Characterization of Celiac Disease-Related Epitopes and Gluten Fractions, and Identification of Associated Loci in Durum Wheat

 ,
,  ,
,  ,
,  ,
,  ,
,  , ,
, ,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Field Trials
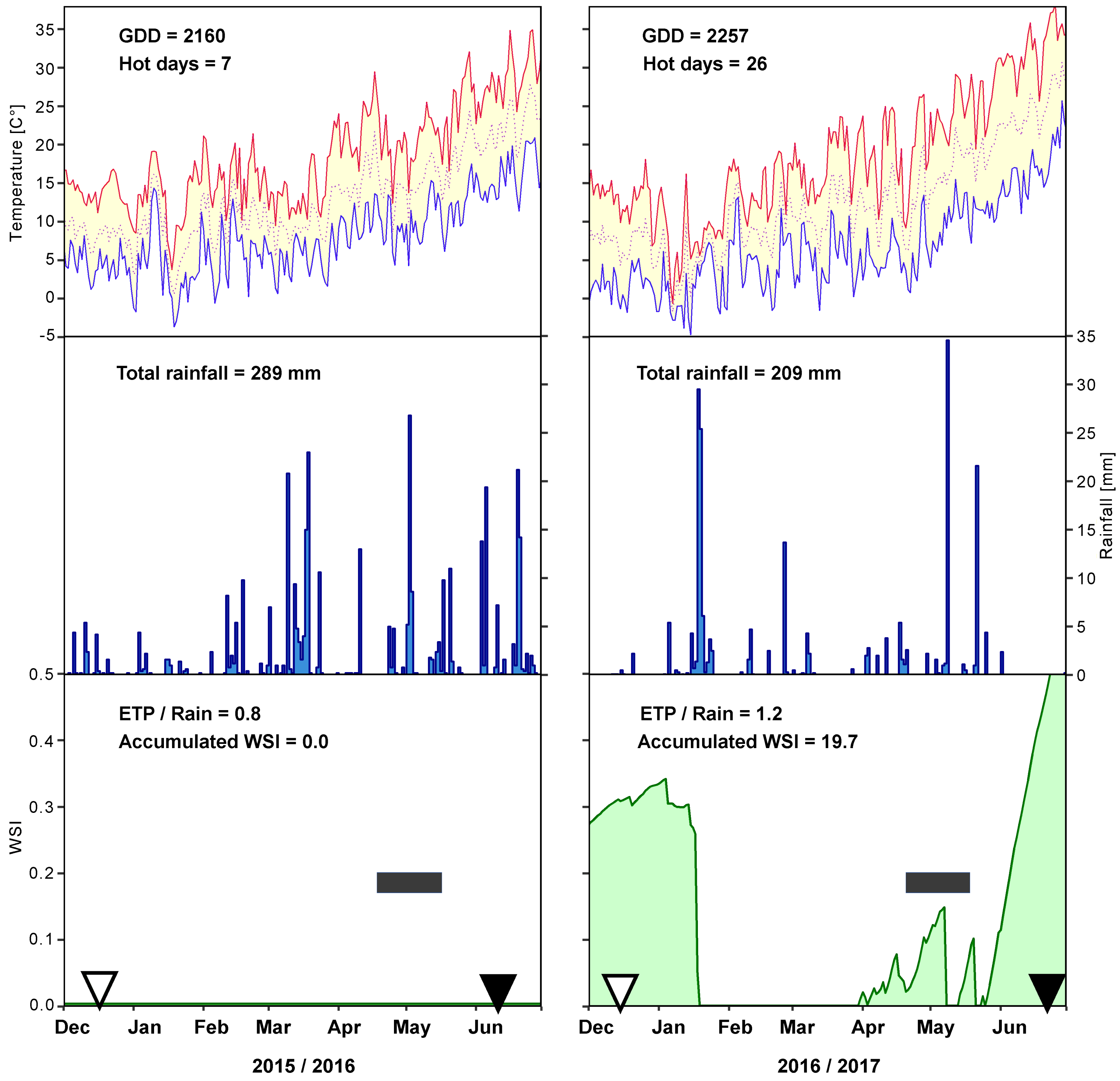
2.2. Weather Conditions
2.3. Grain Quality Analyses
2.4. In Vitro Digestion of Durum Wheat Flours and Quantification of Gluten-Derived Immunogenic/Toxic Peptides
2.5. Genotyping
2.6. GWAS and Identification of Candidate Genes
3. Results
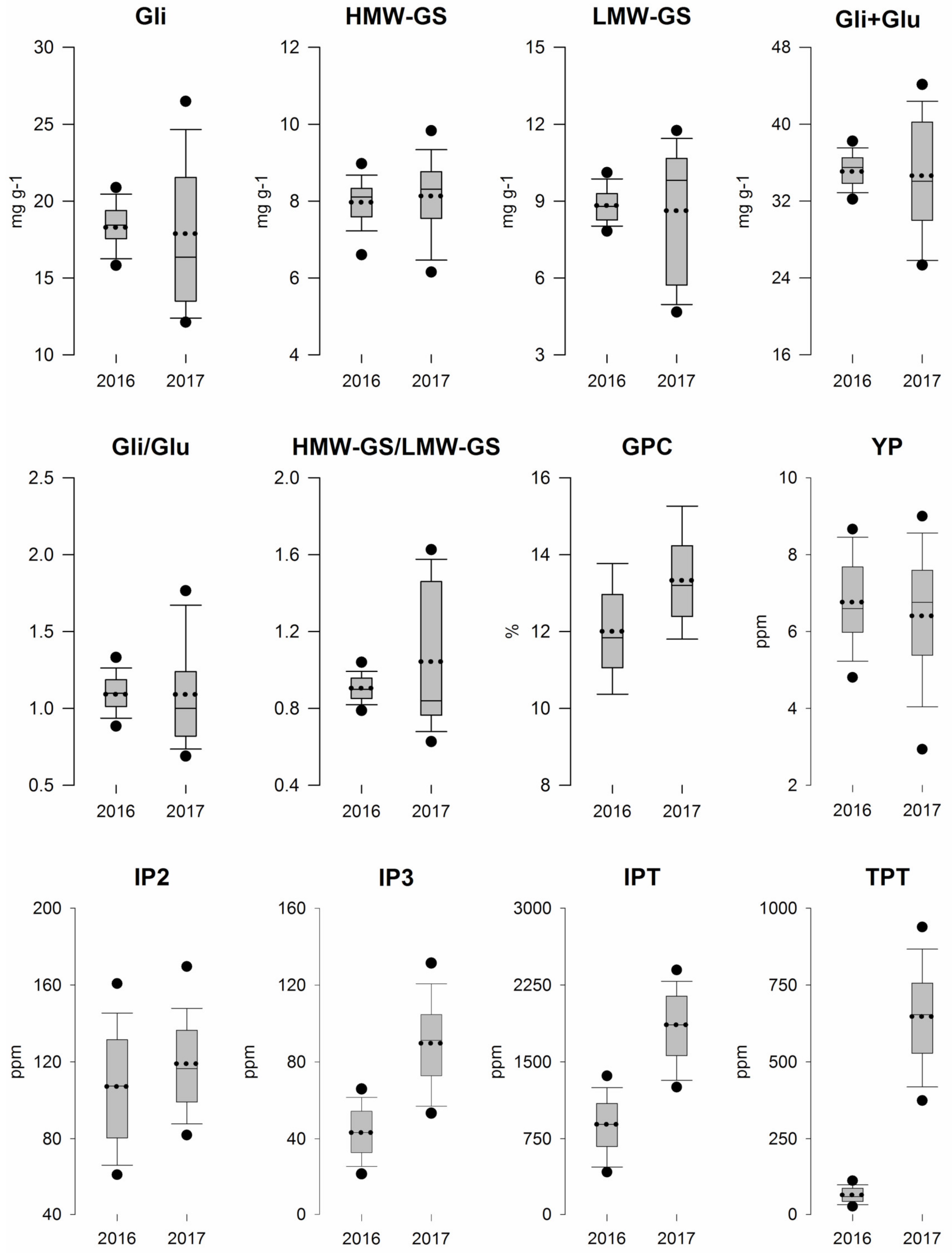
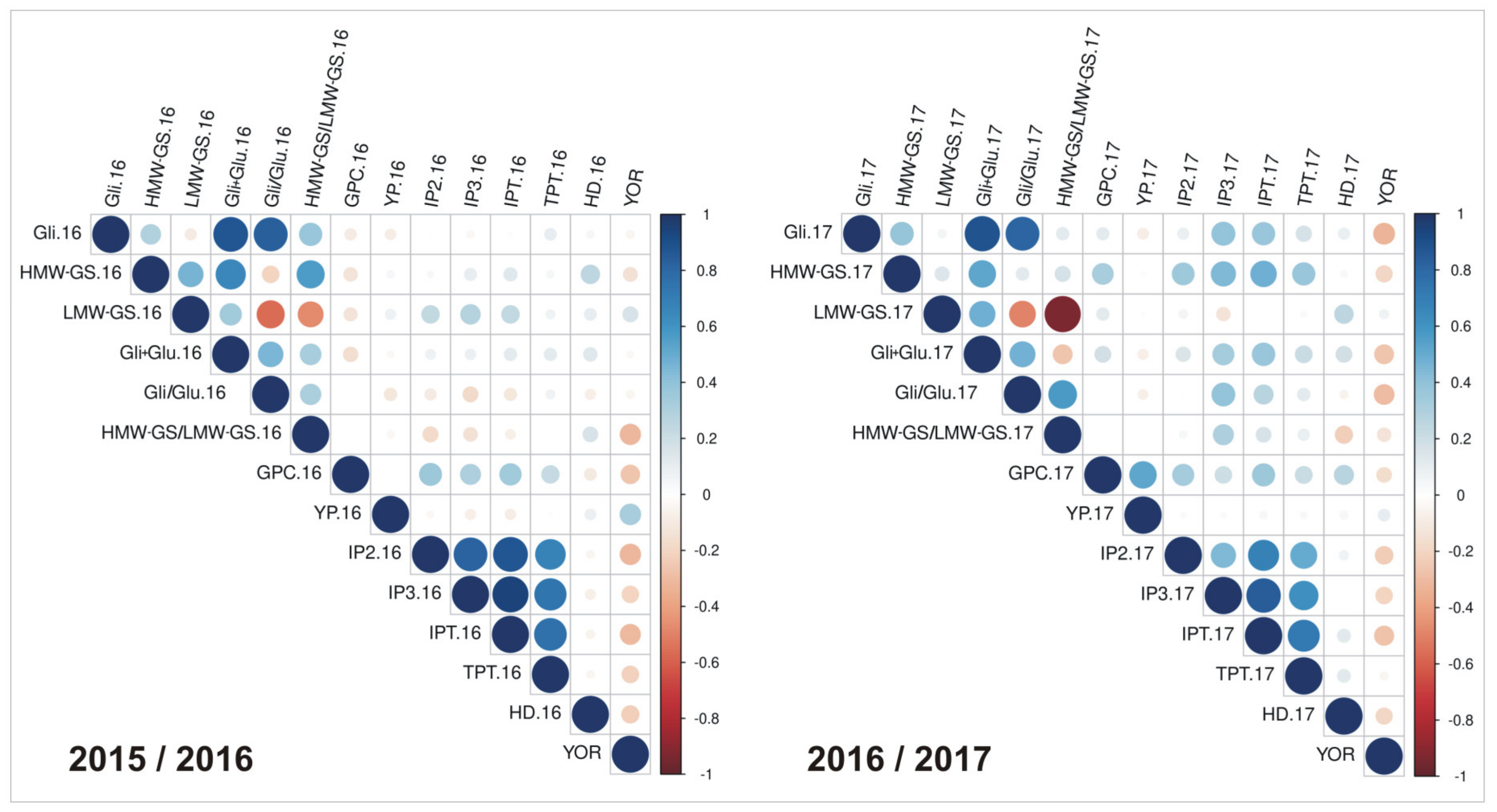
3.1. Field Trials and Grain Quality Analysis
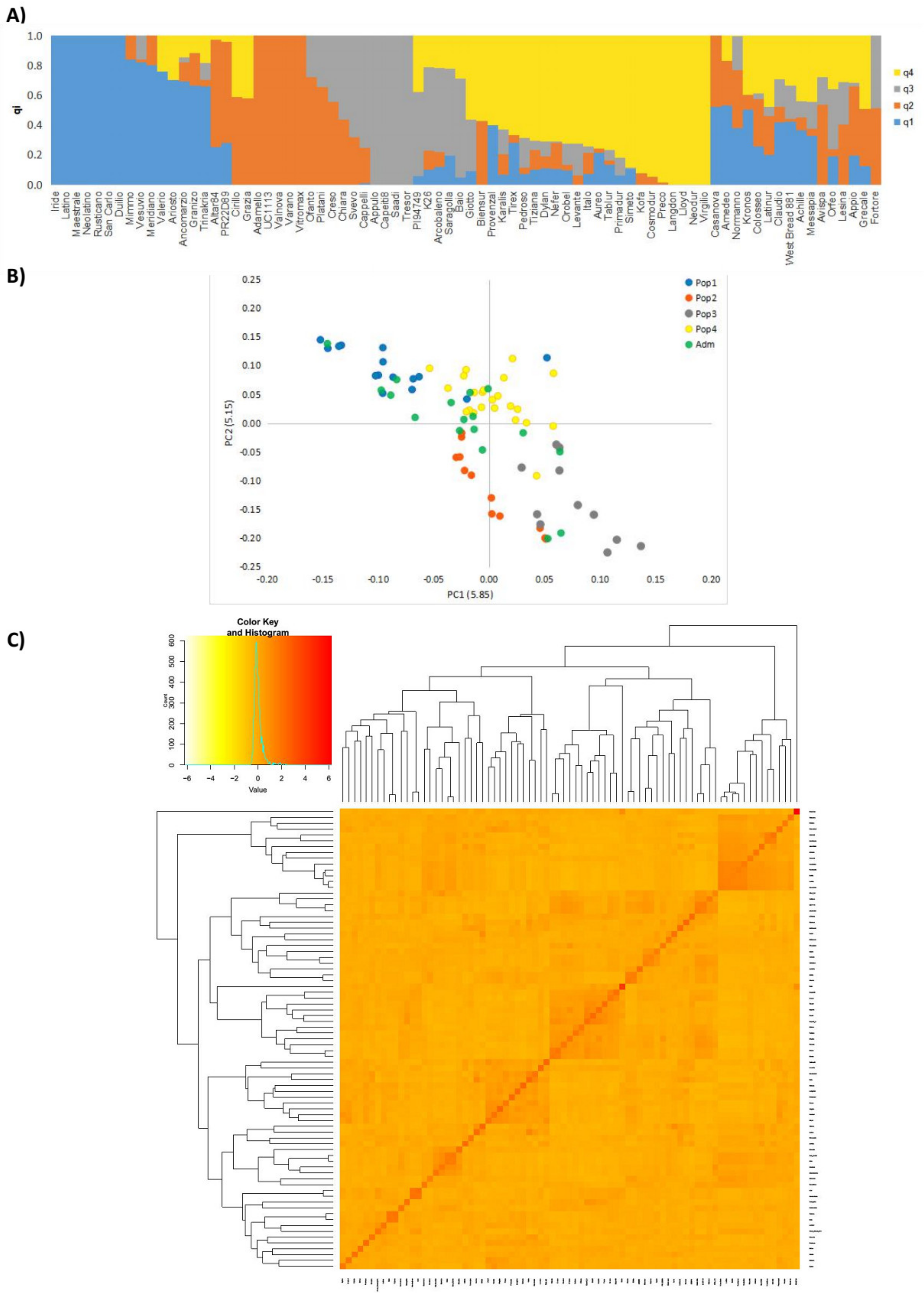
3.2. Population Structure and Linkage Disequilibrium
3.3. Genome-Wide Association Studies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Xynias, I.N.; Mylonas, I.; Korpetis, E.G.; Ninou, E.; Tsaballa, A.; Avdikos, I.D.; Mavromatis, A.G. Durum wheat breeding in the Mediterranean region: Current status and future prospects. Agronomy 2020, 10, 432. [Google Scholar] [CrossRef]
- Lee, S.J.; Rha, M.; Koh, W.; Park, W.; Lee, C.; Kwon, Y.A.; Hwang, J.-K. Measurement of Cooked Noodle Stickiness Using a Modified Instrumental Method. Cereal Chem. J. 2002, 79, 838–842. [Google Scholar] [CrossRef]
- Sissons, M.; Sestili, F.; Botticella, E.; Masci, S.; Lafiandra, D. Can Manipulation of Durum Wheat Amylose Content Reduce the Glycaemic Index of Spaghetti? Foods 2020, 9, 693. [Google Scholar] [CrossRef] [PubMed]
- Sissons, M.J.; Egan, N.E.; Gianibelli, M.C. New Insights Into the Role of Gluten on Durum Pasta Quality Using Reconstitution Method. Cereal Chem. J. 2005, 82, 601–608. [Google Scholar] [CrossRef]
- Cubadda, R.E.; Carcea, M.; Marconi, E.; Trivisonno, M.C. Influence of Gluten Proteins and Drying Temperature on the Cooking Quality of Durum Wheat Pasta. Cereal Chem. J. 2007, 84, 48–55. [Google Scholar] [CrossRef]
- Sissons, M. Role of durum wheat composition on the quality of pasta and bread. Food 2008, 2, 75–90. [Google Scholar]
- Varzakas, T.; Kozub, N.; Xynias, I.N. Quality determination of wheat: Genetic determination, biochemical markers, seed storage proteins—Bread and durum wheat germplasm. J. Sci. Food Agric. 2014, 94, 2819–2829. [Google Scholar] [CrossRef]
- De Vita, P.; Nicosia, O.L.D.; Nigro, F.; Platani, C.; Riefolo, C.; Di Fonzo, N.; Cattivelli, L. Breeding progress in morpho-physiological, agronomical and qualitative traits of durum wheat cultivars released in Italy during the 20th century. Eur. J. Agron. 2007, 26, 39–53. [Google Scholar] [CrossRef]
- Sapone, A.; Bai, J.C.; Ciacci, C.; Dolinsek, J.; Green, P.H.R.; Hadjivassiliou, M.; Kaukinen, K.; Rostami, K.; Sanders, D.S.; Schumann, M.; et al. Spectrum of gluten-related disorders: Consensus on new nomenclature and classification. BMC Med. 2012. [Google Scholar] [CrossRef]
- Ficco, D.B.M.; Prandi, B.; Amaretti, A.; Anfelli, I.; Leonardi, A.; Raimondi, S.; Pecchioni, N.; De Vita, P.; Faccini, A.; Sforza, S.; et al. Comparison of gluten peptides and potential prebiotic carbohydrates in old and modern Triticum turgidum ssp. genotypes. Food Res. Int. 2019. [Google Scholar] [CrossRef]
- Catassi, C.; Fasano, A. Celiac disease. Curr. Opin. Gastroenterol. 2008, 24, 687–691. [Google Scholar] [CrossRef] [PubMed]
- Ciccocioppo, R.; Di Sabatino, A.; Corazza, G.R. The immune recognition of gluten in coeliac disease. Clin. Exp. Immunol. 2005, 140, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Tye-Din, J.A.; Galipeau, H.J.; Agardh, D. Celiac disease: A review of current concepts in pathogenesis, prevention, and novel therapies. Front. Pediatrics 2018, 6, 350. [Google Scholar] [CrossRef] [PubMed]
- Van Herpen, T.W.J.M.; Goryunova, S.V.; van der Schoot, J.; Mitreva, M.; Salentijn, E.; Vorst, O.; Schenk, M.F.; van Veelen, P.A.; Koning, F.; van Soest, L.J.M.; et al. Alpha-gliadin genes from the A, B, and D genomes of wheat contain different sets of celiac disease epitopes. BMC Genom. 2006. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.G.; Liang, H.H.; Bai, S.L.; Zhou, Y.; Sun, G.; Su, Y.R.; Gao, A.L.; Zhang, D.L.; Li, S.P. Molecular Characterization and Variation of the Celiac Disease Epitope Domains among α-Gliadin Genes in Aegilops tauschii. J. Agric. Food Chem. 2017. [Google Scholar] [CrossRef] [PubMed]
- Ozuna, C.V.; Barro, F. Characterization of gluten proteins and celiac disease-related immunogenic epitopes in the Triticeae: Cereal domestication and breeding contributed to decrease the content of gliadins and gluten. Mol. Breed. 2018. [Google Scholar] [CrossRef]
- Camerlengo, F.; Sestili, F.; Silvestri, M.; Colaprico, G.; Margiotta, B.; Ruggeri, R.; Lupi, R.; Masci, S.; Lafiandra, D. Production and molecular characterization of bread wheat lines with reduced amount of $α$-type gliadins. BMC Plant Biol. 2017, 17, 248. [Google Scholar] [CrossRef]
- Ronga, D.; Laviano, L.; Catellani, M.; Milc, J.; Prandi, B.; Boukid, F.; Sforza, S.; Dossena, A.; Graziano, S.; Gullì, M.; et al. Influence of environmental and genetic factors on content of toxic and immunogenic wheat gluten peptides. Eur. J. Agron. 2020. [Google Scholar] [CrossRef]
- IWGSC International Wheat Genome Sequencing Consortium(IWGSC). Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018. [Google Scholar] [CrossRef]
- Avni, R.; Nave, M.; Barad, O.; Baruch, K.; Twardziok, S.O.; Gundlach, H.; Hale, I.; Mascher, M.; Spannagl, M.; Wiebe, K.; et al. Wild emmer genome architecture and diversity elucidate wheat evolution and domestication. Science 2017. [Google Scholar] [CrossRef]
- Maccaferri, M.; Harris, N.S.; Twardziok, S.O.; Pasam, R.K.; Gundlach, H.; Spannagl, M.; Ormanbekova, D.; Lux, T.; Prade, V.M.; Milner, S.G.; et al. Durum wheat genome highlights past domestication signatures and future improvement targets. Nat. Genet. 2019. [Google Scholar] [CrossRef] [PubMed]
- Plessis, A.; Ravel, C.; Bordes, J.; Balfourier, F.; Martre, P. Association study of wheat grain protein composition reveals that gliadin and glutenin composition are trans-regulated by different chromosome regions. J. Exp. Bot. 2013. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Li, F.; Cao, S.; Zhang, K. Genomic and functional genomics analyses of gluten proteins and prospect for simultaneous improvement of end-use and health-related traits in wheat. Theor. Appl. Genet. 2020, 133, 1521–1539. [Google Scholar] [CrossRef] [PubMed]
- Pavan, S.; Delvento, C.; Ricciardi, L.; Lotti, C.; Ciani, E.; D’Agostino, N. Recommendations for Choosing the Genotyping Method and Best Practices for Quality Control in Crop Genome—Wide Association Studies. Front. Genet. 2020. [Google Scholar] [CrossRef] [PubMed]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974. [Google Scholar] [CrossRef]
- Clarke, F.R.; Baker, R.J.; DePauw, R.M. Interplot Interference Disorts Yield Estimates in Spring Wheat. Crop Sci. 1998, 38, 62–66. [Google Scholar] [CrossRef]
- Rizza, F.; Badeck, F.W.; Cattivelli, L.; Lidestri, O.; Di Fonzo, N.; Stanca, A.M. Use of a Water Stress Index to Identify Barley Genotypes Adapted to Rainfed and Irrigated Conditions. Crop Sci. 2004, 44, 2127–2137. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2 Elegant Graphics for Data Analysis (Use R!); Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Visioli, G.; Galieni, A.; Stagnari, F.; Bonas, U.; Speca, S.; Faccini, A.; Pisante, M.; Marmiroli, N. Proteomics of durum wheat grain during transition to conservation agriculture. PLoS ONE 2016, 11, e0156007. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food-an international consensus. Food Funct. 2014. [Google Scholar] [CrossRef]
- Boukid, F.; Prandi, B.; Sforza, S.; Sayar, R.; Seo, Y.W.; Mejri, M.; Yacoubi, I. Understanding the Effects of Genotype, Growing Year, and Breeding on Tunisian Durum Wheat Allergenicity. 2. The Celiac Disease Case. J. Agric. Food Chem. 2017. [Google Scholar] [CrossRef]
- Prandi, B.; Tedeschi, T.; Folloni, S.; Galaverna, G.; Sforza, S. Peptides from gluten digestion: A comparison between old and modern wheat varieties. Food Res. Int. 2017. [Google Scholar] [CrossRef] [PubMed]
- Muqaddasi, Q.H.; Reif, J.C.; Li, Z.; Basnet, B.R.; Dreisigacker, S.; Röder, M.S. Genome-wide association mapping and genome-wide prediction of anther extrusion in CIMMYT spring wheat. Euphytica 2017, 213, 1–7. [Google Scholar] [CrossRef]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.A.M.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. Gigascience 2015. [Google Scholar] [CrossRef] [PubMed]
- Alexander, D.H.; Novembre, J.; Lange, K. Fast model-based estimation of ancestry in unrelated individuals. Genome Res. 2009. [Google Scholar] [CrossRef] [PubMed]
- Glaubitz, J.C.; Casstevens, T.M.; Lu, F.; Harriman, J.; Elshire, R.J.; Sun, Q.; Buckler, E.S. TASSEL-GBS: A high capacity genotyping by sequencing analysis pipeline. PLoS ONE 2014, 9, e90346. [Google Scholar] [CrossRef] [PubMed]
- Kidane, Y.G.; Mancini, C.; Mengistu, D.K.; Frascaroli, E.; Fadda, C.; Pè, M.E.; Dell’Acqua, M. Genome wide association study to identify the genetic base of smallholder farmer preferences of durum wheat traits. Front. Plant Sci. 2017. [Google Scholar] [CrossRef] [PubMed]
- Taranto, F.; D’Agostino, N.; Rodriguez, M.; Pavan, S.; Minervini, A.P.; Pecchioni, N.; Papa, R.; De Vita, P. Whole Genome Scan Reveals Molecular Signatures of Divergence and Selection Related to Important Traits in Durum Wheat Germplasm. Front. Genet. 2020. [Google Scholar] [CrossRef]
- Lipka, A.E.; Tian, F.; Wang, Q.; Peiffer, J.; Li, M.; Bradbury, P.J.; Gore, M.A.; Buckler, E.S.; Zhang, Z. GAPIT: Genome association and prediction integrated tool. Bioinformatics 2012. [Google Scholar] [CrossRef]
- Liu, X.; Huang, M.; Fan, B.; Buckler, E.S.; Zhang, Z. Iterative Usage of Fixed and Random Effect Models for Powerful and Efficient Genome-Wide Association Studies. PLoS Genet. 2016. [Google Scholar] [CrossRef]
- Maccaferri, M.; Ricci, A.; Salvi, S.; Milner, S.G.; Noli, E.; Martelli, P.L.; Casadio, R.; Akhunov, E.; Scalabrin, S.; Vendramin, V.; et al. A high-density, SNP-based consensus map of tetraploid wheat as a bridge to integrate durum and bread wheat genomics and breeding. Plant Biotechnol. J. 2015. [Google Scholar] [CrossRef]
- Blanco, A.; Colasuonno, P.; Gadaleta, A.; Mangini, G.; Schiavulli, A.; Simeone, R.; Digesù, A.M.; De Vita, P.; Mastrangelo, A.M.; Cattivelli, L. Quantitative trait loci for yellow pigment concentration and individual carotenoid compounds in durum wheat. J. Cereal Sci. 2011. [Google Scholar] [CrossRef]
- Colasuonno, P.; Lozito, M.L.; Marcotuli, I.; Nigro, D.; Giancaspro, A.; Mangini, G.; De Vita, P.; Mastrangelo, A.M.; Pecchioni, N.; Houston, K.; et al. The carotenoid biosynthetic and catabolic genes in wheat and their association with yellow pigments. BMC Genom. 2017. [Google Scholar] [CrossRef] [PubMed]
- Zhai, S.; Liu, J.; Xu, D.; Wen, W.; Yan, J.; Zhang, P.; Wan, Y.; Cao, S.; Hao, Y.; Xia, X.; et al. A genome-wide association study reveals a rich genetic architecture of flour color-related traits in bread wheat. Front. Plant Sci. 2018. [Google Scholar] [CrossRef] [PubMed]
- Pozniak, C.J.; Knox, R.E.; Clarke, F.R.; Clarke, J.M. Identification of QTL and association of a phytoene synthase gene with endosperm colour in durum wheat. Theor. Appl. Genet. 2007, 114, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Colasuonno, P.; Marcotuli, I.; Blanco, A.; Maccaferri, M.; Condorelli, G.E.; Tuberosa, R.; Parada, R.; de Camargo, A.C.; Schwember, A.R.; Gadaleta, A. Carotenoid Pigment Content in Durum Wheat (Triticum turgidum L. var durum): An Overview of Quantitative Trait Loci and Candidate Genes. Front. Plant Sci. 2019, 10, 1347. [Google Scholar] [CrossRef]
- Dhar, M.K.; Koul, A.; Kaul, S. Farnesyl pyrophosphate synthase: A key enzyme in isoprenoid biosynthetic pathway and potential molecular target for drug development. New Biotechnol. 2013, 30, 114–123. [Google Scholar] [CrossRef]
- Patil, R.M.; Oak, M.D.; Tamhankar, S.A.; Sourdille, P.; Rao, V.S. Mapping and validation of a major QTL for yellow pigment content on 7AL in durum wheat (Triticum turgidum L. ssp. durum). Mol. Breed. 2008. [Google Scholar] [CrossRef]
- Roncallo, P.F.; Cervigni, G.L.; Jensen, C.; Miranda, R.; Carrera, A.D.; Helguera, M.; Echenique, V. QTL analysis of main and epistatic effects for flour color traits in durum wheat. Euphytica 2012. [Google Scholar] [CrossRef]
- Fiedler, J.D.; Salsman, E.; Liu, Y.; Michalak de Jiménez, M.; Hegstad, J.B.; Chen, B.; Manthey, F.A.; Chao, S.; Xu, S.; Elias, E.M.; et al. Genome-Wide Association and Prediction of Grain and Semolina Quality Traits in Durum Wheat Breeding Populations. Plant Genome 2017. [Google Scholar] [CrossRef]
- Simmonds, N.W. The relation between yield and protein in cereal grain. J. Sci. Food Agric. 1995. [Google Scholar] [CrossRef]
- Fufa, H.; Baenziger, P.S.; Beecher, B.S.; Graybosch, R.A.; Eskridge, K.M.; Nelson, L.A. Genetic improvement trends in agronomic performances and end-use quality characteristics among hard red winter wheat cultivars in Nebraska. Euphytica 2005, 144, 187–198. [Google Scholar] [CrossRef]
- Brevis, J.C.; Dubcovsky, J. Effects of the chromosome region including the Gpc-B1 locus on wheat grain and protein yield. Crop Sci. 2010. [Google Scholar] [CrossRef]
- Shewry, P.R. Improving the protein content and composition of cereal grain. J. Cereal Sci. 2007. [Google Scholar] [CrossRef]
- Aguirrezábal, L.; Martre, P.; Pereyra-Irujo, G.; Izquierdo, N.; Allard, V. Management and Breeding Strategies for the Improvement of Grain and Oil Quality. In Crop Physiology; Elsevier: Amsterdam, The Netherlands, 2009. [Google Scholar]
- Branlard, G.; Dardevet, M.; Saccomano, R.; Lagoutte, F.; Gourdon, J. Genetic diversity of wheat storage proteins and bread wheat quality. Euphytica 2001. [Google Scholar] [CrossRef]
- Sharma, N.; Bhatia, S.; Chunduri, V.; Kaur, S.; Sharma, S.; Kapoor, P.; Kumari, A.; Garg, M. Pathogenesis of Celiac Disease and Other Gluten Related Disorders in Wheat and Strategies for Mitigating Them. Front. Nutr. 2020. [Google Scholar] [CrossRef]
- Slafer, G.A.; Araus, J.L.; Royo, C.; García Del Moral, L.F. Promising eco-physiological traits for genetic improvement of cereal yields in Mediterranean environments. Ann. Appl. Biol. 2005, 146, 61–70. [Google Scholar] [CrossRef]
- Francia, E.; Tondelli, A.; Rizza, F.; Badeck, F.W.; Li Destri Nicosia, O.; Akar, T.; Grando, S.; Al-Yassin, A.; Benbelkacem, A.; Thomas, W.T.B.; et al. Determinants of barley grain yield in a wide range of Mediterranean environments. Field Crop Res. 2011, 120, 169–178. [Google Scholar] [CrossRef]
- Isidro, J.; Álvaro, F.; Royo, C.; Villegas, D.; Miralles, D.J.; García Del Moral, L.F. Changes in duration of developmental phases of durum wheat caused by breeding in Spain and Italy during the 20th century and its impact on yield. Ann. Bot. 2011. [Google Scholar] [CrossRef]
- Giunta, F.; Motzo, R.; Pruneddu, G. Has long-term selection for yield in durum wheat also induced changes in leaf and canopy traits? Field Crop Res. 2008. [Google Scholar] [CrossRef]
- Giunta, F.; Motzo, R.; Virdis, A. Development of durum wheat and triticale cultivars as affected by thermo-photoperiodic conditions. Aust. J. Agric. Res. 2001. [Google Scholar] [CrossRef]
- Francia, E.; Tondelli, A.; Rizza, F.; Badeck, F.W.; Thomas, W.T.B.; van Eeuwijk, F.; Romagosa, I.; Michele Stanca, A.; Pecchioni, N. Determinants of barley grain yield in drought-prone Mediterranean environments. Ital. J. Agron. 2013, 8, 1–8. [Google Scholar] [CrossRef]
- Triboi, E.; Martre, P.; Girousse, C.; Ravel, C.; Triboi-Blondel, A.M. Unravelling environmental and genetic relationships between grain yield and nitrogen concentration for wheat. Eur. J. Agron. 2006. [Google Scholar] [CrossRef]
- Dupont, F.M.; Hurkman, W.J.; Vensel, W.H.; Tanaka, C.; Kothari, K.M.; Chung, O.K.; Altenbach, S.B. Protein accumulation and composition in wheat grains: Effects of mineral nutrients and high temperature. Eur. J. Agron. 2006. [Google Scholar] [CrossRef]
- Shewry, P.R.; Tatham, A.S. Improving wheat to remove coeliac epitopes but retain functionality. J. Cereal Sci. 2016, 67, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Taranto, F.; Nicolia, A.; Pavan, S.; De Vita, P.; D’Agostino, N. Biotechnological and Digital Revolution for Climate-Smart Plant Breeding. Agronomy 2018, 8, 277. [Google Scholar] [CrossRef]
- Merlino, M.; Leroy, P.; Chambon, C.; Branlard, G. Mapping and proteomic analysis of albumin and globulin proteins in hexaploid wheat kernels (Triticum aestivum L.). Theor. Appl. Genet. 2009, 118, 1321–1337. [Google Scholar] [CrossRef] [PubMed]
- She, M.; Ye, X.; Yan, Y.; Howit, C.; Belgard, M.; Ma, W. Gene networks in the synthesis and deposition of protein polymers during grain development of wheat. Funct. Integr. Genom. 2011, 11, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Noma, S.; Kawaura, K.; Hayakawa, K.; Abe, C.; Tsuge, N.; Ogihara, Y. Comprehensive molecular characterization of the α/β-gliadin multigene family in hexaploid wheat. Mol. Genet. Genom. 2016. [Google Scholar] [CrossRef]
- Ravel, C.; Fiquet, S.; Boudet, J.; Dardevet, M.; Vincent, J.; Merlino, M.; Michard, R.; Martre, P. Conserved cis-regulatory modules in promoters of genes encoding wheat high-molecular-weight glutenin subunits. Front. Plant Sci. 2014. [Google Scholar] [CrossRef]
- Duan, L.; Han, S.; Wang, K.; Jiang, P.; Gu, Y.; Chen, L.; Mu, J.; Ye, X.; Li, Y.; Yan, Y.; et al. Analyzing the action of evolutionarily conserved modules on HMW-GS 1Ax1 promoter activity. Plant Mol. Biol. 2020. [Google Scholar] [CrossRef]
- Van Herpen, T.W.J.M.; Riley, M.; Sparks, C.; Jones, H.D.; Gritsch, C.; Dekking, E.H.; Hamer, R.J.; Bosch, D.; Salentijn, E.M.J.; Smulders, M.J.M.; et al. Detailed analysis of the expression of an alpha-gliadin promoter and the deposition of alpha-gliadin protein during wheat grain development. Ann. Bot. 2008. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, K.; Li, G.; Li, Y.; Zhang, Y.; Liu, Z.; Ye, X.; Xia, X.; He, Z.; Cao, S. Dissecting conserved cis-regulatory modules of Glu-1 promoters which confer the highly active endosperm-specific expression via stable wheat transformation. Crop J. 2019. [Google Scholar] [CrossRef]
- Boudet, J.; Merlino, M.; Plessis, A.; Gaudin, J.C.; Dardevet, M.; Perrochon, S.; Alvarez, D.; Risacher, T.; Martre, P.; Ravel, C. The bZIP transcription factor SPA Heterodimerizing Protein represses glutenin synthesis in Triticum aestivum. Plant J. 2019. [Google Scholar] [CrossRef] [PubMed]
- Ravel, C.; Martre, P.; Romeuf, I.; Dardevet, M.; El-Malki, R.; Bordes, J.; Duchateau, N.; Brunel, D.; Balfourier, F.; Charmet, G. Nucleotide polymorphism in the wheat transcriptional activator spa influences its pattern of expression and has pleiotropic effects on grain protein composition, dough viscoelasticity, and grain hardness. Plant Physiol. 2009, 151, 2133–2144. [Google Scholar] [CrossRef]
- Dong, G.; Ni, Z.; Yao, Y.; Nie, X.; Sun, Q. Wheat Dof transcription factor WPBF interacts with TaQM and activates transcription of an alpha-gliadin gene during wheat seed development. Plant Mol. Biol. 2007, 63, 73–84. [Google Scholar] [CrossRef]
- Moehs, C.P.; Austill, W.J.; Holm, A.; Large, T.A.G.; Loeffler, D.; Mullenberg, J.; Schnable, P.S.; Skinner, W.; Van Boxtel, J.; Wu, L.; et al. Development of decreased-gluten wheat enabled by determination of the genetic basis of lys3a barley. Plant Physiol. 2019. [Google Scholar] [CrossRef]
- Ravel, C.; Nagy, I.J.; Martre, P.; Sourdille, P.; Dardevet, M.; Balfourier, F.; Pont, C.; Giancola, S.; Praud, S.; Charmet, G. Single nucleotide polymorphism, genetic mapping, and expression of genes coding for the DOF wheat prolamin-box binding factor. Funct. Integr. Genom. 2006, 6, 310–321. [Google Scholar] [CrossRef]
- Branlard, G.; Lesage, V.S.; Bancel, E.; Martre, P.; Méleard, B.; Rhazi, L. Coping with Wheat Quality in a Changing Environment: Proteomics Evidence for Stress Caused by Environmental Changes. In Advances in Wheat Genetics: From Genome to Field; Springer: Tokyo, Japan, 2015; pp. 255–264. [Google Scholar]
- Salentijn, E.M.J.; Esselink, D.G.; Goryunova, S.V.; van der Meer, I.M.; Gilissen, L.J.W.J.; Smulders, M.J.M. Quantitative and qualitative differences in celiac disease epitopes among durum wheat varieties identified through deep RNA-amplicon sequencing. BMC Genom. 2013, 14, 1–16. [Google Scholar] [CrossRef]
- Spaenij-Dekking, L.; Kooy-Winkelaar, Y.; Van Veelen, P.; Drijfhout, J.W.; Jonker, H.; Van Soest, L.; Smulders, M.J.M.; Bosch, D.; Gilissen, L.J.W.J.; Koning, F. Natural variation in toxicity of wheat: Potential for selection of nontoxic varieties for celiac disease patients. Gastroenterology 2005, 129, 797–806. [Google Scholar] [CrossRef]
- Zhang, H.; Zhu, H.; Pan, Y.; Yu, Y.; Luan, S.; Li, L. A DTX/MATE-type transporter facilitates abscisic acid efflux and modulates ABA sensitivity and drought tolerance in Arabidopsis. Mol. Plant 2014. [Google Scholar] [CrossRef]
- Takamiya, K.I.; Tsuchiya, T.; Ohta, H. Degradation pathway(s) of chlorophyll: What has gene cloning revealed? Trends Plant Sci. 2000, 5, 426–431. [Google Scholar] [CrossRef]
- Schreiber, A.W.; Sutton, T.; Caldo, R.A.; Kalashyan, E.; Lovell, B.; Mayo, G.; Muehlbauer, G.J.; Druka, A.; Waugh, R.; Wise, R.P.; et al. Comparative transcriptomics in the Triticeae. BMC Genom. 2009. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wei, K. Comparative functional genomics analysis of cytochrome P450 gene superfamily in wheat and maize. BMC Plant Biol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Morita, R.; Sato, Y.; Masuda, Y.; Nishimura, M.; Kusaba, M. Defect in non-yellow coloring 3, an α/β hydrolase-fold family protein, causes a stay-green phenotype during leaf senescence in rice. Plant J. 2009. [Google Scholar] [CrossRef] [PubMed]
- Distelfeld, A.; Avni, R.; Fischer, A.M. Senescence, nutrient remobilization, and yield in wheat and barley. J. Exp. Bot. 2014, 65, 3783–3798. [Google Scholar] [CrossRef]
- Shi, S.; Azam, F.I.; Li, H.; Chang, X.; Li, B.; Jing, R. Mapping QTL for stay-green and agronomic traits in wheat under diverse water regimes. Euphytica 2017. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
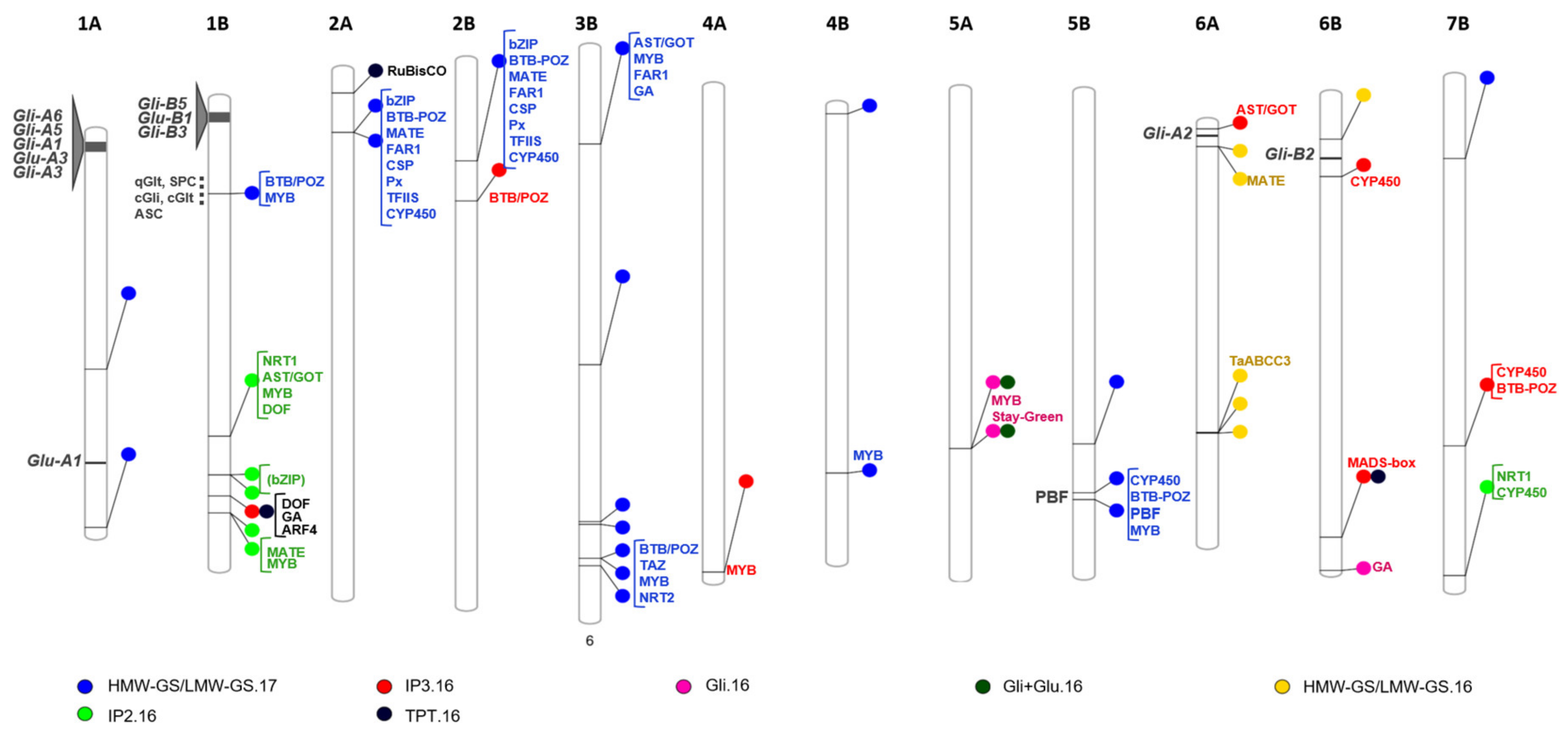
| Trait | SNP | Chr. | Position (bp) | cM | p-Value | R2 § | Allelic Effect | Transcript ID | Annotation |
|---|---|---|---|---|---|---|---|---|---|
| Protein Component | |||||||||
| Gli.16 | BobWhite_c19155_246 | 5A | 528,942,215 | 111.5 | 2.58 × 10−7 | 0.269 | 2.478 | TRIDC5AG047350 | Protein STAY-GREEN, chloroplastic |
| wsnp_Ex_c4026_7281501 | 5A | 529,160,493 | 111.5 | 2.58 × 10−7 | 0.269 | −3.61 | TRIDC5AG047410 | Two-component response regulator-like PRR95 | |
| BS00003958_51 | 6B | 701,071,618 | n.a. | 1.69 × 10−5 | 0.189 | 1.802 | TRIDC6BG073020 | Unknown function | |
| HMW-GS/LMW-GS.16 a | Tdurum_contig42729_433 | 6A | 34,690,496 | 29.1 | 3.37 × 10−5 | n.a. | 0.037 | TRIDC6AG008160 | MATE efflux family protein |
| Tdurum_contig42729_380 | 6A | 34,690,549 | 29.1 | 3.37 × 10−5 | n.a. | −0.037 | TRIDC6AG008160 | MATE efflux family protein | |
| Tdurum_contig78006_158 | 6A | 456,240,525 | 56.8 | 2.59 × 10−5 | n.a. | 0.035 | TRIDC6AG037510 | Fatty acid oxidation complex subunit α | |
| Tdurum_contig76709_195 | 6A | 456,677,777 | 58.4 | 7.13 × 10−5 | n.a. | 0.032 | TRIDC6AG037650 | α/β-hydrolases superfamily protein | |
| BS00036878_51 | 6A | 457,248,758 | 58.4 | 4.32 × 10−5 | n.a. | 0.037 | TRIDC6AG037700 | Receptor-like kinase 1 | |
| IAAV7349 | 6B | 64,846,436 | n.a. | 3.37 × 10−5 | n.a. | 0.037 | TRIDC6BG011970 | ARC6 | |
| HMW-GS/LMW-GS.17 a | GENE-1214_288 | 1A | 349,690,319 | n.a. | 0.0000079 | n.a. | −3.308 | TRIDC1AG029170 | Armadillo repeat-containing protein 7 |
| BS00065170_51 | 1A | 582,636,662 | 156 | 0.0000079 | n.a. | −3.944 | TRIDC1AG063120 | Plant protein of unknown function (DUF247) | |
| BS00035690_51 | 1B | 138,929,471 | 34.6 | 0.0000079 | n.a. | −3.083 | TRIDC1BG017610 | Glucan 1,3-β-glucosidase A | |
| Kukri_c40953_658 | 2A | 91,415,342 | n.a. | 0.0000079 | n.a. | −3.308 | TRIDC2AG017740 | Cytochrome P450 superfamily protein | |
| Kukri_c67546_279 | 2A | 91,421,659 | n.a. | 0.0000079 | n.a. | −3.308 | TRIDC2AG017760 | Sulfotransferase | |
| RFL_Contig1385_326 | 2B | 147,198,649 | n.a. | 0.0000079 | n.a. | −1.444 | TRIDC2BG021670 | RNA-binding protein 39 | |
| RAC875_c25375_236 | 3B | 140,912,116 | n.a. | 0.0000079 | n.a. | −2.281 | TRIDC3BG020330 | UDP-Glycosyltransferase superfamily protein | |
| Kukri_c7804_2504 | 3B | 467,024,111 | n.a. | 0.0000079 | n.a. | −2.281 | TRIDC3BG042750 | Chloride channel E | |
| IACX3426 | 3B | 698,180,277 | n.a. | 0.0000079 | n.a. | −2.281 | TRIDC3BG065770 | Undescribed protein | |
| Ku_c46571_2583 | 3B | 701,422,737 | 130.1 | 0.0000079 | n.a. | −2.281 | TRIDC3BG066320 | Dicer-like 1 | |
| Kukri_c13345_481 | 3B | 751,717,521 | 144.8 | 0.0000079 | n.a. | −2.281 | TRIDC3BG072410 | Mitochondrial substrate carrier family protein | |
| BS00060666_51 | 3B | 751,945,425 | 144.8 | 0.0000079 | n.a. | −2.281 | TRIDC3BG072450 | Homeobox protein LUMINIDEPENDENS | |
| BS00105878_51 | 3B | 762,885,605 | n.a. | 0.0000079 | n.a. | −2.281 | TRIDC3BG074270 | Unknown function | |
| CAP12_rep_c4571_181 | 4B | 11,639,988 | n.a. | 0.0000079 | n.a. | −1.586 | TRIDC4BG003010 | Undescribed protein | |
| Kukri_rep_c79943_189 | 4B | 541,798,790 | n.a. | 0.0000079 | n.a. | −1.586 | TRIDC4BG046050 | Undescribed protein | |
| IAAV5683 | 5B | 518,614,625 | n.a. | 0.0000079 | n.a. | −3.308 | TRIDC5BG052520 | Proteasome subunit α-type-7-B | |
| Excalibur_c3165_730 | 5B | 590,819,414 | 131.2 | 0.0000079 | n.a. | −3.308 | TRIDC5BG062980 | BTB/POZ domain-containing protein | |
| IACX5390 | 5B | 600,810,891 | 136.7 | 0.0000079 | n.a. | −3.308 | TRIDC5BG064250 | Actin depolymerizing factor 6 | |
| wsnp_Ku_c11060_18147688 | 7B | 119,625,169 | 57.7 | 0.0000079 | n.a. | −3.308 | TRIDC7BG015000 | RNA-binding protein 47 | |
| Gli+Glu.16 | BobWhite_c19155_246 | 5A | 528,942,215 | 111.5 | 7.45 × 10−6 | 0.32 | 4.875 | TRIDC5AG047350 | Protein STAY-GREEN, chloroplastic |
| wsnp_Ex_c4026_7281501 | 5A | 529,160,493 | 111.5 | 7.45 × 10−6 | 0.32 | −4.875 | TRIDC5AG047410 | Two-component response regulator-like PRR95 | |
| CD-Related Peptides a | |||||||||
| IP2.16 | BS00041355_51 * | 1B | 496,196,589 | n.a. | 1.25 × 10−5 | n.a. | −1.669 | TRIDC1BG045920 | Unknown function |
| IP2.16 | wsnp_JD_c6331_7499060 * | 1B | 553,899,486 | n.a. | 6.36 × 10−5 | n.a. | 8.735 | TRIDC1BG052690 | Undescribed protein |
| IP2.16 | Excalibur_rep_c66322_448 * | 1B | 553,901,557 | n.a. | 1.08 × 10−5 | n.a. | 5.159 | TRIDC1BG052690 | Undescribed protein |
| IP2.16 | Excalibur_rep_c107047_605 * | 1B | 609,661,176 | 98.8 | 1.00 × 10−4 | n.a. | 3.146 | TRIDC1BG060060 | Choline/ethanolamine kinase |
| IP2.16 | Tdurum_contig96049_200 * | 1B | 610,156,207 | 98.8 | 1.00 × 10−4 | n.a. | 3.146 | TRIDC1BG060130 | Undescribed protein |
| IP2.16 | RAC875_c34012_983 * | 7B | 734,184,474 | n.a. | 6.09 × 10−5 | n.a. | 2.568 | TRIDC7BG073100 | 1,4-α-glucan branching enzyme GlgB |
| IP3.16 | BobWhite_c20073_382 | 1B | 584,479,982 | 156.3 | 2.94 × 10−14 | n.a. | 8.541 | Intergenic | |
| IP3.16 | RFL_Contig5290_1493 | 2B | 205,965,316 | 87.4 | 2.26 × 10−11 | n.a. | 5.350 | TRIDC2BG028530 | HXXXD-type acyl-transferase family protein |
| IP3.16 | BobWhite_rep_c66361_594 | 4A | 715,175,771 | 167.5 | 5.67 × 10−5 | n.a. | 2.875 | TRIDC4AG069440 | Protein CWC15 homolog |
| IP3.16 | BobWhite_c22086_444 | 6A | 8,529,327 | 4.3 | 2.69 × 10−7 | n.a. | 3.921 | TRIDC6AG002330 | COPII coat assembly protein SEC16 |
| IP3.16 | RAC875_c17559_3102 ** | 6B | 120,052,990 | 54.4 | 2.35 × 10−7 | n.a. | 2.858 | TRIDC6BG017260 | Endoglucanase 11 |
| IP3.16 | Excalibur_c92249_102 | 6B | 652,231,937 | 114.5 | 1.10 × 10−7 | n.a. | 1.389 | TRIDC6BG062810 | Unknown function |
| IP3.16 | Kukri_c39759_102 | 7B | 542,938,246 | n.a. | 3.14 × 10−6 | n.a. | 0.963 | TRIDC7BG048190 | Methionine-tRNA ligase |
| TPT.16 | BobWhite_c20073_382 | 1B | 584,479,982 | 156.3 | 6.50 × 10−6 | n.a. | −5.119 | TRIDC1BG056400 | Auxin response factor 4 |
| TPT.16 | CAP8_rep_c8022_270 | 2A | 33,508,142 | 35.6 | 3.06 × 10−6 | n.a. | 6.925 | TRIDC2AG007400 | Ribulose bisphosphate carboxylase small chain |
| TPT.16 | Excalibur_c92249_102 | 6B | 652,231,937 | 114.5 | 1.63 × 10−5 | n.a. | −6.045 | TRIDC6BG062810 | Unknown function |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taranto, F.; D’Agostino, N.; Catellani, M.; Laviano, L.; Ronga, D.; Milc, J.; Prandi, B.; Boukid, F.; Sforza, S.; Graziano, S.; et al. Characterization of Celiac Disease-Related Epitopes and Gluten Fractions, and Identification of Associated Loci in Durum Wheat. Agronomy 2020, 10, 1231. https://doi.org/10.3390/agronomy10091231
Taranto F, D’Agostino N, Catellani M, Laviano L, Ronga D, Milc J, Prandi B, Boukid F, Sforza S, Graziano S, et al. Characterization of Celiac Disease-Related Epitopes and Gluten Fractions, and Identification of Associated Loci in Durum Wheat. Agronomy. 2020; 10(9):1231. https://doi.org/10.3390/agronomy10091231
Chicago/Turabian StyleTaranto, Francesca, Nunzio D’Agostino, Marcello Catellani, Luca Laviano, Domenico Ronga, Justyna Milc, Barbara Prandi, Fatma Boukid, Stefano Sforza, Sara Graziano, and et al. 2020. "Characterization of Celiac Disease-Related Epitopes and Gluten Fractions, and Identification of Associated Loci in Durum Wheat" Agronomy 10, no. 9: 1231. https://doi.org/10.3390/agronomy10091231
APA StyleTaranto, F., D’Agostino, N., Catellani, M., Laviano, L., Ronga, D., Milc, J., Prandi, B., Boukid, F., Sforza, S., Graziano, S., Gullì, M., Visioli, G., Marmiroli, N., Badeck, F.-W., Minervini, A. P., Pecorella, I., Pecchioni, N., De Vita, P., & Francia, E. (2020). Characterization of Celiac Disease-Related Epitopes and Gluten Fractions, and Identification of Associated Loci in Durum Wheat. Agronomy, 10(9), 1231. https://doi.org/10.3390/agronomy10091231