
Comparison of Methods to Evaluate Rice (Oryza sativa) Germplasm for Tolerance to Low Temperature at the Seedling Stage


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
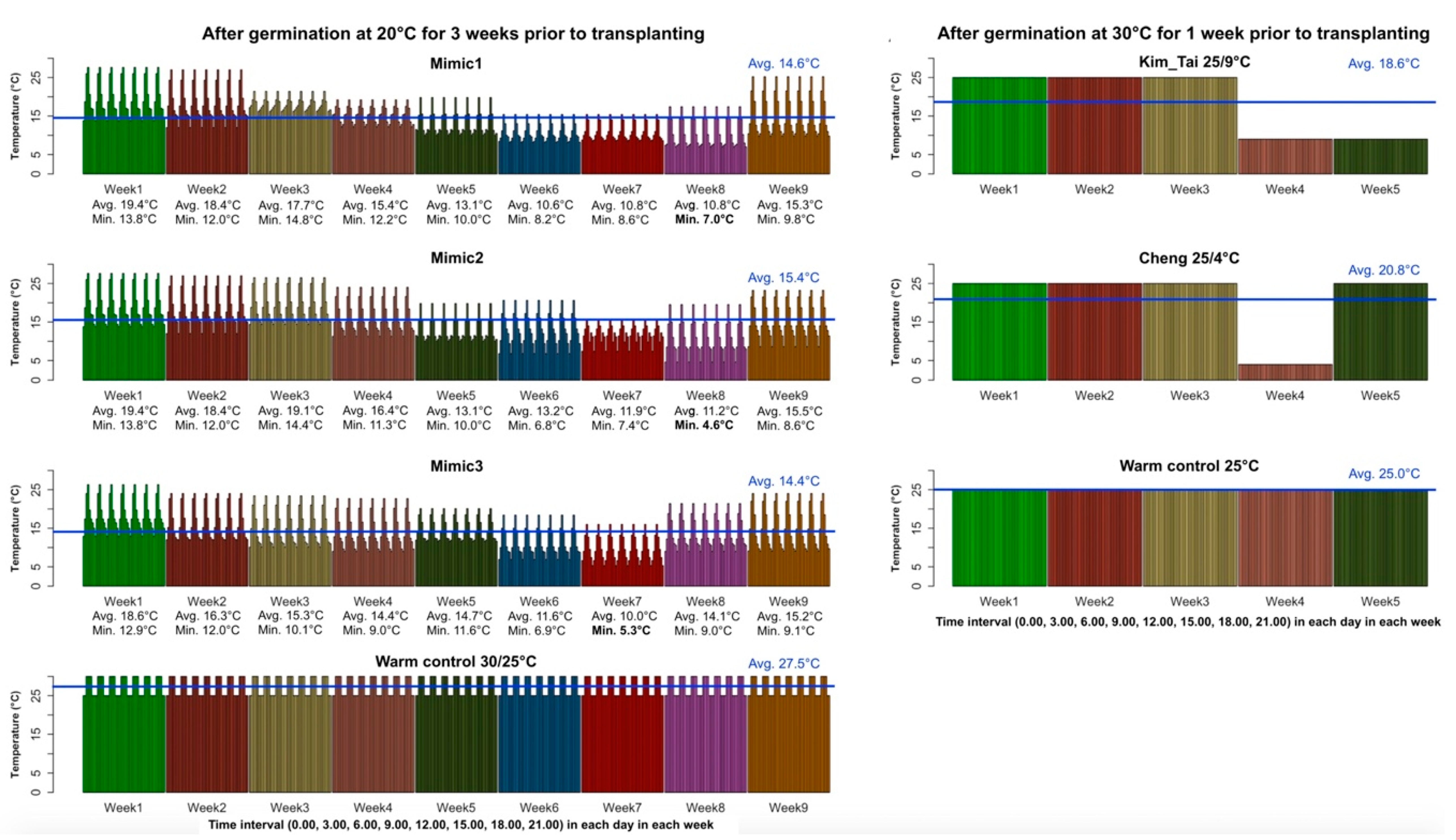
2.2. Controlled Environment Screens
2.3. Experimental Design
2.4. Statistical Analysis
3. Results
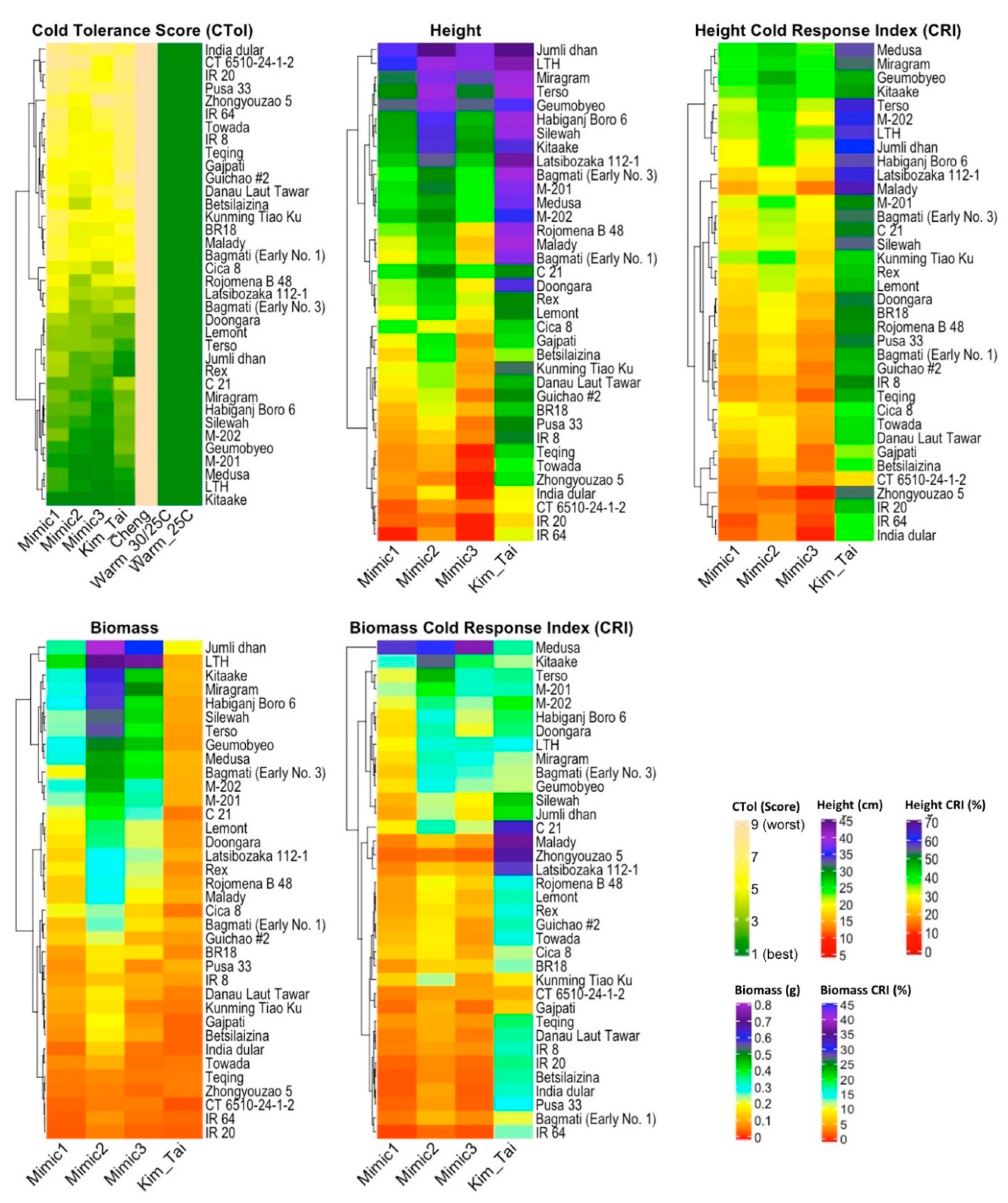
3.1. Germination Stage Screening
3.2. Seedling Stage Screening
3.3. Comparison of Germination Stage and Seedling Stage Screens
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- GRiSP (Global Rice Science Partnership). Rice Almanac, 4th ed.; International Rice Research Institute: Los Baños, Philippines, 2013; p. 283. [Google Scholar]
- Sundaram, J.K. Lessons from the 2008 World Food Crisis. Econ. Political Wkly. 2010, 45, 35–40. [Google Scholar]
- Kaneda, C.; Beachell, H.M. Response of indica-japonica rice hybrids to low temperatures. SABRAO J. 1974, 6, 17–32. [Google Scholar]
- Mackill, D.J.; Lei, X. Genetic variation for traits related to temperate adaptation of rice cultivar. Crop Sci. 1997, 37, 1340–1346. [Google Scholar] [CrossRef]
- Kaw, R.N.; Khush, G.S. Combining ability for low-temperature tolerance in rice. In Rice Genetics I; International Rice Research Institute: Los Baños, Philippines, 1986; pp. 593–612. [Google Scholar]
- Suh, J.P.; Lee, C.K.; Lee, J.H.; Kim, J.J.; Kim, S.M.; Cho, Y.C.; Park, S.H.; Shin, J.C.; Kim, Y.G.; Jena, K.K. Identification of quantitative trait loci for seedling cold tolerance using RILs derived from a cross between japonica and tropical japonica rice cultivars. Euphytica 2012, 184, 101–108. [Google Scholar] [CrossRef]
- Hossain, M.; Naher, F.; Shahabuddin, Q. Food Security and Nutrition in Bangladesh: Progress and Determinants. Electron. J. Agric. Dev. Econ. 2005, 2, 103–132. [Google Scholar]
- Li, T.G.; Visperas, R.M.; Vergara, B.S. Correlation of cold tolerance at different growth stages in rice. Acta Bot. Sin. 1981, 23, 203–207. [Google Scholar]
- Bangladesh: Disaster Report; Disaster Management Bureau: Dhaka, 2011; Available online: https://www.disasterforum.org/files/Bangladesh_Disaster_Report_2011.pdf (accessed on 9 May 2019).
- Andaya, V.C.; Mackill, D.J. Mapping of QTLs associated with cold tolerance during the vegetative stage in rice. J. Exp. Bot. 2003, 54, 2579–2585. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.H.; Li, S.; Wei, L.; Wei, C.; Zhu, Y.G. A major QTL conferring cold tolerance at the early seedling stage using recombinant inbred lines of rice (Oryza sativa L.). Plant Sci. 2005, 168, 527–534. [Google Scholar] [CrossRef]
- Han, L.; Qiao, Y.; Zhang, S.; Zhang, Y.; Cao, G.; Kim, J.; Lee, K.; Koh, H. Identification of Quantitative Trait Loci for Cold Response of Seedling Vigor Traits in Rice. J. Genet. Genom. 2007, 34, 239–246. [Google Scholar] [CrossRef]
- Lou, Q.; Chen, L.; Sun, Z.; Xing, Y.; Li, J.; Xu, X.; Mei, H.; Luo, L. A major QTL associated with cold tolerance at seedling stage in rice (Oryza sativa L.). Euphytica 2007, 158, 87–94. [Google Scholar] [CrossRef]
- Jiang, L.; Xun, M.; Wang, J.; Wan, J. QTL analysis of cold tolerance at seedling stage in rice (Oryza sativa L.) using recombination inbred lines. J. Cereal Sci. 2008, 48, 173–179. [Google Scholar] [CrossRef]
- Kim, S.I.; Tai, T.H. Evaluation of seedling cold tolerance in rice cultivars: A comparison of visual ratings and quantitative indicators of physiological changes. Euphytica 2011, 178, 437–447. [Google Scholar] [CrossRef]
- Cheng, L.; Wang, J.; Uzokwe, V.; Meng, L.; Wang, Y.; Sun, Y.; Zhu, L.; Xu, J.; Li, Z. Genetic Analysis of Cold Tolerance at Seedling Stage and Heat Tolerance at Anthesis in Rice (Oryza sativa L.). J. Integr. Agric. 2012, 11, 359–367. [Google Scholar] [CrossRef]
- Kim, S.M.; Suh, J.P.; Lee, C.K.; Lee, C.K.; Kim, Y.G.; Jena, K.K. QTL mapping and development of candidate gene-derived DNA markers associated with seedling cold tolerance in rice (Oryza sativa L.). Mol. Genet. Genom. 2014, 289, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhang, S.; Dong, J.; Yang, T.; Mao, X.; Liu, Q.; Wang, X.; Liu, B. A novel functional gene associated with cold tolerance at the seedling stage in rice. Plant Biotechnol. J. 2017, 15, 1141–1148. [Google Scholar] [CrossRef]
- Lv, Y.; Guo, Z.; Li, X.; Ye, H.; Li, X.; Xiong, L. New insights into the genetic basis of natural chilling and cold shock tolerance in rice by genome-wide association analysis. Plant Cell Environ. 2016, 39, 556–570. [Google Scholar] [CrossRef] [PubMed]
- International Rice Research Institute. Standard Evaluation System for Rice; International Rice Research Institute: Los Baños, Philippines, 1996. [Google Scholar]
- Zhang, F.; Ma, X.-F.; Gao, Y.-M.; Hao, X.-B.; Li, Z.-K. Genome-wide response to selection and genetic basis of cold tolerance in rice (Oryza sativa L.). BMC Genet. 2014, 15, 55. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Lenth, R. Emmeans: Estimated Marginal Means, Aka Least-Squares Means; 2019; Available online: http://th.archive.ubuntu.com/cran/web/packages/emmeans/index.html (accessed on 20 February 2021).
- Revelle, W. Psych: Procedures for Psychological, Psychometric, and Personality Research; Northwestern University: Evanston, IL, USA, 2018. [Google Scholar]
- Gu, Z.; Eils, R.; Schlesner, M. Complex Heatmaps Reveal Patterns and Correlations in Multidimensional Genomic Data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Sharifi, P. Evaluation on Sixty-Eight Rice Germplasms in Cold Tolerance at Germination. Rice Sci. 2010, 17, 77–81. [Google Scholar] [CrossRef]
- Dashtmain, F.P.; Hosseini, K.M.; Esfahani, M. Methods for rice genotypes cold tolerance evaluation at germination stage. Int. J. Agric. Crop Sci. 2013, 5, 2111–2116. [Google Scholar]
- Lone, J.A.; Khan, M.N.; Bhat, M.A.; Shikari, A.B.; Wani, S.H.; Sofi, N.R.; Khan, M.I.; Lone, R.A. Cold Tolerance at Germination and Seedling Stages of Rice: Methods of Evaluation and Characterization of Thirty Rice Genotypes under Stress Conditions. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 1103–1109. [Google Scholar] [CrossRef]
- Da Cruz, R.P.; Milach, S.C.K. Cold tolerance at the germination stage of rice: Methods of evaluation and characterization of genotypes. Sci. Agric. 2004, 61, 1–8. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Random Model | Fixed Model | |||||
|---|---|---|---|---|---|---|---|
| % of Total Variance | Mean Sq | ||||||
| Germination Proportion | Shoot Length | Root Length | Df | Germination Proportion | Shoot Length | Root Length | |
| Cultivar | 27.1 *** | 55.1 *** | 35.7 *** | 35 | 0.083731 *** | 30.8441 *** | 8.1656 *** |
| Treatment | 3.7 * | 3.4 NS | 10.0 ** | 1 | 0.155715 *** | 23.9260 *** | 20.2986 *** |
| Cultivar:Treatment | 4.0 NS | 19.7 *** | 25.9 *** | 35 | 0.016230 NS | 3.7019 *** | 1.4950 *** |
| Residual | 65.1 | 21.8 | 28.4 | 395 | 0.012165 | 0.6447 | 0.2652 |
| All Treatments (5 Cold and 2 Warm) | Mimic1, Mimic2, Mimic3 and Kim_Tai | Mimic1, Mimic2 and Mimic3 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Source | CTol | Plant Height | Biomass | CTol | Plant Height | Biomass | Height CRI | Biomass CRI | CTol | Plant Height | Biomass | Height CRI | Biomass CRI |
| Chamber | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Cultivar | 12.3 *** | 7.5 *** | 1.5 * | 66.3 *** | 48.6 *** | 30.3 *** | 16.9 *** | 19.8 *** | 67.4 *** | 63.6 *** | 42.3 *** | 55.9 *** | 51.8 *** |
| Treatment | 65.9 ** | 82.7 *** | 78.8 *** | 2.1 | 19.3 | 10.3 | 50.2 * | 12.7 | 3.3 | 5.4 | 3.7 | 6.6 | 2.5 |
| Treatment:Chamber | 2.5 *** | 1.9 *** | 0.8 *** | 4.7 *** | 12.2 *** | 24.9 *** | 8.3 *** | 13.1 *** | 4.8 *** | 15.7 *** | 25.4 *** | 18.6 *** | 19.6 *** |
| Cultivar:Chamber | 0.0 | 0.0 | 0.0 | 2.1 | 0.0 | 0.0 | 10.8 *** | 0. 0 | 0.2 | 0.0 | 0.0 | 0.0 | 0.0 |
| Cultivar:Treatment | 8.6 *** | 3.2 *** | 12.3 *** | 2.1 ** | 1.9 * | 7.4 *** | 0.0 | 16.0 *** | 1.3 * | 0.1 | 0.7 | 0.0 | 0.0 |
| Cultivar:Treatment: Chamber | 3.1 *** | 0.1 | 0.0 | 3.4 *** | 3.4 *** | 11.8 *** | 0.0 | 8.5 *** | 2.2 ** | 3.3 *** | 11.5 *** | 3.4 *** | 10.0 *** |
| Trays within chamber | 0.2 * | 0.1 * | 0.0 | 0.6 * | 0.2 * | 0.2 | 0.2 * | 0.2 | 0.7 * | 0.1 | 0.3 | 0.2 | 0.2 |
| Chamber:Treatment:Trays within chamber | 0.6 *** | 0.2 *** | 0.5 *** | 1.2 *** | 0.2 | 1.3 *** | 0.1 | 0.7 *** | 1.3 *** | 0.3 * | 1.3 *** | 0.4 * | 0.9 *** |
| Residual | 6.9 | 4.3 | 6.0 | 17.6 | 14.3 | 13.9 | 13.5 | 29.0 | 18.8 | 11.4 | 14.7 | 14.9 | 15.0 |
| Total | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 |
| Source | All Treatments (5 Cold and 2 Warm-Controls) | Mimic1, Mimic2, Mimic3 and Kim_Tai (Best 4 Cold Treatments) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CTol | Plant Height | Biomass | CTol | Plant Height | Biomass | Height CRI | Biomass CRI | |||
| NuDf | Mean sq | Mean sq | Mean sq | NuDf | Mean sq | Mean sq | Mean sq | Mean sq | Mean sq | |
| Cultivar | 35 | 17.03 *** | 2361.35 *** | 2.35 *** | 35 | 62.43 *** | 1179.03 *** | 0.14 *** | 1081.05 *** | 403.81 *** |
| Treatment | 6 | 20.29 * | 1672.18 *** | 7.65 *** | 3 | 2.27 NS | 75.99 NS | 0.013 NS | 469.53 ** | 70.70 NS |
| Cultivar:Treatment | 210 | 2.74 *** | 160.19 *** | 0.95 *** | 105 | 1.58 ** | 29.31 *** | 0.015 *** | 103.84 *** | 82.15 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, N.; Reinke, R.; Sacks, E.J. Comparison of Methods to Evaluate Rice (Oryza sativa) Germplasm for Tolerance to Low Temperature at the Seedling Stage. Agronomy 2021, 11, 385. https://doi.org/10.3390/agronomy11020385
Sharma N, Reinke R, Sacks EJ. Comparison of Methods to Evaluate Rice (Oryza sativa) Germplasm for Tolerance to Low Temperature at the Seedling Stage. Agronomy. 2021; 11(2):385. https://doi.org/10.3390/agronomy11020385
Chicago/Turabian StyleSharma, Nirmal, Russell Reinke, and Erik J. Sacks. 2021. "Comparison of Methods to Evaluate Rice (Oryza sativa) Germplasm for Tolerance to Low Temperature at the Seedling Stage" Agronomy 11, no. 2: 385. https://doi.org/10.3390/agronomy11020385
APA StyleSharma, N., Reinke, R., & Sacks, E. J. (2021). Comparison of Methods to Evaluate Rice (Oryza sativa) Germplasm for Tolerance to Low Temperature at the Seedling Stage. Agronomy, 11(2), 385. https://doi.org/10.3390/agronomy11020385