | 322 ± 83 | 2.83 ± 0.72 bcd | 52.0 ± 7.1 bcde | 247.9 ± 33.9 g |
| ARMEN06-06 (Ae. triuncialis) | 185 ± 27 | 1.47 ± 0.22 ab | 67.7 ± 4.0 defgh | 560.8 ± 33.0 i |
| SVKBUR05-13 (Ae. cylindrica) | 343 ± 78 | 2.44 ± 0.55 bcd | 39.6 ± 3.7 bc | 177.5 ± 6.7 f |
| SVKHUN06-48 (Ae. cylindrica) | 414 ± 89 | 3.01 ± 0.64 cd | 62.1 ± 3.4 cdefg | 230.2 ± 12.7 g |
| Average—Aegilops | 296 | 1.81 | 46.35 | 258.1 |
| Cuticular Transpiration | Dry Mass Per Plant | Dry Mass Per Leaf Area | Plant Leaf Area | Leaf Area Index | Flag Leaf Area | Water Loss by CT Per m2 | Water Loss by CT Per DM | |
|---|---|---|---|---|---|---|---|---|
| Cuticular transpiration | - | −0.55 * | −0.23 | −0.65 * | −0.65 * | −0.63 * | −0.34 | 0.79 * |
| Dry mass per plant | −0.55 * | - | 0.41 | 0.72 * | 0.72 * | 0.82 * | 0.70 * | −0.65 * |
| Dry mass per leaf area | −0.23 | 0.41 | - | 0.19 | 0.19 | 0.33 | 0.21 | −0.34 |
| Plant leaf area | −0.65 * | 0.72 * | 0.19 | - | 0.99 * | 0.75 * | 0.87 * | −0.42 * |
| Leaf area index | −0.65 * | 0.72 * | 0.19 | 0.99 * | - | 0.75 * | 0.87 * | −0.42 * |
| Flag leaf area | −0.63 * | 0.82 * | 0.33 | 0.75 * | 0.75 * | - | 0.65 * | −0.60 * |
| Water loss by CT per m2 | −0.34 | 0.70 * | 0.21 | 0.87 * | 0.87 * | 0.65 * | - | −0.16 |
| Water loss by CT per DM | 0.79 * | −0.65 * | −0.34 | −0.42 * | −0.42 * | −0.60 * | −0.16 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gašparovič, K.; Živčák, M.; Brestič, M.; Hauptvogel, P. Diversity of Leaf Cuticular Transpiration and Growth Traits in Field-Grown Wheat and Aegilops Genetic Resources. Agronomy 2021, 11, 522. https://doi.org/10.3390/agronomy11030522
Gašparovič K, Živčák M, Brestič M, Hauptvogel P. Diversity of Leaf Cuticular Transpiration and Growth Traits in Field-Grown Wheat and Aegilops Genetic Resources. Agronomy. 2021; 11(3):522. https://doi.org/10.3390/agronomy11030522
Chicago/Turabian StyleGašparovič, Kristína, Marek Živčák, Marián Brestič, and Pavol Hauptvogel. 2021. "Diversity of Leaf Cuticular Transpiration and Growth Traits in Field-Grown Wheat and Aegilops Genetic Resources" Agronomy 11, no. 3: 522. https://doi.org/10.3390/agronomy11030522
APA StyleGašparovič, K., Živčák, M., Brestič, M., & Hauptvogel, P. (2021). Diversity of Leaf Cuticular Transpiration and Growth Traits in Field-Grown Wheat and Aegilops Genetic Resources. Agronomy, 11(3), 522. https://doi.org/10.3390/agronomy11030522