Effects of Plant Growth Regulators and N Rate on the Morphological Traits of Generative Tillers and Seed Yields of Chewings and Strong Creeping Red Fescue



Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Experimental Design
2.3. Crop Management
2.4. Measurement
2.5. Statistical Analysis
3. Results
3.1. Morphological Traits of Generative Tillers
3.2. Seed Yield and Yield Components
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bitarafan, Z.; Rasmussen, J.; Westergaard, J.C.; Andreasen, C. Seed yield and lodging assessment in red fescue (Festuca rubra L.) sprayed with trinexapac-ethyl. Agronomy 2019, 9, 617. [Google Scholar] [CrossRef]
- Braun, R.; Patton, A.J.; Watkins, E.; Koch, P.L.; Anderson, N.P.; Bonos, S.A.; Brilman, L.A. Fine fescues: A review of the species, their improvement, production, establishment, and management. Crop Sci. 2020, 60, 1142–1187. [Google Scholar] [CrossRef]
- Duru, M.; Cruz, P.; Magda, D. Using plant traits to compare sward structure and composition of grass species cross environment al gradients. Appl. Veg. Sci. 2004, 7, 11–18. [Google Scholar] [CrossRef]
- Stukonis, V.; Juzenas, S.; Ceseviciene, J.; Norkeviciene, E. Assesment of morfo-anatomical traits of red fescue (Festuca rubra L.) germplasm differing in origin. Zemdirbyste 2015, 102, 437–442. [Google Scholar] [CrossRef][Green Version]
- De Vazquez Aldana, B.R.; Gundel, P.E.; Garcia Criado, B.; Garcia Ciudad, A.; Garcia Sanchez, A.; Zabalgogeacoa, I. Germination response of endophytic Festuca rubra seeds in presence of arsenic. Grass Forage Sci. 2013, 69, 462–469. [Google Scholar] [CrossRef]
- Bertin, C.; Senesac, A.F.; Rossi, F.S.; DiTommaso, A.; Westo, L.A. Evaluation of selected fine-leaf fescue cultivars for their turfgrass quality and weed suppressive ability in field settings. HortTechnology 2009, 19, 660–668. [Google Scholar] [CrossRef]
- Szczepanek, M.; Onofri, A. Chewing, strong and slender creeping red fescue response to sowing time and method. Crop Sci. 2013, 53, 2613–2622. [Google Scholar] [CrossRef]
- Boelt, B. Undersowing Poa pratensis L., Festuca rubra L., Festuca pratensis Huds., Dactylis glomerata L., Lolium perenne L., for seed production in five cover crops. III. The effect of autumn applied nitrogen on seed yield of the undersown grasses. J. Appl. Seed Prod. 1997, 15, 55–61. [Google Scholar]
- Macháč, R. Effects of trinexapac-ethyl (Moddus) on seed yields and its quality of eleven temperate grass species. In Breeding Strategies for Sustainable Forage and Turf Grass Improvement; Barth, S., Milbourne, D., Eds.; Springer: Berlin, Germany, 2013; pp. 359–363. [Google Scholar]
- Baczyński, M.; Szczepanek, M.; Bech, A. Seed quality of red fescue (Festuca rubra ssp. rubra and F. r. ssp. commutata) depending on nitrogen fertilization and plant growth regulators. In Proceedings of the 14th Scientific and Technical Seminar on Seed and Seedlings, Prague, Czech Republic, 7 February 2019; pp. 186–192. [Google Scholar]
- Chynoweth, R.J.; Moot, D.J. Seed growth of three perennial ryegrass cultivars sown on two dates and treated with trinexapac ethyl straw shortener. Grass Forage Sci. 2016, 72. [Google Scholar] [CrossRef]
- Griffith, S.M. Changes in dry matter, carbohydrate and seed yield resulting from lodging in three temperate grass species. Ann. Bot. 2000, 85, 675–680. [Google Scholar] [CrossRef]
- Haldrup, C. Growth regulation, fungicides and nitrogen interaction in seed crop production. In Proceedings of the 6th International Herbage Seed Conference, Giennestad, Norway, 18–20 June 2007; pp. 211–2013. [Google Scholar]
- Young, W.C., III; Silberstein, T.B.; Chastain, T.G.; Garbacik, C.J. Response of creeping red fescue (Festuca rubra L.) and perennial ryegrass (Lolium perenne L.) to spring nitrogen fertility and plant growth regulator applications in Oregon. In Proceedings of the 6th International Herbage Seed Conference, Giennestad, Norway, 18–20 June 2007; pp. 201–205. [Google Scholar]
- Rademacher, W. Growth retardants: Effect on gibberellin biosynthesis and metabolic pathways. Annu. Rev. Plant Biol. 2000, 51, 501–531. [Google Scholar] [CrossRef] [PubMed]
- Rajala, A.; Peltonen-Sainio, P.; Onnela, M.; Jackson, M. Effects of applying stem-shortening plant growth regulators to leaves on root elongation by seedlings of wheat, oat and barley: Mediation by ethylene. Plant Growth Regul. 2002, 38, 51–59. [Google Scholar] [CrossRef]
- Borm, G.E.L. Effects of the application rate and time of the growth regulator trinexapac-ethyl in seed crops of Lolium perenne L. in relation to spring nitrogen rate. Field Crops Res. 2008, 105, 182–192. [Google Scholar] [CrossRef]
- Rolston, M.; Chynoweth, R.; Kelly, M.; McCloy, B.; Trethewey, J. Seed yield response of four cocksfoot (Dactylis glomerata L.) cultivars following the application of stem-shortening plant growth regulators. N. Z. J. Agric. Res. 2014, 53, 332–341. [Google Scholar] [CrossRef]
- Rolston, M.; Trethewey, J.; Chynoweth, R.; McCloy, B. Trinexapac-ethyl delays lodging and increases seed yield in perennial ryegrass seed crops. N. Z. J. Agric. Res. 2010, 53, 403–406. [Google Scholar] [CrossRef]
- Chastain, T.; Young, W.C., III; Garbacik, C.J.; Silberstein, T.B. Trinexapac-ethyl rate and application timing effects on seed yield and yield components in tall fescue. Field Crops Res. 2015, 173, 8–13. [Google Scholar] [CrossRef]
- Szczepanek, M.; Stypczyńska, Z.; Dziamski, A.; Wichrowska, D. Above- and below-ground part growth in chewings and strong creeping red fescue grown for seed resulting from retardants and N fertilization. Agronomy 2020, 10, 4. [Google Scholar] [CrossRef]
- Berthet, P.; Gerard, G. A statistical study of microdistribution of oribatei (Acari) Part I. The distribution pattern. Oikos 1965, 16, 214. [Google Scholar] [CrossRef]
- Łomnicki, A. Introduction to Statistics for Naturalists, 4th ed.; PWN: Warsaw, Poland, 2010; p. 280. (In Polish) [Google Scholar]
- Stanisz, A. Easy Course of Statistic Using Statistica PL and Medicine Examples, 1. Basic Statistic; StatSoft Polska: Krakow, Poland, 2006; p. 532. (In Polish) [Google Scholar]
- Leps, J.; Smilauer, P. Multivariate Analysis of Ecological Data Using CANOCO; Cambridge University Press: Cambridge, UK, 2003; p. 283. [Google Scholar]
- Piernik, A. Numerical Methods in Ecology; Wydawnictwo Naukowe UMK: Torun, Poland, 2008; p. 98. (In Polish) [Google Scholar]
- Legendre, P.; Oksanen, J.; ter Braak, C.J.F. Testing the significance of canonical axes in redundancy analysis. Methods Ecol. Evol. Br. Ecol. Soc. 2011, 2, 269–277. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology, 2nd ed.; Elsevier Science BV: Amsterdam, The Netherlands, 1998; p. 853. [Google Scholar]
- Stanisz, A. Easy Course of Statistic Using Statistica PL and Medicine Examples, 2. Linear and Non-Linear Models; StatSoft Polska: Krakow, Poland, 2007; p. 868. (In Polish) [Google Scholar]
- Mathiassen, S.K.; Rabolle, M.; Boelt, B.; Kudsk, P. Factors affecting the activity of Moddus M in red fescue. In Proceedings of the 6th International Herbage Seed Conference, Giennestad, Norway, 18–20 June 2007; pp. 169–171. [Google Scholar]
- Zhang, Y.; Su, S.; Tabori, M.; Yu, J.; Chabot, D.; Baninasab, B.; Wang, X.; Ma, B.; Li, C.; Khanizadeh, S. Effect of selected plant growth regulators on yield and stem height of spring wheat in Ontario. J. Agric. Sci. 2017, 9, 30–42. [Google Scholar] [CrossRef][Green Version]
- Wiersma, J.J.; Dai, J.; Durgan, B.R. Optimum timing and rate of trinexapac-ethyl to reduce lodging in spring wheat. Agron. J. 2011, 103, 864–870. [Google Scholar] [CrossRef]
- Zapiola, M.L.; Chastain, T.G.; Garbacik, C.J.; Young, W.C. Trinexapac-ethyl and burning effects on seed yield components in strong creeping red fescue. Agron. J. 2014, 106, 1371–1378. [Google Scholar] [CrossRef]
- Chastain, T.; Young, W.C., III; Silberstein, T.B.; Garbacik, C.J. Performance of trinexapac-ethyl on Lolium perenne seed crops in diverse lodging environments. Field Crops Res. 2014, 157, 65–70. [Google Scholar] [CrossRef]
- McCann, S.E.; Huang, B. Effects of trinexapac-ethyl foliar application on creeping bentgrass responses to combined drought and heat stress. Crop Sci. 2007, 47, 2121–2128. [Google Scholar] [CrossRef]
- Young, W.C.; Chilcote, D.O.; Youngberg, H.W. Chemical dwarfing and response of cool-season grass seed crops to spring-applied nitrogen. Agron. J. 1999, 91, 344–350. [Google Scholar] [CrossRef]
- Janovszky, J. Effects of production factors on seed yield and yield components of red fescue (Festuca rubra ssp. genuine Hack). 2. Effects of production factors on panicle productivity. Cereal Res. Commun. 1994, 22, 211–217. [Google Scholar]
- Young, W.C.; Chilcote, D.O.; Youngberg, H.W. Split-applied nitrogen and productivity of cool-season grass seed crops. Agron. J. 1999, 91, 339–343. [Google Scholar] [CrossRef]
- Fairey, N.A. Cultivar-specific management for seed production of creeping red fescue. Can. J. Plant Sci. 2006, 86, 1099–1105. [Google Scholar] [CrossRef]
- Fairey, N.A.; Lefkovitch, L.P. Crop density and seed production of creeping red fescue (Festuca rubra L. var. rubra). 1. Yield and plant development. Can. J. Plant Sci. 1996, 76, 291–298. [Google Scholar] [CrossRef]



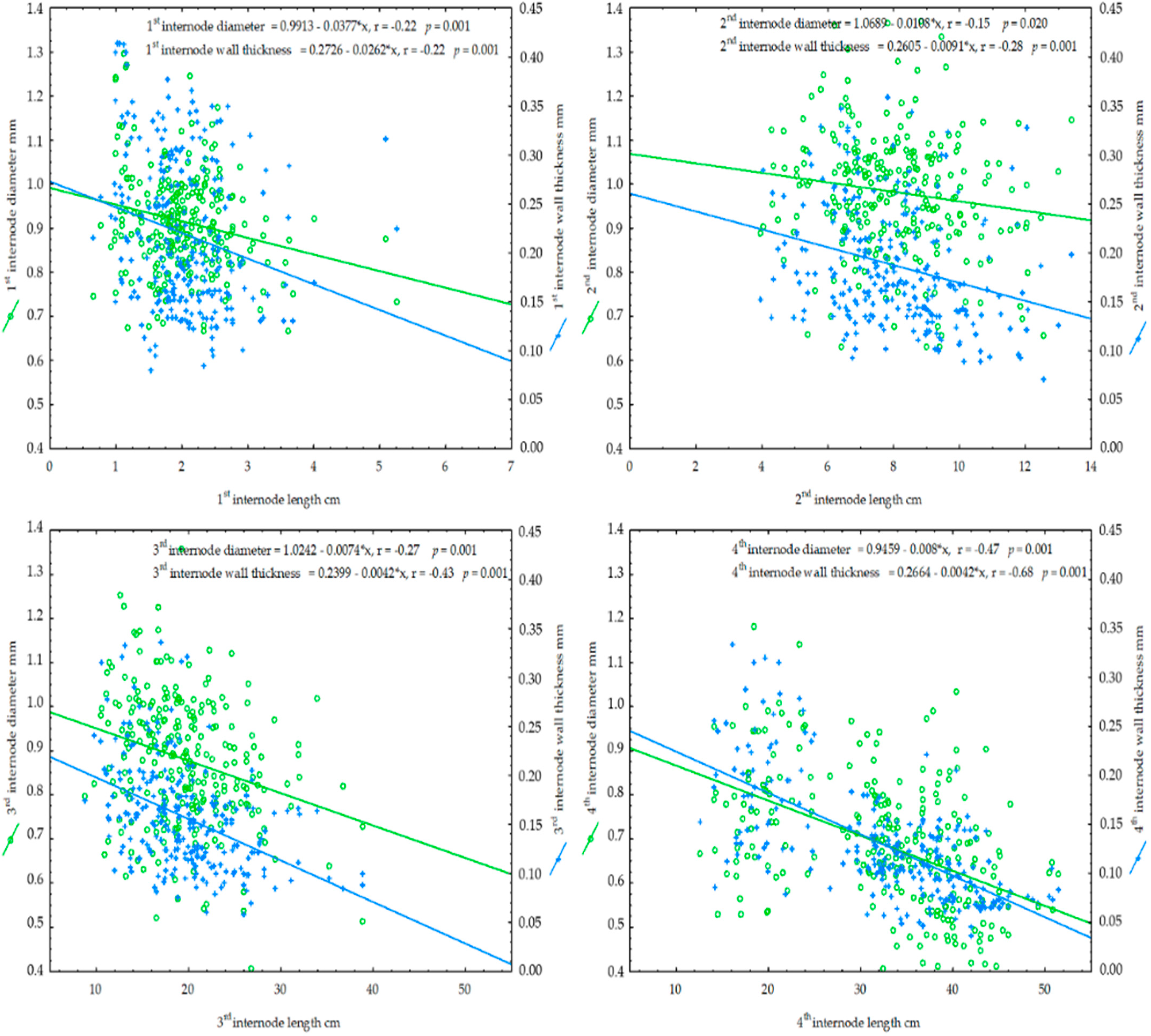{kind=link}
{kind=link}
| Plant Growth Regulator | Cultivar | Nitrogen Rate |
|---|---|---|
| Control- without treatment | cv. Nista strong creeping red fescue Festuca rubra L ssp. rubra | 40 kg ha−1 |
| CCC–chloromequat chloride 1350 a.i. g ha−1, at the beginning of stem elongation stage (BBCH 30–31) | ||
| CCC–chloromequat chloride 625 a.i. g ha−1, at the beginning of stem elongation stage (BBCH 30–31) | ||
| + ET–ethephon 240 a.i. g ha−1, at flag the leaf stage (BBCH 37–39) | cv. Dorosa Chewings red fescue Festuca rubra L ssp. commutata | 70 kg ha−1 |
| CCC–chloromequat chloride 625 a.i. g ha−1, at the beginning of stem elongation stage (BBCH 30–39) | ||
| + TE–trinexapac-ethyl 75 a.i. g ha−1, at the flag leaf stage (BBCH 37–39) |
| Specification | Internode Number | Plant Growth Regulator | Cultivar | N Rate | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | p | df | MS | F | p | df | MS | F | p | df | MS | Residuals | ||
| Internode | 1st | 4.67 | 0.003 | 3 | 1.768 | 21.52 | <0.001 | 1 | 8.141 | 0.10 | 0.751 | 1 | 0.038 | 0.378 |
| length | 2nd | 25.83 | <0.001 | 3 | 64.109 | 12.70 | <0.001 | 1 | 31.531 | 0.01 | 0.942 | 1 | 0.013 | 2.482 |
| cm | 3rd | 4.66 | 0.003 | 3 | 123.362 | 4.35 | 0.038 | 1 | 115.216 | 0.45 | 0.505 | 1 | 11.795 | 26.481 |
| 4th | 0.92 | 0.430 | 3 | 80.189 | 0.83 | 0.363 | 1 | 72.000 | <0.001 | 0.970 | 1 | 0.120 | 86.819 | |
| Internode | 1st | 0.55 | 0.652 | 3 | 0.007 | 4.85 | 0.029 | 1 | 0.062 | 0.53 | 0.467 | 1 | 0.007 | 0.013 |
| diameter | 2nd | 3.19 | 0.024 | 3 | 0.059 | 0.01 | 0.919 | 1 | <0.001 | 0.03 | 0.858 | 1 | <0.001 | 0.019 |
| mm | 3rd | 1.80 | 0.148 | 3 | 0.037 | 3.08 | 0.081 | 1 | 0.063 | 4.21 | 0.041 | 1 | 0.087 | 0.021 |
| 4th | 1.64 | 0.180 | 3 | 0.040 | 0.83 | 0.362 | 1 | 0.020 | 3.70 | 0.055 | 1 | 0.091 | 0.024 | |
| Internode wall | 1st | 0.79 | 0.499 | 3 | 0.004 | 9.49 | 0.002 | 1 | 0.053 | 1.21 | 0.273 | 1 | 0.007 | 0.005 |
| thickness | 2nd | 3.12 | 0.027 | 3 | 0.011 | 1.14 | 0.286 | 1 | 0.004 | 0.24 | 0.625 | 1 | <0.001 | 0.003 |
| mm | 3rd | 1.13 | 0.340 | 3 | 0.003 | 1.13 | 0.290 | 1 | 0.003 | 1.45 | 0.23 | 1 | 0.004 | 0.003 |
| 4th | 0.28 | 0.837 | 3 | <0.001 | 0.94 | 0.333 | 1 | 0.003 | 0.57 | 0.451 | 1 | 0.002 | 0.003 | |
| Treatment | 1st Internode Length cm | 2nd Internode Length cm | 3rd Internode Length cm | 4th Internode Length cm | |
|---|---|---|---|---|---|
| PGR | Control | 2.23 ± 0.75 a | 9.49 ± 1.78 a | 21.5 ± 5.57 a | 34.2 ± 9.50 a |
| CCC | 1.92 ± 0.49 b | 7.58 ± 1.36 b | 19.8 ± 5.49 ab | 31.9 ± 8.92 a | |
| CCC+ET | 2.10 ± 0.72 ab | 7.76 ± 1.55 b | 18.6 ± 5.22 b | 32.9 ± 8.80 a | |
| CCC+TE | 1.87 ± 0.56 b | 7.25 ± 1.73 b | 18.6 ± 4.62 b | 31.7 ± 9.43 a | |
| Cultivar | Nista | 2.21 ± 0.70 a | 8.37 ± 1.57 a | 19.0 ± 4.57 b | 32.2 ± 8.94 a |
| Dorosa | 1.85 ± 0.54 b | 7.67 ± 2.00 b | 20.3 ± 5.96 a | 33.2 ± 9.38 a | |
| N rate | 40 | 2.04 ± 0.70 a | 8.03 ± 1.79 a | 19.8 ± 5.86 a | 32.7 ± 9.64 a |
| kg ha−1 | 70 | 2.02 ± 0.60 a | 8.01 ± 1.87 a | 19.4 ± 4.79 a | 32.6 ± 8.70 a |
| Treatment | 1st Internode Diameter mm | 2nd Internode Diameter mm | 3rd Internode Diameter mm | 4th Internode Diametermm | |
|---|---|---|---|---|---|
| PGR | Control | 0.900 ± 0.109 a | 0.943 ± 0.128 b | 0.851 ± 0.155 a | 0.651 ± 0.172 a |
| CCC | 0.921 ± 0.110 a | 0.977 ± 0.143 ab | 0.876 ± 0.152 a | 0.688 ± 0.146 a | |
| CCC + ET | 0.923 ± 0.129 a | 0.995 ± 0.131 ab | 0.883 ± 0.134 a | 0.695 ± 0.137 a | |
| CCC + TE | 0.915 ± 0.100 a | 1.014 ± 0.132 a | 0.909 ± 0.140 a | 0.677 ± 0.167 a | |
| Cultivar | Nista | 0.930 ± 0.111 a | 0.983 ± 0.139 a | 0.896 ± 0.151 a | 0.695 ± 0.168 a |
| Dorosa | 0.899 ± 0.112 b | 0.982 ± 0.132 a | 0.864 ± 0.140 a | 0.677 ± 0.145 a | |
| N rate | 40 | 0.920 ± 0.105 a | 0.984 ± 0.138 a | 0.861 ± 0.149 b | 0.667 ± 0.158 a |
| kg ha−1 | 70 | 0.910 ± 0.120 a | 0.981 ± 0.133 a | 0.898 ± 0.141 a | 0.704 ± 0.154 a |
| Treatment | 1st Internode Wall Thickness mm | 2nd Internode Wall Thickness mm | 3rd Internode Wall Thickness mm | 4th Internode Wall Thickness mm | |
|---|---|---|---|---|---|
| PGR | Control | 0.213 ± 0.073 a | 0.170 ± 0.058 b | 0.147 ± 0.051 a | 0.123 ± 0.055 a |
| CCC | 0.212 ± 0.072 a | 0.184 ± 0.053 ab | 0.159 ± 0.051 a | 0.129 ± 0.057 a | |
| CCC+ET | 0.229 ± 0.082 a | 0.199 ± 0.063 a | 0.158 ± 0.057 a | 0.128 ± 0.057 a | |
| CCC+TE | 0.223 ± 0.078 a | 0.195 ± 0.062 ab | 0.164 ± 0.052 a | 0.132 ± 0.060 a | |
| Cultivar | Nista | 0.234 ± 0.079 a | 0.191 ± 0.063 a | 0.160±0.056 a | 0.132 ± 0.063 a |
| Dorosa | 0.205 ± 0.071 b | 0.183 ± 0.056 a | 0.153 ± 0.049 a | 0.125 ± 0.050 a | |
| N rate | 40 | 0.214 ± 0.077 a | 0.185 ± 0.058 a | 0.153 ± 0.052 a | 0.125 ± 0.057 a |
| kg ha−1 | 70 | 0.225 ± 0.076 a | 0.189 ± 0.061 a | 0.161 ± 0.053 a | 0.131 ± 0.057 a |
| Internode Length | Internode Diameter | Internode Wall Thickness | |||
|---|---|---|---|---|---|
| Successive Internode | r (X,Y); p | Successive Internode | r (X,Y); p | Successive Internode | r (X,Y); p |
| 1st | 0.2066; p = 0.001 | 1st | −0.2760; p < 0.001 | 1st | 0.1201; p = 0.055 |
| 2nd | −0.1753; p = 0.005 | 2nd | −0.1707; p = 0.006 | 2nd | 0.0024; p = 0.969 |
| 3rd | −0,3679; p < 0.001 | 3rd | 0.0407; p = 0.663 | 3rd | 0.2505; p < 0.001 |
| 4th | −0.5205; p < 0.001 | 4th | 0.1984; p = 0.001 | 4th | 0.3638; p < 0.001 |
| Specification | PGR | Cultivar | N Rate | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | p | df | MS | F | p | df | MS | F | p | df | MS | |
| Seed yield, Mg ha−1 | 5.07 | 0.002 | 3 | 0.221 | 108.49 | <0.001 | 1 | 4.734 | 16.19 | <0.001 | 1 | 0.706 |
| Straw yield, Mg ha−1 | 3.79 | 0.011 | 3 | 11.384 | 16.83 | <0.001 | 1 | 50.508 | 4.93 | 0.027 | 1 | 14.800 |
| Lodging at flowering, % | 12.50 | <0.001 | 3 | 1387.353 | 12.46 | <0.001 | 1 | 1383.163 | 24.77 | <0.001 | 1 | 2749.838 |
| Lodging at ripening, % | 24.73 | <0.001 | 3 | 6041.687 | 10.52 | 0.001 | 1 | 2570.657 | 11.07 | 0.001 | 1 | 2705.206 |
| Generative tillers, no m−2 | 1.39 | 0.248 | 3 | 550,645.83 | 312.96 | <0.001 | 1 | 124,379,742.23 | 3.39 | 0.067 | 1 | 1,346,589.687 |
| Panicle length, cm | 7.85 | <0.001 | 3 | 8.949 | 251.49 | < 0.001 | 1 | 286.539 | 14.41 | <0.001 | 1 | 16.421 |
| Spikelets, no panicle−1 | 4.56 | 0.004 | 3 | 69.551 | 2.95 | 0.087 | 1 | 44.993 | 31.86 | <0.001 | 1 | 486.415 |
| Florets, no spikelet−1 | 1.79 | 0.149 | 3 | 1.061 | 4.59 | 0.033 | 1 | 2.718 | <0.01 | 0.958 | 1 | 0.002 |
| Seeds, no spikelet−1 | 1.22 | 0.302 | 3 | 0.422 | 5.56 | 0.019 | 1 | 1.920 | 0.89 | 0.346 | 1 | 0.307 |
| TSW, g | 24.50 | <0.001 | 3 | 0.372 | 10.66 | 0.001 | 1 | 0.162 | 0.02 | 0.901 | 1 | <0.001 |
| Treatment | Seed Yield Mg ha−1 | Straw Yield Mg ha−1 | Lodging Index at Flowering % | Lodging Index at Ripening % | Generative Tillers No m−2 | |
|---|---|---|---|---|---|---|
| PGR | Control | 0.97 ± 0.22 b | 5.07 ± 2.02 a | 84.8 ± 14.9 b | 61.4 ± 17.7 b | 2325 ± 903 a |
| CCC | 1.09 ± 0.26 a | 4.42 ± 1.81 ab | 93.8 ± 7.8 a | 80.6 ± 14.6 a | 2443 ± 876 a | |
| CCC + ET | 1.10 ± 0.26 a | 4.48 ± 1.72 ab | 94.8 ± 12.2 a | 79.0 ± 16.0 a | 2540 ± 978 a | |
| CCC + TE | 1.06 ± 0.26 ab | 4.05 ± 1.53 b | 93.6 ± 8.6 a | 82.3 ± 15.9 a | 2451 ± 991 a | |
| Cultivar | Nista | 0.92 ± 0.26 b | 4.950 ± 2.08 a | 89.5 ± 12.6 b | 72.7 ± 16.8 b | 1754 ± 808 b |
| Dorosa | 1.19 ± 0.17 a | 4.062 ± 1.35 b | 94.1 ± 10.6 a | 79.0 ± 18.9 a | 3148 ± 357 a | |
| N rate | 40 | 1.01 ± 0.25 b | 4.266 ± 1.67 b | 95.1 ± 9.4 a | 79.1 ± 17.8 a | 2378 ± 924 a |
| kg ha−1 | 70 | 1.11 ± 0.26 a | 4.747 ± 1.90 a | 88.5 ± 13.1 b | 72.6 ± 17.9 b | 2523 ± 946 a |
| Treatment | Panicle Length cm | Spikelets No Panicle−1 | Florets No Spikelet−1 | Seeds No Spikelet−1 | TSW g | |
|---|---|---|---|---|---|---|
| PGR | Control | 8.89 ± 2.56 b | 24.9 ± 3.95 b | 3.53 ± 0.66 a | 2.42 ± 0.60 a | 0.87 ± 0.18 b |
| CCC | 9.64 ± 1.61 a | 26.1 ± 4.04 ab | 3.71 ± 0.79 a | 2.55 ± 0.58 a | 1.00 ± 0.10 a | |
| CCC + ET | 9.72 ± 1.35 a | 26.5 ±3.45 ab | 3.65 ± 0.71 a | 2.57 ± 0.56 a | 1.03 ± 0.10 a | |
| CCC + TE | 9.51 ± 1.57 a | 27.5 ± 5.06 a | 3.84 ± 0.87 a | 2.61 ± 0.58 a | 1.01 ± 0.10 a | |
| Cultivar | Nista | 10.50 ± 1.11 a | 26.7 ± 4.55 a | 3.79 ± 0.67 a | 2.63 ± 0.52 a | 1.00 ± 0.12 a |
| Dorosa | 8.38 ± 1.15 b | 25.8 ± 3.88 a | 3.58 ± 0.84 b | 2.45 ± 0.63 b | 0.95 ± 0.15 b | |
| N rate | 40 | 9.18 ± 1.16 b | 24.9 ± 3.66 b | 3.69 ± 0.74 a | 2.50 ± 0.59 a | 0.98 ± 0.15 a |
| kg ha−1 | 70 | 9.69 ± 1.60 a | 27.6 ± 4.34 a | 3.68 ± 0.80 a | 2.57 ± 0.57 a | 0.98 ± 0.13 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szczepanek, M.; Baczyński, M.; Graczyk, R. Effects of Plant Growth Regulators and N Rate on the Morphological Traits of Generative Tillers and Seed Yields of Chewings and Strong Creeping Red Fescue. Agronomy 2021, 11, 65. https://doi.org/10.3390/agronomy11010065
Szczepanek M, Baczyński M, Graczyk R. Effects of Plant Growth Regulators and N Rate on the Morphological Traits of Generative Tillers and Seed Yields of Chewings and Strong Creeping Red Fescue. Agronomy. 2021; 11(1):65. https://doi.org/10.3390/agronomy11010065
Chicago/Turabian StyleSzczepanek, Małgorzata, Maciej Baczyński, and Radomir Graczyk. 2021. "Effects of Plant Growth Regulators and N Rate on the Morphological Traits of Generative Tillers and Seed Yields of Chewings and Strong Creeping Red Fescue" Agronomy 11, no. 1: 65. https://doi.org/10.3390/agronomy11010065
APA StyleSzczepanek, M., Baczyński, M., & Graczyk, R. (2021). Effects of Plant Growth Regulators and N Rate on the Morphological Traits of Generative Tillers and Seed Yields of Chewings and Strong Creeping Red Fescue. Agronomy, 11(1), 65. https://doi.org/10.3390/agronomy11010065