Physiological and Morphometric Response of Forage Grass Species and Their Biomass Distribution Depending on the Term and Frequency of Water Deficiency


Abstract
:1. Introduction
2. Materials and Methods
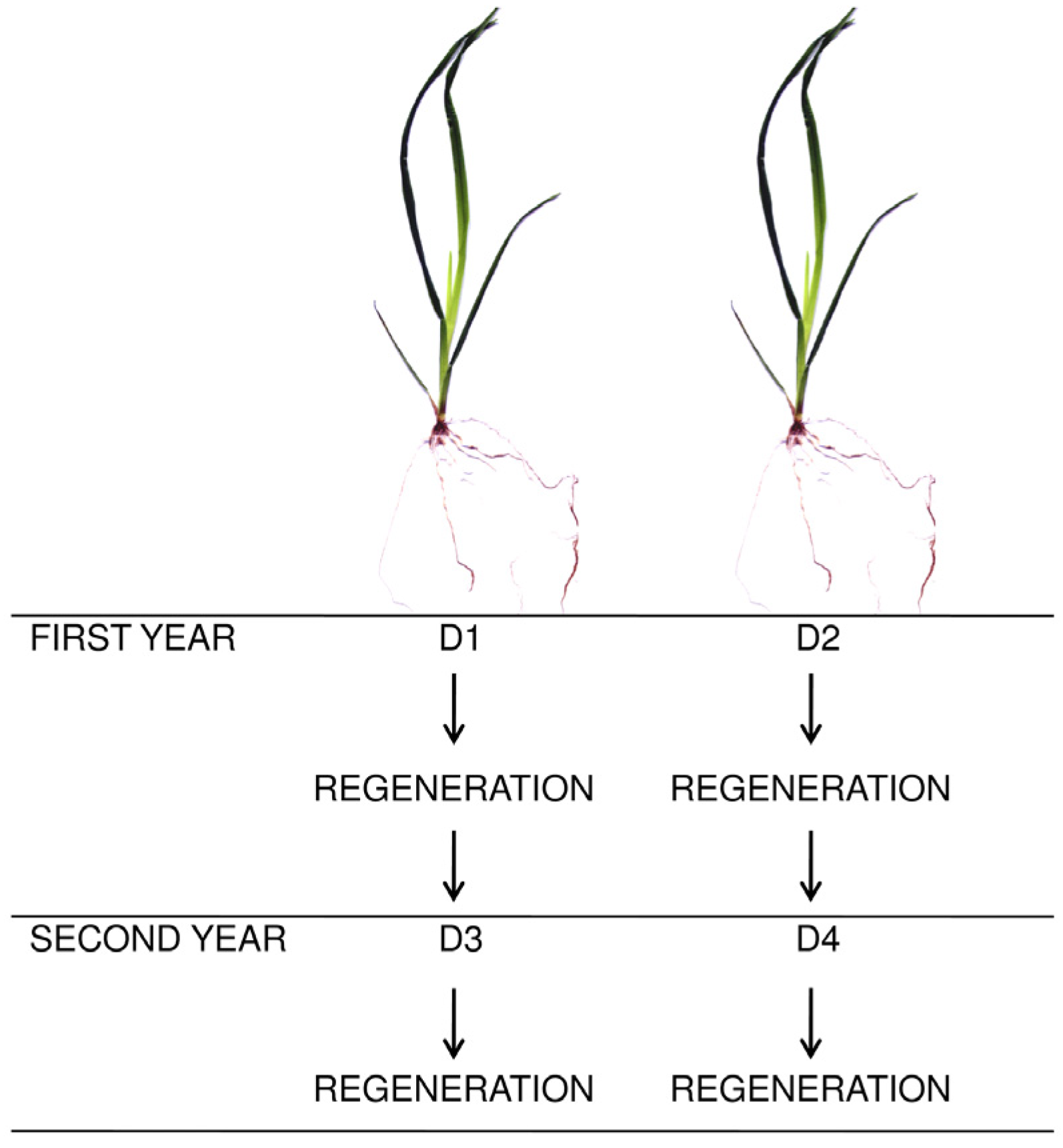
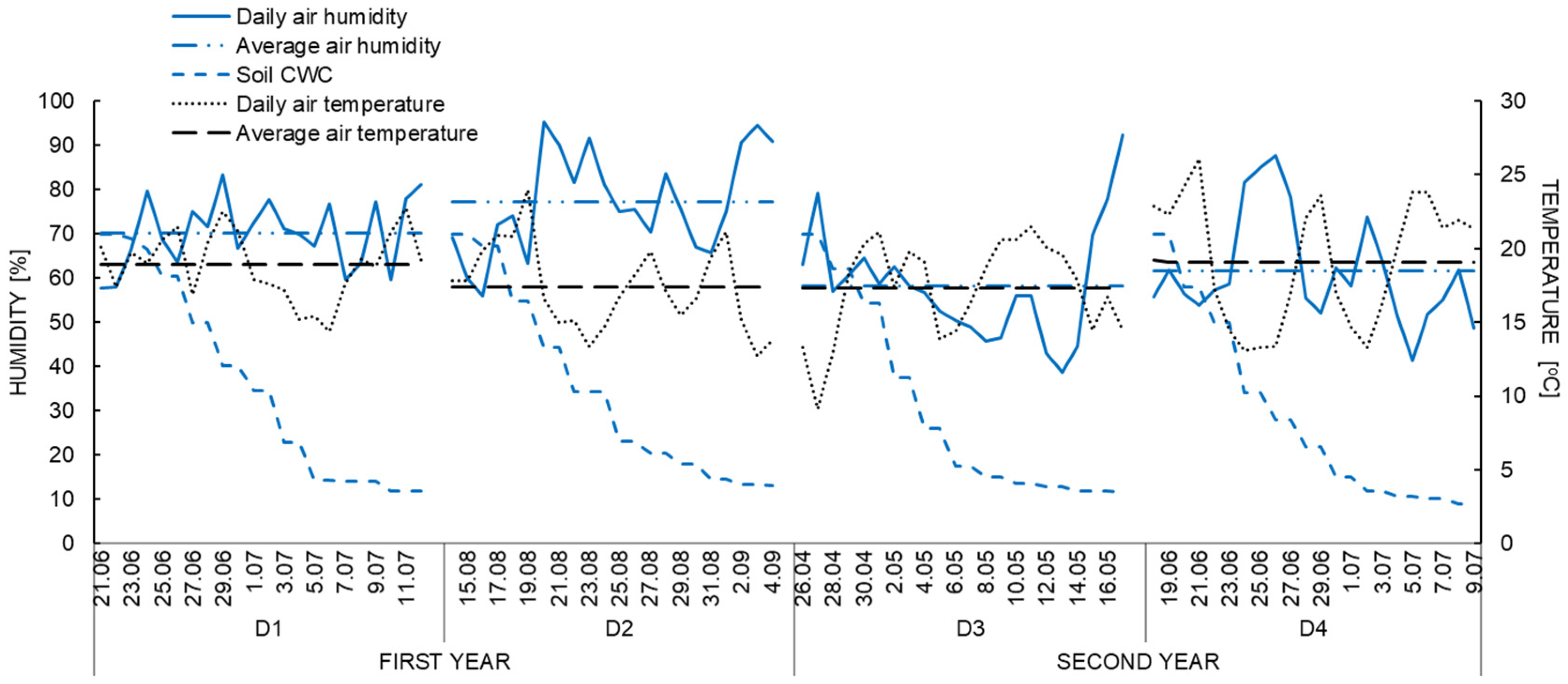
2.1. Experimental Design and Management
2.2. Methods and Measurements
2.3. Statistical Analysis
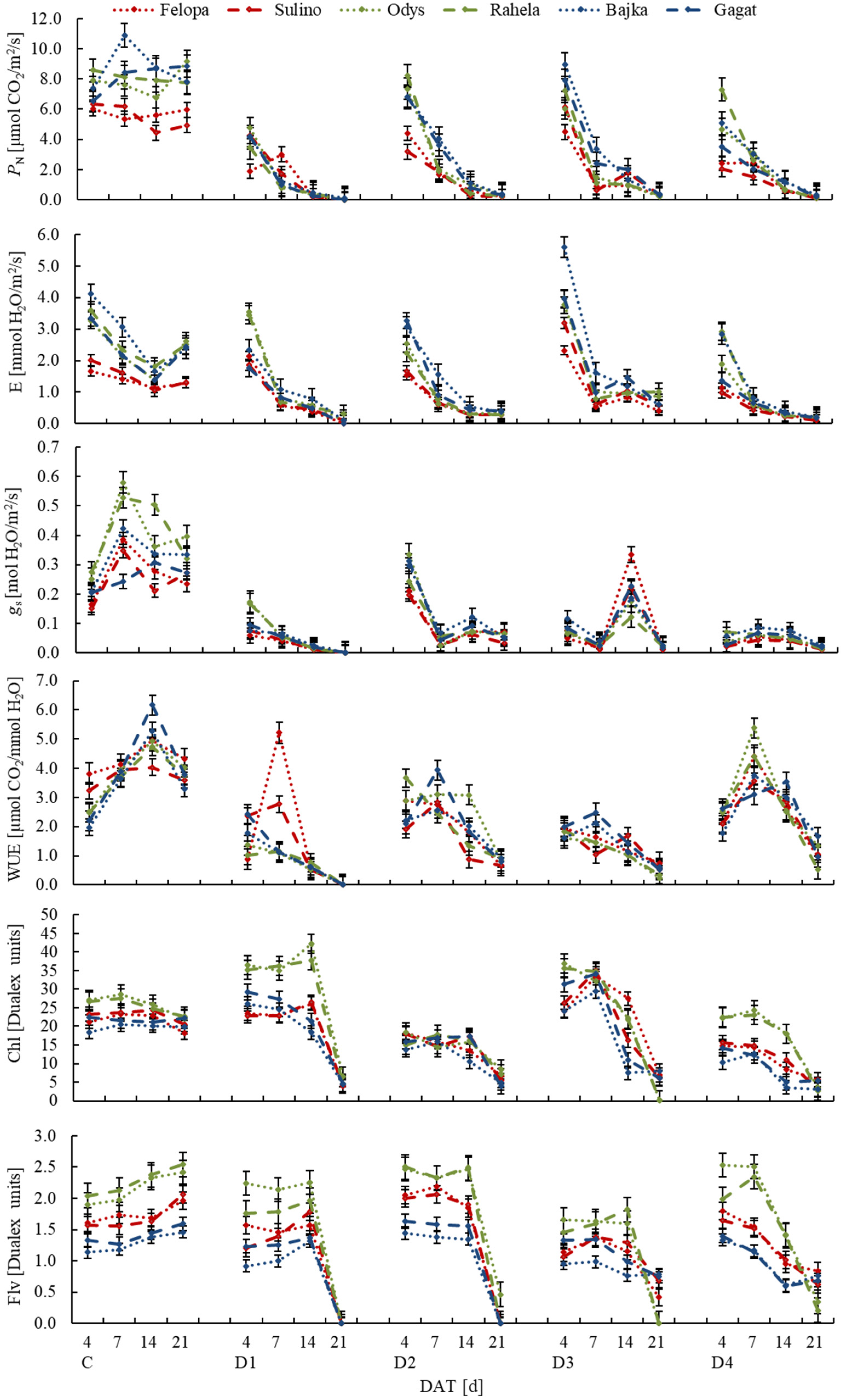
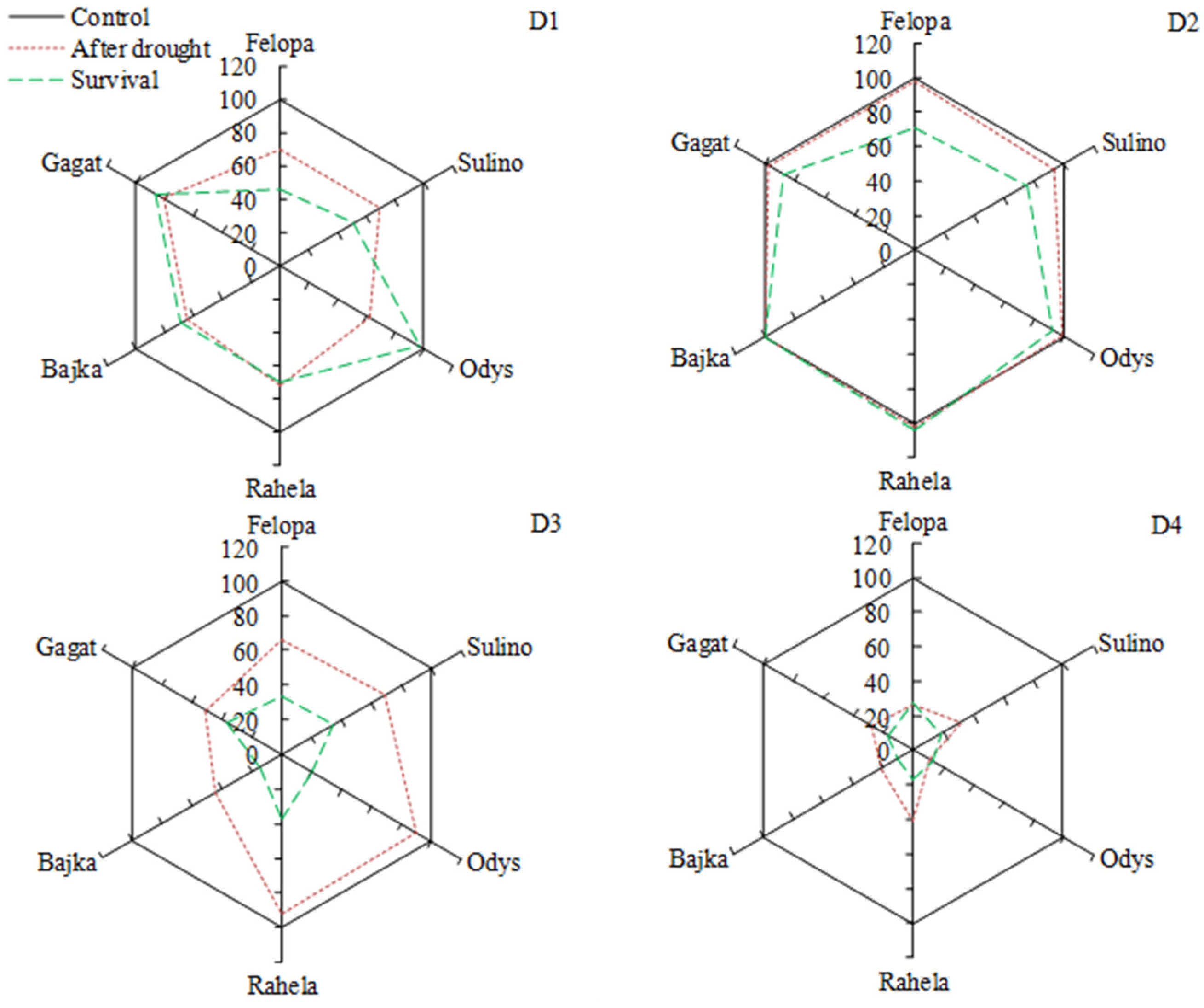
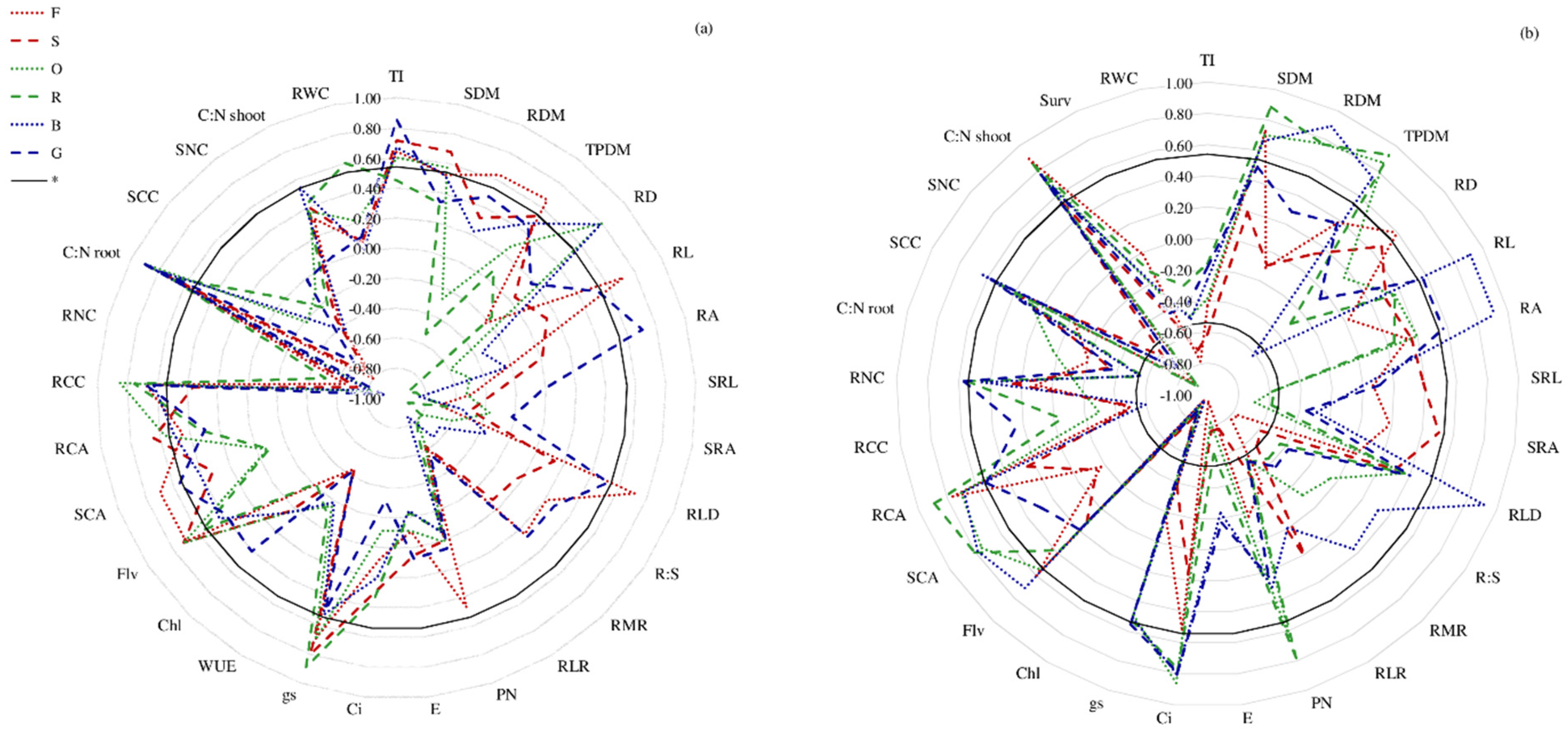
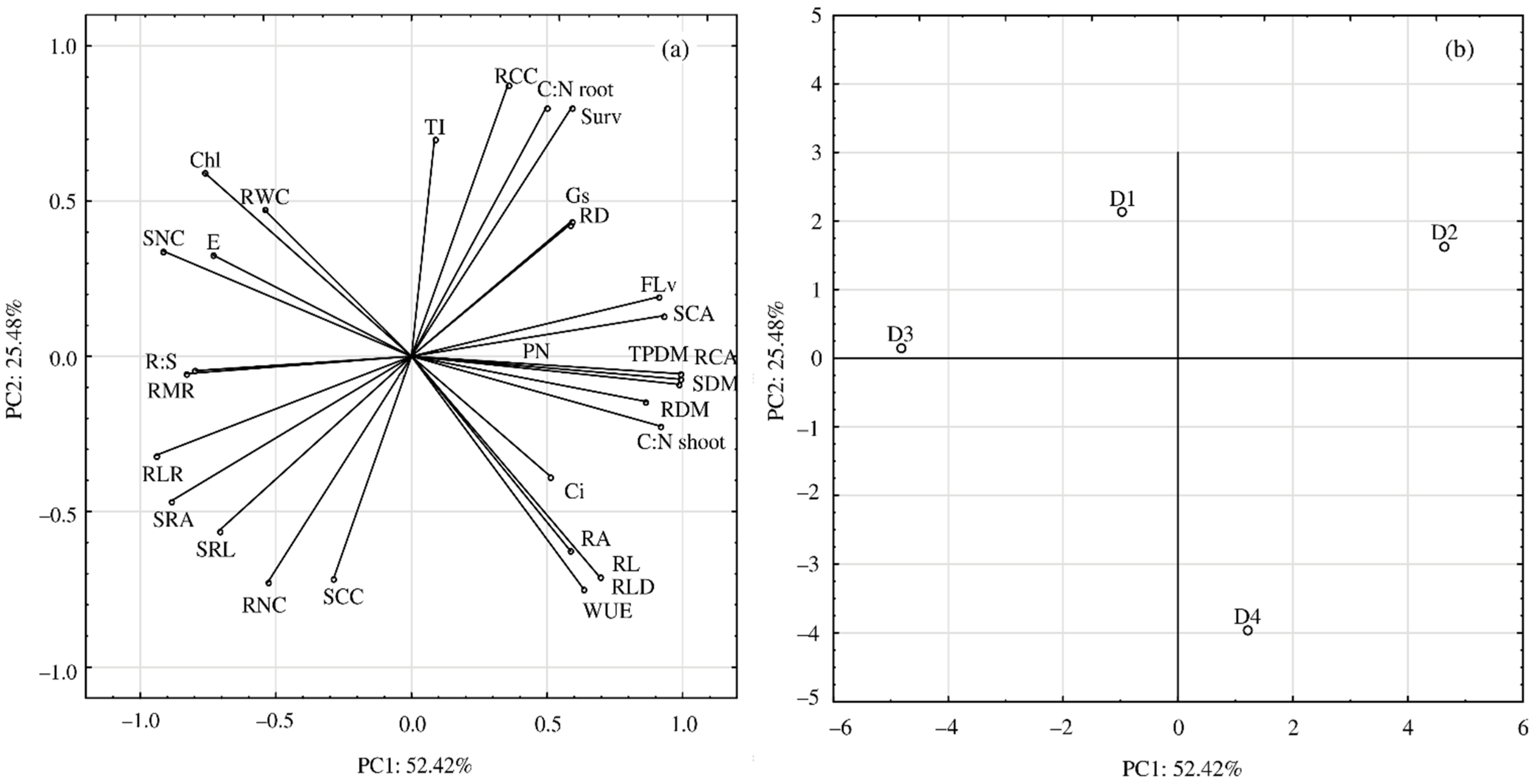
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Łabędzki, L.; Bąk, B. Monitoring and forecasting the course and impact of water deficit in rural areas. Infrastruct. Ecol. Rural Areas 2013, 2, 65–76. (In Polish) [Google Scholar]
- Robertson, T.R.; Bell, C.W.; Zak, J.C.; Tissue, D.T. Precipitation timing and magnitude differentially affect aboveground annual net primary productivity in three perennial species in a Chihuahuan Desert grassland. New Phytol. 2009, 181, 230–242. [Google Scholar] [CrossRef]
- Kipling, P.; Virkajärvi, R.P.; Breitsameter, L.; Curnel, Y.; De Swaef, T.; Gustavsson, A.M.; Hennart, S.; Höglind, M.; Järvenranta, K.; Minet, J.; et al. Key challenges and priorities for modelling European grasslands under climate change. Total Environ. 2016, 566–567, 851–864. [Google Scholar] [CrossRef] [Green Version]
- Marshall, A.H.; Collins, R.P.; Humphreys, M.W.; Scullion, J. A new emphasis on root traits for perennial grass and legume varieties with environmental and ecological benefits. Food Energy Secur. 2016, 5, 26–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosmala, A.; Perlikowski, D.; Pawłowicz, I.; Rapacz, M. Changes in the chloroplast proteome following water deficit and subsequent watering in a high- and a low-drought-tolerant genotype of Festuca Arundinacea. J. Exp. Bot. 2012, 63, 6161–6172. [Google Scholar] [CrossRef] [Green Version]
- Bothe, A.; Westermeier, P.; Wosnitza, A.; Willner, E.; Schum, A.; Dehmer, K.J.; Hartmann, S. Drought tolerance in perennial ryegrass (Lolium perenne L.) as assessed by two contrasting phenotyping systems. J. Agron. Crop. Sci. 2018, 204, 375–389. [Google Scholar] [CrossRef]
- Ghesquiere, M.; Humphreys, M.W.; Zwierzykowski, Z. Festulolium. In Fodder Crops and Amenity Grasses; Boller, B., Posselt, U.K., Veronesi, F., Eds.; Springer: New York, NY, USA, 2010; pp. 293–315. [Google Scholar]
- Yamada, T.J.; Forster, W.; Humphreys, M.W.; Takamizo, T. Genetics and molecular breeding in Lolium/Festuca grass species complex. Grass. Sci. 2005, 51, 89–106. [Google Scholar] [CrossRef]
- Turner, N.C.; Wright, G.C.; Siddique, K.H.M. Adaptation of grain legumes (pulses) to water-limited environments. Adv. Agron. 2001, 71, 123–231. [Google Scholar]
- Hura, T.; Hura, K.; Grzesiak, M.; Rzepka, A. Effect of long-term drought stress on leaf gas exchange and fluorescence parameters in C3 and C4 plants. Acta Physiol. Plant 2007, 29, 103–113. [Google Scholar] [CrossRef]
- Zlatev, Z.; Lidon, F.C. An overview on drought induced changes in plant growth, water relations and photosynthesis. Emir. J. Food Agric. 2012, 24, 57–72. [Google Scholar]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef] [Green Version]
- Munné-Bosch, S.; Alegre, L. Cross-stress tolerance and stress “memory” in plants: An integrated view. Env. Exp. Bot. 2013, 94, 1–2. [Google Scholar] [CrossRef]
- Nosalewicz, A.; Siecińska, J.; Śmiech, M.; Nosalewicz, M.; Wiącek, D.; Pecio, A.; Wach, D. Transgenerational effects of temporal drought stress on spring barley morphology and functioning. Environ. Exp. Bot. 2016, 131, 120–127. [Google Scholar] [CrossRef]
- Verslues, P.E.; Agarwal, M.; Katiyar-Agarwal, S.; Zhu, J.; Zhu, J.K. Methods and concepts in quantifying resistance to drought, salt and freezing, abiotic stresses that affect plant water status. Plant J. 2006, 45, 523–539. [Google Scholar] [CrossRef] [PubMed]
- Deak, K.I.; Malamy, J. Osmotic regulation of root system architecture. Plant J. 2005, 43, 17–28. [Google Scholar] [CrossRef]
- Basu, S.; Ramegowda, V.; Kumar, A.; Pereira, A. Plant adaptation to drought stress [version 1; peer review: 3 approved]. F1000 Res. 2016, 5, 1554. [Google Scholar] [CrossRef]
- FAO. World Reference Base for Soil Resources (WRBSR) 2014. In International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Resources Report No. 106; FAO: Rome, Italy, 2015. [Google Scholar]
- Turner, N.C. Techniques and experimental approaches for the measurement of plant water status. Plant Soil 1981, 58, 339–366. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [Green Version]
- Dąbrowski, P.; Baczewska-Dąbrowska, A.H.; Kalaji, H.M.; Goltsev, V.; Paunov, M.; Rapacz, M.; Wójcik-Jagła, M.; Pawluśkiewicz, B.; Bąba, W.; Brestic, M. Exploration of chlorophyll a fluorescence and plant gas exchange parameters as indicators of drought tolerance in Perennial Ryegrass. Sensors 2019, 19, 2736. [Google Scholar] [CrossRef] [Green Version]
- Lawlor, D.W.; Cornic, G. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant Cell Env. 2002, 25, 275–294. [Google Scholar] [CrossRef] [Green Version]
- Hussain, S.; Ulhassan, Z.; Brestic, M.; Zivcak, M.; Zhou, W.; Allakhverdiev, S.I.; Yang, X.; Safdar, M.E.; Yang, W.; Liu, W. Photosynthesis research under climate change. Photosynth Res. 2021, 150, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Vera, U.M.; Siebers, M.; Gray, S.B.; Drag, D.W.; Rosenthal, D.M.; Kimball, B.A. Global warming can negate the expected CO2 stimulation in photosynthesis and productivity for soybean grown in the Midwestern United States. Plant Physiol. 2013, 162, 410–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumari, V.V.; Roy, A.; Vijayan, R.; Banerjee, P.; Verma, V.C.; Nalia, A.; Pramanik, M.; Mukherjee, B.; Ghosh, A.; Reja, M.H.; et al. Drought and heat stress in cool-season food legumes in sub-tropical regions: Consequences, adaptation, and mitigation strategies. Plants 2021, 10, 1038. [Google Scholar] [CrossRef]
- Deikman, J.; Petracek, M.; Heard, J.E. Drought tolerance through biotechnology: Improving translation from the laboratory to farmers’ fields. Curr. Opin. Biotechnol. 2012, 23, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Lipiec, J.; Doussan, C.A.; Nosalewicz, A.; Kondracka, K. Effect of drought and heat stresses on plant growth and yield: A review. Int. Agrophys. 2013, 27, 463–477. [Google Scholar] [CrossRef]
- Croft, H.; Chen, J.M.; Luo, X.; Bartlett, P.; Chen, B.; Staebler, R.M. Leaf chlorophyll content as a proxy for leaf photosynthetic capacity. Glob. Chang. Biol. 2017, 23, 3513–3524. [Google Scholar] [CrossRef]
- Loka, D.; Harper, J.; Humphreys, M.; Gasior, D.; Gwynn-Jones, D.; Scullion, J.; Doonan, J.; Kingston-Smith, A.; Dodd, R.; Wang, J.; et al. Impacts of abiotic stresses on the physiology and metabolism of cool-season grasses: A review. Food Energy Secur. 2018, 8, e00152. [Google Scholar] [CrossRef] [Green Version]
- Li, R.H.; Guo, P.G.; Michael, B.; Stefania, G.; Salvatore, C. Evaluation of chlorophyll content and fluorescence parameters as indicators of drought tolerance in barley. Agric. Sci. China 2006, 5, 751–757. [Google Scholar] [CrossRef]
- Staniak, M.; Bojarszczuk, J.; Kraska, P.; Kwiatkowski, C.; Harasim, E. Prolonged drought stress induced changes in yield and physiological processes of Trifolium repens and Festulolium braunii. Biol. Plant. 2020, 64, 701–709. [Google Scholar] [CrossRef]
- Pappi, P.; Nikoloudakis, N.; Fanourakis, D.; Zambounis, A.; Delis, C.; Tsaniklidis, G. Differential Triggering of the Phenylpropanoid Biosynthetic Pathway Key Genes Transcription upon Cold Stress and Viral Infection in Tomato Leaves. Horticulturae 2021, 7, 448. [Google Scholar] [CrossRef]
- Basu, S.; Roychoudhury, A.; Saha, P.P.; Sengupta, D.N. Differential antioxidative responses of indica rice cultivars to drought stress. Plant Growth Regul. 2010, 60, 51–59. [Google Scholar] [CrossRef]
- Ma, D.; Sun, D.; Wang, C.; Li, Y.; Guo, T. Expression of flavonoid biosynthesis genes and accumulation of flavonoid in wheat leaves in response to drought stress. Plant Physiol. Biochem. 2014, 80, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Bolinder, M.A.; Janzen, H.H.; Gregorich, E.G.; Angers, D.A.; VandenBygaart, A.J. An approach for estimating net primary productivity and annual carbon inputs to soil for common agricultural crops in Canada. Agric. Ecosyst. Environ. 2007, 118, 29–42. [Google Scholar] [CrossRef]
- Raven, J.A.; Handley, L.L.; Andrews, M. Global aspects of C/N interactions determining plant-environment interactions. J. Exp. Bot 2004, 55, 11–25. [Google Scholar] [CrossRef]
- Nio, S.A.; Cawthray, G.R.; Wade, L.J.; Colmer, T.D. Pattern of solutes accumulated during leaf osmotic adjustment as related to duration of water deficit for wheat at the reproductive stage. Plant Physiol. Biochem. 2011, 49, 1126–1137. [Google Scholar] [CrossRef]
- Reguera, M.; Peleg, Z.; Abdel-Tawab, Y.M.; Tumimbang, E.B.; Delatorre, C.A.; Blumwald, E. Stress-induced cytokinin synthesis increases drought tolerance through the coordinated regulation of carbon and nitrogen assimilation in rice. Plant Physiol. 2013, 163, 1609–1622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leport, L.; Turner, N.C.; French, R.J.; Barr, M.D.; Duda, R.; Davies, S.L. Physiological responses of chickpea genotypes to terminal drought in a Mediterranean-type environment. Eur. J. Agron. 2006, 11, 279–291. [Google Scholar] [CrossRef]
- Kahmen, A.; Perner, J.; Buchmann, N. Diversity-dependent productivity in semi-natural grasslands following climate perturbations. Funct. Ecol. 2005, 19, 594–601. [Google Scholar] [CrossRef]
- Xu, W.; Cui, K.; Xu, A.; Nie, L.; Huang, J.; Peng, S. Drought stress condition increases root to shoot ratio via alteration of carbohydrate partitioning and enzymatic activity in rice seedlings. Acta Physiol. Plant. 2015, 37, 9. [Google Scholar] [CrossRef]
- Nosalewicz, A.; Siecińska, J.; Kondracka, K.; Nosalewicz, M. The functioning of Festuca arundinacea and Lolium perenne under drought is improved to a different extend by the previous exposure to water deficit. Environ. Exp. Bot. 2018, 156, 271–278. [Google Scholar] [CrossRef]
- Liu, Y.; Li, P.; Xu, G.C.; Xiao, L.; Ren, Z.P.; Li, Z.B. Growth, morphological, and physiological responses to drought stress in Bothriochloa ischaemum. Front. Plant Sci. 2017, 8, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anjum, S.A.; Xiao-yu Xie, X.; Wang, L.; Saleem, M.F.; Man, C.; Le, W. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar] [CrossRef]
- Taheri-Garavand, A.; Rezaei Nejad, A.; Fanourakis, D.; Fatahi, S.; Majd, M.A. Employment of artificial neural networks for non-invasive estimation of leaf water status using color features: A case study in Spathiphyllum wallisii. Acta Physiol Plant 2021, 43, 78. [Google Scholar] [CrossRef]
- Blum, A. Effective use of water (EUW) and not water-use efficiency (WUE) is the target of crop yield improvement under drought stress. Field Crop. Res. 2009, 112, 119–123. [Google Scholar] [CrossRef]
- Abdollahi, B.M.; Saeidnia, F.; Majidi, M.M.; Mirlohi, A. Growth traits associated with drought survival, recovery and persistence of cocksfoot (Dactylis glomerata) under prolonged drought treatments. Crop. Pasture Sci. 2019, 70, 85–94. [Google Scholar]
- Borawska-Jarmułowicz, B.; Mastalerczuk, G.; Dąbrowski, P.; Kalaji, H.M.; Wytrążek, K. Improving tolerance in seedlings of some Polish varieties of Dactylis glomerata to water deficit by application of simulated drought during seed germination. Photosynth 2020, 58, 355–363. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Description and Unit | Trait | Description and Unit |
|---|---|---|---|
| Chl | Content of chlorophyll (Dualex unit) | RMR | Root mass ratio—root dry mass to total plant dry mass (g/g) |
| Ci | Sub-stomatal CO2 concentration (µmol CO2/mol air) | RNC | Root nitrogen content (g/kg DM) |
| E | Transpiration rate (mmol H2O/m2/s) | Root C:N | Root carbon to nitrogen ratio |
| Flv | Content of flavonols (Dualex unit) | RWC | Relative water content (%) |
| gs | Stomatal conductance (mol H2O/m2/s) | SCA | Shoot carbon accumulation (kg/plant) |
| PN | Net photosynthetic rate (μmol CO2/m2/s) | SCC | Shoot carbon content (g/kg DM) |
| R:S | Root to shoot ratio—root dry mass to shoot dry mass (g/g) | SDM | Shoot dry mass (g/plant) |
| RA | Root area (cm2/plant) | Shoot C:N | Shoot carbon to nitrogen ratio |
| RCA | Root carbon accumulation (kg/plant) | SNC | Shoot nitrogen content (g/kg DM) |
| RCC | Root carbon content (g/kg DM) | SRA | Specific root area (m2/g) |
| RD | Root diameter (mm) | SRL | Specific root length (m/g) |
| RDM | Root dry mass (g/plant) | Surv | Survival of plants (%) |
| RL | Total root length (m/plant) | TI | Tillering intensity—the number of shoots per plant (no./plant) |
| RLD | Root length density (cm/cm3) | TPDM | Total plant dry mass (g/plant) |
| RLR | Root length ratio—root length to total plant dry mass (m/g) | WUE | Water use efficiency (µmol CO2/mmol H2O) |
| Factor | Features | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PN | E | Ci | gs | WUE | Chl | Flv | SCC | SNC | Shoot C:N | SCA | RCC | RNC | Root C:N | RCA | ||
| year | ||||||||||||||||
| 1st | 4.11 a | 1.44 a | 564 b | 0.27 b | 2.76 a | 20.3 a | 1.54 a | 400 a | 16.5 a | 28.9 b | 2.22 a | 377 b | 9.7 a | 39.0 b | 2.54 b | |
| 2nd | 5.28 b | 1.79 b | 525 a | 0.14 a | 2.85 a | 19.9 a | 1.49 a | 408 b | 20.4 b | 22.1 a | 2.30 a | 366 a | 14.4 b | 26.0 a | 2.40 a | |
| water conditions | ||||||||||||||||
| C | 7.33 b | 2.15 c | 567 b | 0.32 c | 3.85 c | 22.9 b | 1.76 c | 402 b | 14.8 b | 28.7 b | 3.08 c | 371 b | 10.9 b | 34.8 b | 3.15 e | |
| D1 | 1.38 a | 0.95 a | 429 a | 0.04 a | 1.08 a | 22.6 b | 1.17 a | 397 a | 27.0 c | 14.8 a | 1.96 b | 375 b | 9.4 a | 39.8 c | 1.82 b | |
| D2 | 2.40 a | 0.99 a | 584 b | 0.19 b | 2.04 b | 13.4 a | 1.50 b | 403 b | 11.1 a | 37.2 c | 2.17 b | 396 c | 10.2 b | 38.9 c | 2.88 d | |
| D3 | 2.52 a | 1.62 b | 523 b | 0.08 a | 1.31 a | 21.3 b | 1.09 a | 409 c | 29.1 d | 14.4 a | 1.34 a | 369 b | 15.8 d | 23.5 a | 1.44 a | |
| D4 | 1.91 a | 0.73 a | 551 b | 0.04 a | 2.57 b | 12.0 a | 1.26 ab | 408 c | 14.0 b | 29.4 b | 1.92 b | 345 a | 15.0 c | 23.2 a | 2.36 c | |
| species | ||||||||||||||||
| Fb | 3.57 a | 1.11 a | 415 a | 0.17 a | 2.90 a | 19.3 b | 1.48 b | 400 a | 19.0 b | 24.2 a | 2.28 b | 363 a | 12.6 b | 30.9 a | 2.59 b | |
| Fa | 5.07 b | 1.81 b | 474 b | 0.25 b | 2.74 a | 23.2 c | 1.88 c | 407 c | 17.3 a | 27.6 b | 2.50 c | 382 b | 11.6 a | 34.1 c | 2.49 b | |
| Lp | 5.43 b | 1.92 b | 745 c | 0.20 a | 2.77 a | 17.8 a | 1.17 a | 404 b | 19.0 b | 24.8 a | 1.99 a | 368 a | 11.9 a | 32.5 b | 2.32 a | |
| cultivars | ||||||||||||||||
| Fb | Felopa | 3.62 a | 1.07 a | 419 a | 0.18 a | 3.11 a | 19.0 ab | 1.49 b | 398 a | 18.9 b | 24.5 ab | 2.13 b | 358 a | 12.7 c | 30.1 a | 2.60 b |
| Fb | Sulino | 3.51 a | 1.15 a | 410 a | 0.16 a | 2.69 a | 19.6 b | 1.46 b | 402 b | 19.2 b | 24.0 a | 2.44 c | 367 b | 12.4 bc | 31.7 b | 2.58 b |
| Fa | Odys | 4.99 b | 1.75 b | 475 a | 0.25 b | 2.83 a | 23.5 c | 1.89 c | 407 c | 17.4 a | 28.1 c | 2.69 d | 388 d | 11.7 a | 34.8 d | 2.45 ab |
| Fa | Rahela | 5.16 b | 1.87 b | 472 a | 0.25 b | 2.64 a | 23.0 c | 1.87 c | 406 c | 17.3 a | 27.0 c | 2.31 bc | 376 bc | 11.6 a | 33.5 cd | 2.54 b |
| Lp | Bajka | 5.66 b | 2.12 b | 831 c | 0.21 ab | 2.62 a | 16.6 a | 1.10 a | 407 c | 18.8 b | 25.2 b | 1.85 a | 380 c | 12.0 ab | 33.0 bc | 2.18 a |
| Lp | Gagat | 5.21 b | 1.72 b | 659 b | 0.18 a | 2.93 a | 18.9 ab | 1.23 a | 401 ab | 19.1 b | 24.4 ab | 2.13 b | 357 a | 11.8 a | 32.0 b | 2.46 ab |
| Factor | Features | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TI | SDM | RDM | TPDM | RD | RL | RA | SRL | SRA | RLD | R:S | RMR | RLR | RWC | ||
| year | |||||||||||||||
| 1st | 26.0 a | 6.42 b | 5.97 a | 15.56 b | 0.48 b | 346 a | 4952 a | 60.18 a | 869 a | 4.74 a | 0.63 a | 0.38 a | 3.24 a | 48.25 a | |
| 2nd | 27.3 a | 5.94 a | 6.24 b | 15.06 a | 0.44 a | 408 b | 5597 b | 70.87 b | 974 b | 5.59 b | 0.70 b | 0.41 b | 3.86 b | 47.75 a | |
| water conditions | |||||||||||||||
| C | 31.9 b | 7.92 e | 8.27 c | 19.67 e | 0.47 c | 479 d | 6693 d | 61.53 a | 861 ab | 6.56 d | 0.72 c | 0.41 c | 3.45 b | 81.40 e | |
| D1 | 19.4 a | 4.62 b | 5.29 b | 12.69 b | 0.48 c | 299 a | 4573 b | 60.24 a | 937 bc | 4.09 a | 0.71 c | 0.41 c | 3.66 b | 11.45 b | |
| D2 | 31.9 b | 7.22 d | 5.55 b | 16.22 d | 0.46 bc | 340 b | 4524 b | 59.79 a | 803 a | 4.65 b | 0.52 a | 0.34 a | 2.77 a | 15.93 c | |
| D3 | 18.0 a | 3.56 a | 3.64 a | 9.13 a | 0.43 a | 277 a | 3785 a | 78.49 c | 1071 d | 3.80 a | 0.67 bc | 0.40 c | 4.25 c | 25.52 d | |
| D4 | 16.3 a | 5.84 c | 5.61 b | 14.48 c | 0.45 b | 386 c | 5379 c | 71.56 b | 998 cd | 5.30 c | 0.64 b | 0.38 b | 3.73 bc | 5.51 a | |
| species | |||||||||||||||
| Fb | 25.2 b | 6.55 c | 6.34 b | 15.94 b | 0.44 a | 401 b | 5429 b | 65.62 b | 884 b | 5.50 b | 0.66 b | 0.40 b | 3.45 b | 47.61 a | |
| Fa | 20.7 a | 6.18 b | 6.55 b | 15.58 b | 0.49 b | 264 a | 3920 a | 42.28 a | 633 a | 3.61 a | 0.73 b | 0.42 c | 2.62 a | 48.73 a | |
| Lp | 34.1 c | 5.80 a | 5.42 a | 14.42 a | 0.44 a | 465 c | 6474 c | 88.67 c | 1248 c | 6.37 c | 0.60 a | 0.37 a | 4.58 c | 47.66 a | |
| cultivars | |||||||||||||||
| Fb | Felopa | 24.2 bc | 6.62 d | 6.00 b | 15.63 bc | 0.45 a | 389 b | 5268 b | 67.79 c | 913 b | 5.34 b | 0.62 b | 0.38 b | 3.42 b | 48.39 ab |
| Fb | Sulino | 26.3 cd | 6.48 cd | 6.69 c | 16.24 c | 0.44 a | 413 b | 5589 b | 63.45 c | 856 b | 5.67 b | 0.71 c | 0.41 c | 3.48 b | 46.83 a |
| Fa | Odys | 21.3 ab | 6.07 b | 6.97 c | 15.91 c | 0.50 c | 255 a | 3835 a | 39.07 a | 590 a | 3.49 a | 0.78 d | 0.43 d | 2.53 a | 48.53 ab |
| Fa | Rahela | 20.0 a | 6.30 bc | 6.14 b | 15.24 b | 0.48 b | 272 a | 4006 a | 45.49 b | 676 a | 3.73 a | 0.67 bc | 0.40 c | 2.72 a | 48.93 b |
| Lp | Bajka | 39.2 e | 5.41 a | 4.92 a | 13.76 a | 0.45 a | 476 c | 6742 d | 97.59 e | 1397 d | 6.52 c | 0.56 a | 0.36 a | 5.00 d | 47.26 ab |
| Lp | Gagat | 29.0 d | 6.20 bc | 5.92 b | 15.07 b | 0.44 a | 454 c | 6206 c | 79.75 d | 1100 c | 6.22 c | 0.63 b | 0.38 b | 4.16 c | 48.05 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mastalerczuk, G.; Borawska-Jarmułowicz, B. Physiological and Morphometric Response of Forage Grass Species and Their Biomass Distribution Depending on the Term and Frequency of Water Deficiency. Agronomy 2021, 11, 2471. https://doi.org/10.3390/agronomy11122471
Mastalerczuk G, Borawska-Jarmułowicz B. Physiological and Morphometric Response of Forage Grass Species and Their Biomass Distribution Depending on the Term and Frequency of Water Deficiency. Agronomy. 2021; 11(12):2471. https://doi.org/10.3390/agronomy11122471
Chicago/Turabian StyleMastalerczuk, Grażyna, and Barbara Borawska-Jarmułowicz. 2021. "Physiological and Morphometric Response of Forage Grass Species and Their Biomass Distribution Depending on the Term and Frequency of Water Deficiency" Agronomy 11, no. 12: 2471. https://doi.org/10.3390/agronomy11122471
APA StyleMastalerczuk, G., & Borawska-Jarmułowicz, B. (2021). Physiological and Morphometric Response of Forage Grass Species and Their Biomass Distribution Depending on the Term and Frequency of Water Deficiency. Agronomy, 11(12), 2471. https://doi.org/10.3390/agronomy11122471