Biochemical Response of Oakleaf Lettuce Seedlings to Different Concentrations of Some Metal(oid) Oxide Nanoparticles



Abstract
1. Introduction
2. Materials and Methods
2.1. Characteristics of Nanoparticles
2.2. Plant Material and Nanoparticle Application
2.3. Fresh and Dry Weight
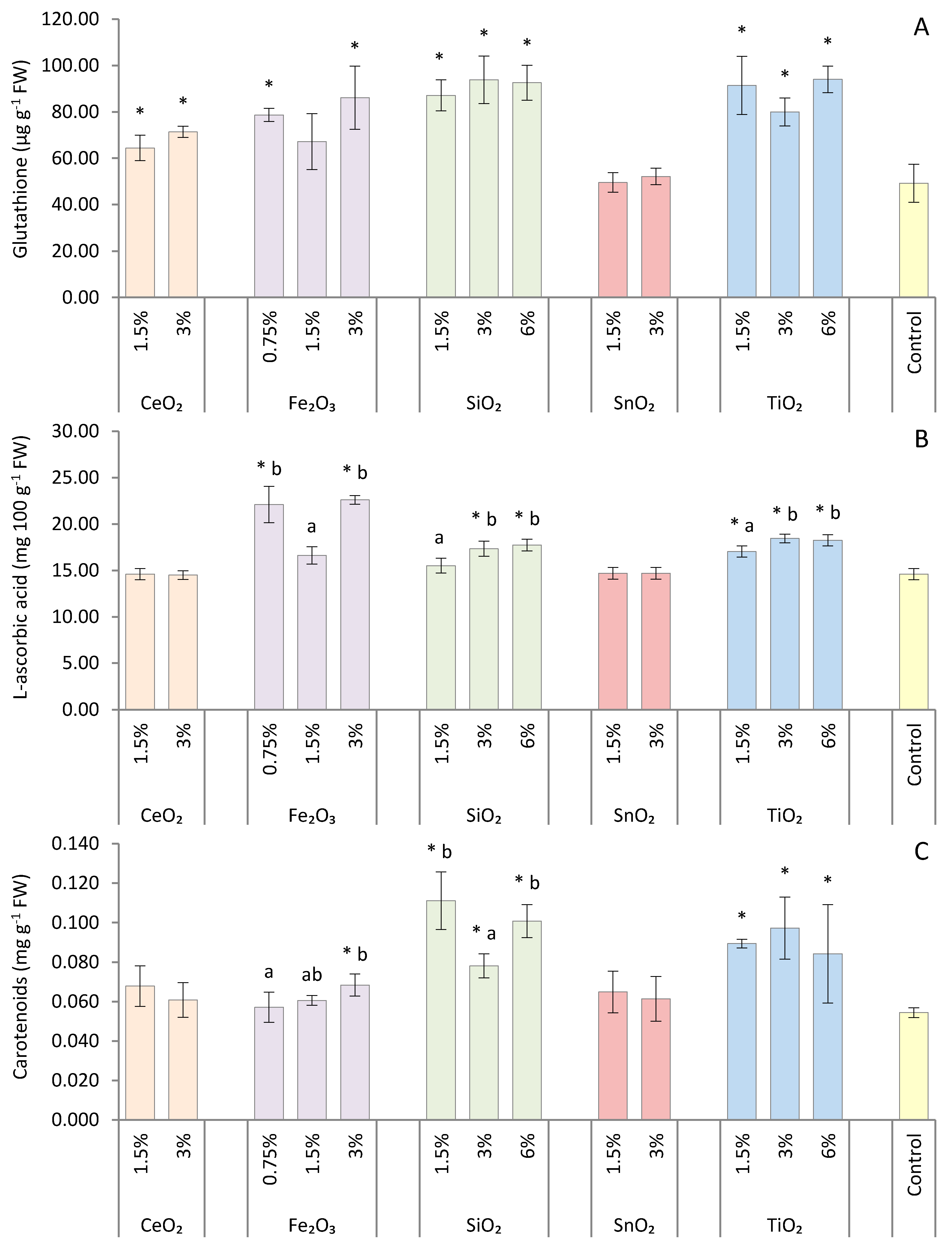
2.4. Chlorophyll and Carotenoid Content
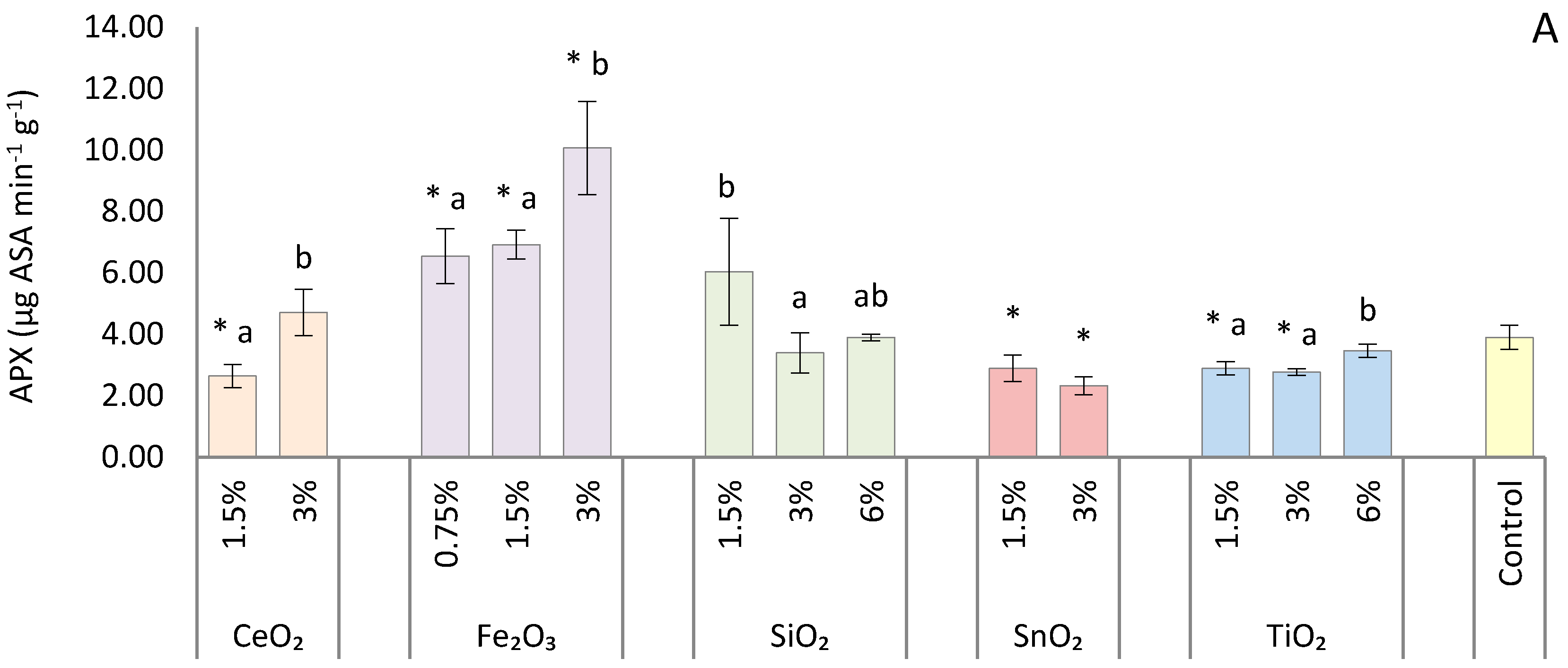
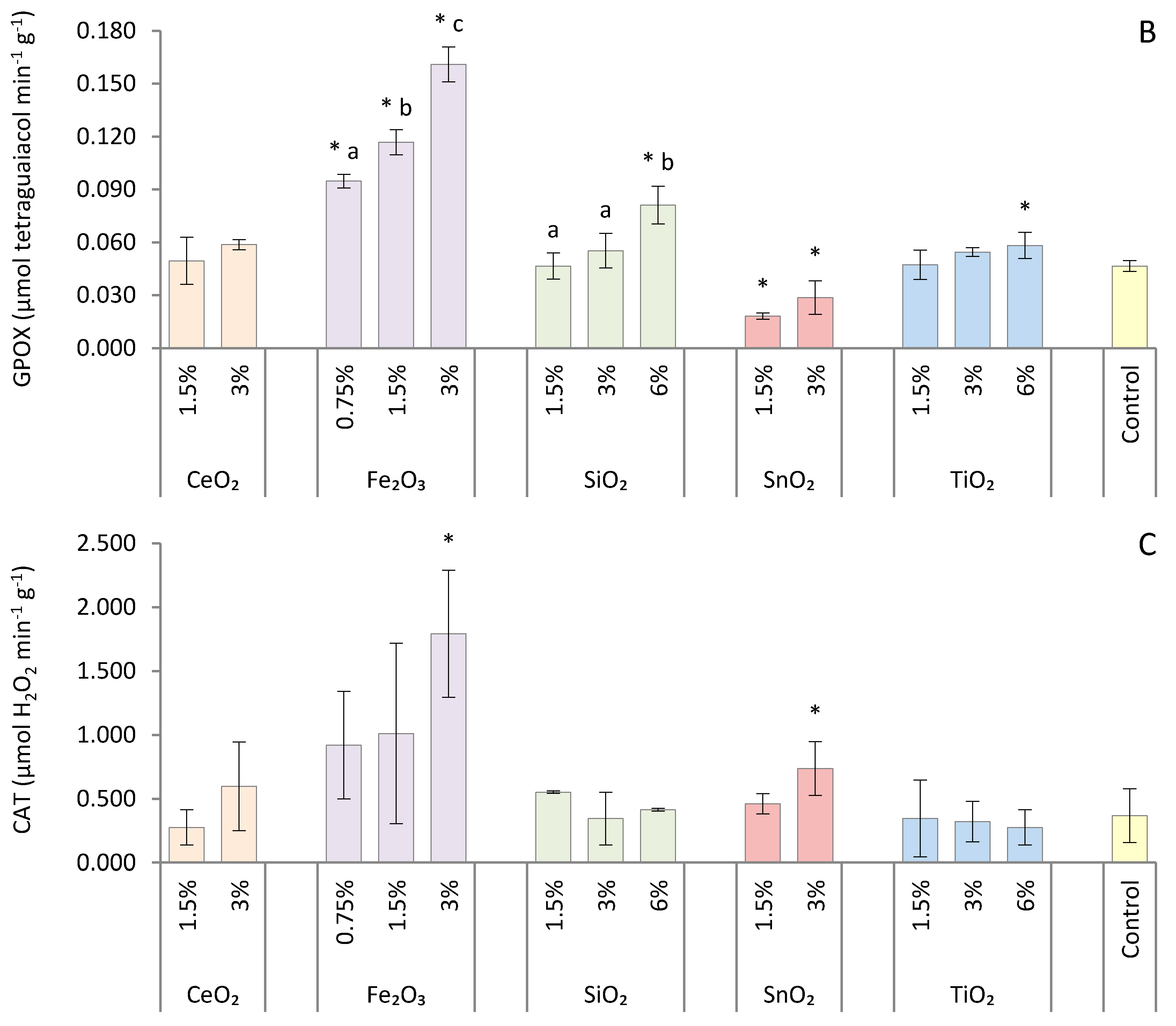
2.5. Antioxidant Enzyme Assays
2.6. Glutathione Content
2.7. L-ascorbic Acid Content
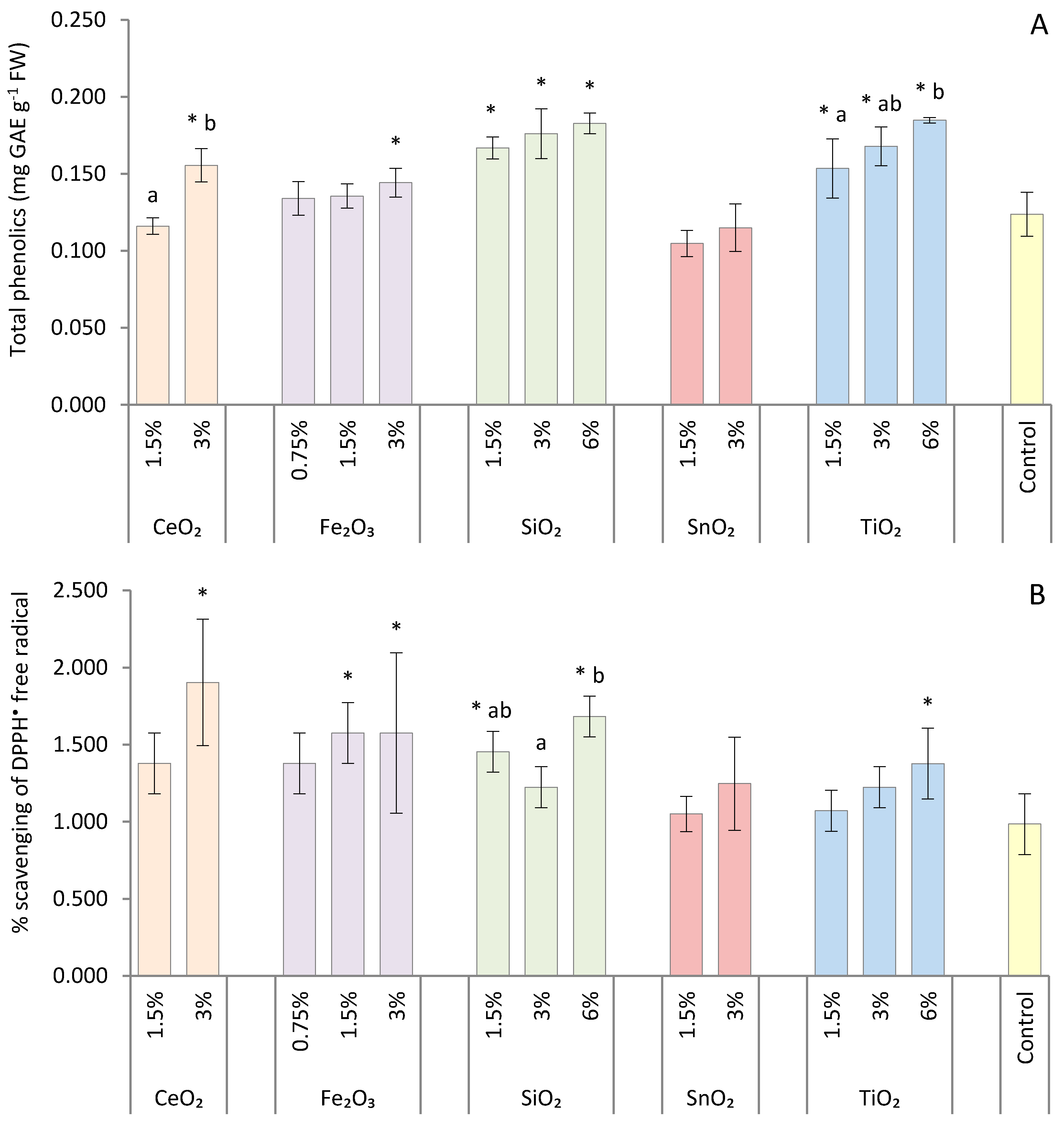
2.8. Total Phenolic Content
2.9. DPPH• Radical Scavenging Activity
2.10. Content of Ce, Fe, Si, Sn, and Ti
2.11. Statistical Analysis
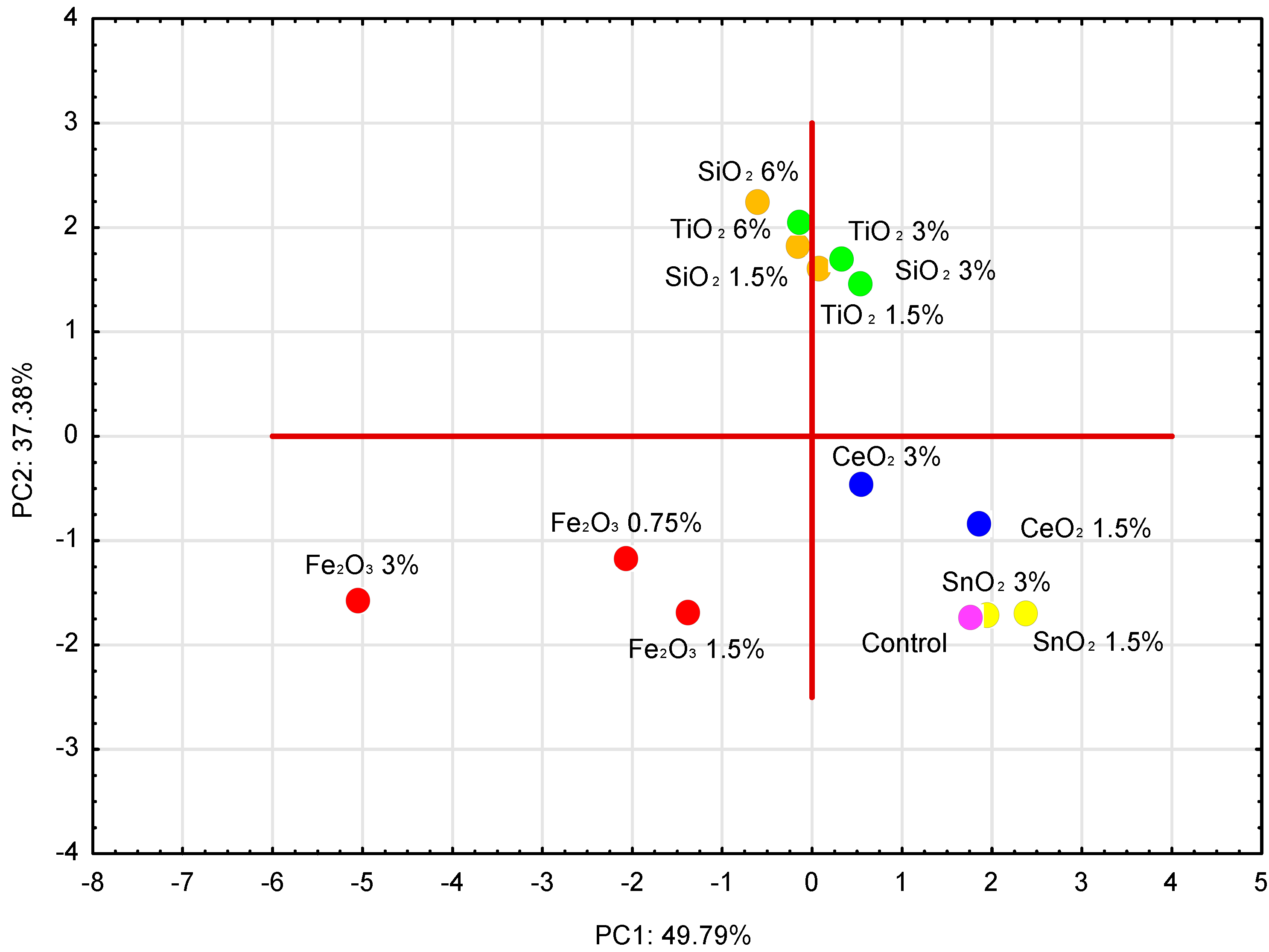
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sanzari, I.; Leone, A.; Ambrosone, A. Nanotechnology in plant science: To make a long story short. Front. Bioeng. Biotechnol. 2019, 7, 120. [Google Scholar] [CrossRef] [PubMed]
- Masarovičová, E.; Kráľová, K. Metal nanoparticles and plants. Ecol. Chem. Eng. S 2013, 20, 9–22. [Google Scholar]
- Nowack, B.; Bucheli, T.D. Occurrence, behavior and effects of nanoparticles in the environment. Environ. Pollut. 2007, 150, 5–22. [Google Scholar] [CrossRef]
- Khodakovskaya, M.V.; de Silva, K.; Biris, A.S.; Dervishi, E.; Villagarcia, H. Carbon nanotubes induce growth enhancement of tobacco cells. ACS Nano 2012, 6, 2128–2135. [Google Scholar] [CrossRef] [PubMed]
- Navarro, N.; Baun, A.; Behra, R.; Hartmann, N.B.; Filser, J.; Miao, A.-J.; Quigg, A.; Santschi, P.H.; Sigg, L. Environmental behavior and ecotoxicity of engineered nanoparticles to algae, plants, and fungi. Ecotoxicology 2008, 17, 372–386. [Google Scholar] [CrossRef] [PubMed]
- García-Gómez, C.; Fernández, M.D. Impacts of metal oxide nanoparticles on seed germination, plant growth and development. In Comprehensive Analytical Chemistry, Analysis, Fate, and Toxicity of Engineered Nanomaterials in Plants; Verma, S.K., Das, A.K., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; Volume 84, pp. 75–124. [Google Scholar]
- Milewska-Hendel, A.; Zubko, M.; Stróż, D.; Kurczyńska, E.U. Effect of nanoparticles surface charge on the Arabidopsis thaliana (L.) roots development and their movement into the root cells and protoplasts. Int. J. Mol. Sci. 2019, 20, 1650. [Google Scholar] [CrossRef] [PubMed]
- Rico, C.M.; Majumdar, S.; Duarte-Gardea, M.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Interaction of nanoparticles with edible plants and their possible implications in the food chain. J. Agric. Food Chem. 2011, 59, 3485–3498. [Google Scholar] [CrossRef]
- Li, J.; Hu, J.; Ma, C.; Wang, Y.; Wu, C.; Huang, J.; Xing, B. Uptake, translocation and physiological effects of magnetic iron oxide (γ-Fe2O3) nanoparticles in corn (Zea mays L.). Chemosphere 2016, 159, 326–334. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef]
- Zheng, L.; Hong, F.; Lu, S.; Liu, C. Effect of nano-TiO2 on strength of naturally aged seeds and growth of spinach. Biol. Trace Elem. Res. 2005, 104, 83–92. [Google Scholar] [CrossRef]
- Larue, C.; Castillo-Michel, H.; Sobanska, S.; Cécillon, L.; Bureau, S.; Barthès, V.; Ouerdane, L.; Carrière, M.; Sarret, G. Foliar exposure of the crop Lactuca sativa to silver nanoparticles: Evidence for internalization and changes in Ag speciation. J. Hazard. Mater. 2014, 264, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Larue, C.; Castillo-Michel, H.; Sobanska, S.; Trcera, N.; Sorieul, S.; Cécillon, L.; Ouerdane, L.; Legros, S.; Sarret, G. Fate of pristine TiO2 nanoparticles and aged paint-containing TiO2 nanoparticles in lettuce crop after foliar exposure. J. Hazard. Mater. 2014, 273, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 603, 591–592. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [PubMed]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Zhang, Z.; Pang, X.; Duan, X.; Ji, Z.L.; Jiang, Y. Role of peroxidase in anthocyanine degradation in litchi fruit pericarp. Food Chem. 2005, 90, 47–52. [Google Scholar] [CrossRef]
- Guri, A. Variation in glutathione and ascorbic acid content among selected cultivars of Phaseolus vulgaris prior to and after exposure to ozone. Can. J. Plant Sci. 1983, 63, 733–737. [Google Scholar] [CrossRef]
- Krełowska-Kułas, M. Badanie Jakości Produktów Spożywczych [The Study of Food Quality]; PWN: Warsaw, Poland, 1993. [Google Scholar]
- Djeridane, A.; Yousfi, M.; Nadjemi, B.; Boutassouna, D.; Stocker, P.; Vidal, N. Antioxidant activity of some Algerian medicinal plants extracts containing phenolic compound. Food Chem. 2006, 97, 654–660. [Google Scholar] [CrossRef]
- Molyneux, P. The use of the stable free radical diphenylpicrylhydrazyl (DPPH) for estimating antioxidant activity. Songklanakarin J. Sci. Technol. 2004, 26, 211–219. [Google Scholar]
- Barros, J.A.; de Souza, P.F.; Schiavo, D.; Nóbrega, J.A. Microwave-assisted digestion using diluted acid and base solutions for plant analysis by ICP OES. J. Anal. Atom. Spectrom. 2016, 31, 337–343. [Google Scholar] [CrossRef]
- Kalisz, A.; Sękara, A.; Smoleń, S.; Grabowska, A.; Gil, J.; Komorowska, M.; Kunicki, E. Survey of 17 elements, including rare earth elements, in chilled and non-chilled cauliflower cultivars. Sci. Rep. 2019, 9, 5416. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Qayyum, M.F.; Ok, Y.S.; Adrees, M.; Ibrahim, M.; Zia-Ur-Rehman, M.; Farid, M.; Abbas, F. Effect of metal and metal oxide nanoparticles on growth and physiology of globally important food crops: A critical review. J. Hazard. Mater. 2017, 322, 2–16. [Google Scholar] [CrossRef] [PubMed]
- Feizi, H.; Moghaddam, P.R.; Shahtahmassebi, N.; Fotovat, A. Assessment of concentrations of nano and bulk iron oxide particles on early growth of wheat (Triticum aestivum L.). Annu. Res. Rev. Biol. 2013, 3, 752–761. [Google Scholar]
- Jeyasubramanian, K.; Thoppeya, U.U.G.; Hikkua, G.S.; Selvakumara, N.; Subramaniab, A.; Krishnamoorthy, K. Enhancement in growth rate and productivity of spinach grown in hydroponics with iron oxide nanoparticles. RSC Adv. 2016, 6, 15451. [Google Scholar] [CrossRef]
- Servin, A.D.; Morales, M.I.; Castillo-Michel, H.; Hernandez-Viezcas, J.A.; Munoz, B.; Zhao, L.; Nunez, J.E.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Synchrotron verification of TiO2 accumulation in cucumber fruit: A possible pathway of TiO2 nanoparticle transfer from soil into the food chain. Environ. Sci. Technol. 2013, 47, 11592–11598. [Google Scholar] [CrossRef] [PubMed]
- Du, W.; Tan, W.; Peralta-Videa, J.B.; Gardea-Torresdey, J.L.; Ji, R.; Yin, Y.; Guo, H. Interaction of metal oxide nanoparticles with higher terrestrial plants: Physiological and biochemical aspects. Plant Physiol. Biochem. 2017, 110, 210–225. [Google Scholar] [CrossRef]
- Karimi, J.; Mohsenzadeh, S. Effects of silicon oxide nanoparticles on growth and physiology of wheat seedlings. Russ. J. Plant Physiol. 2016, 63, 119–123. [Google Scholar] [CrossRef]
- Sun, D.; Hussain, H.I.; Yi, Z.; Rookes, J.E.; Kong, L.; Cahill, D.M. Mesoporous silica nanoparticles enhance seedling growth and photosynthesis in wheat and lupin. Chemosphere 2016, 152, 81–91. [Google Scholar] [CrossRef]
- Wang, M.; Liu, X.; Hu, J.; Li, J.; Huang, J. Nano-ferric oxide promotes watermelon growth. J. Biomater. Nanobiotechnol. 2015, 6, 160–167. [Google Scholar] [CrossRef]
- Miller, G.W.; Jen Huang, I.; Welkiel, G.W.; Pushnik, J.C. Function of iron in plants with special emphasis on chloroplasts and photosynthetic activity. In Iron Nutrition in Soils and Plants; Abadia, J., Ed.; Kluwer Academic Publishers: Berlin, Germany, 1995; pp. 19–28. [Google Scholar]
- Briat, J.-F.; Dubos, C.; Gaymard, F. Iron nutrition, biomass production, and plant product quality. Trends Plant Sci. 2015, 20, 33–40. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, S.; Xu, M.; Xiao, L.; Dai, Z.; Li, J. The impacts of γ-Fe2O3 and Fe3O4 nanoparticles on the physiology and fruit quality of muskmelon (Cucumis melo) plants. Environ. Pollut. 2019, 249, 1011–1018. [Google Scholar] [CrossRef]
- Ghafari, H.; Razmjoo, J. Effect of foliar application of nano-iron oxidase, iron chelate and iron sulphate rates on yield and quality of wheat. Int. J. Agron. Plant Prod. 2013, 4, 2997–3003. [Google Scholar]
- Siddiqi, K.S.; Husen, A. Plant response to engineered metal oxide nanoparticles. Nanoscale Res. Lett. 2017, 12, 92. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, R.; Maali-Amiri, R.; Mantrib, N.L. Effect of TiO2 nanoparticles on oxidative damage and antioxidant defense systems in chickpea seedlings during cold stress. Russ. J. Plant Physiol. 2014, 61, 768–775. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Ann. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed]
- Jahani, S.; Saadatmand, S.; Mahmoodzadeh, H.; Khavari-Nejad, R.A. Effects of cerium oxide nanoparticles on biochemical and oxidative parameters in marigold leaves. Toxicol. Environ. Chem. 2018, 100, 677–692. [Google Scholar] [CrossRef]
- Yoganandham, S.T.; Ravindranath, R.R.S.; Sathyamoorthy, G.; Renuka, R.R.; Lakshminarayanan, A. Hematite nanoparticles: Synthesis, characterization and aquatic ecotoxicity effects. Res. J. Biotech. 2019, 14, 21–30. [Google Scholar]
- Tripathi, D.K.; Singh, S.; Singh, V.P.; Prasad, S.M.; Chauhan, D.K.; Dubey, N.K. Silicon nanoparticles more efficiently alleviate arsenate toxicity than silicon in maize cultiver and hybrid differing in arsenate tolerance. Front. Environ. Sci. 2016, 4, 46. [Google Scholar] [CrossRef]
- Rico, C.M.; Hong, J.; Morales, M.I.; Zhao, L.; Barrios, A.C.; Zhang, J.-Y.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Effect of cerium oxide nanoparticles on rice: A study involving the antioxidant defense system and in vivo fluorescence imaging. Environ. Sci. Technol. 2013, 47, 5635–5642. [Google Scholar] [CrossRef]
- Kim, Y.-H.; Khan, A.L.; Waqas, M.; Lee, I.-J. Silicon regulates antioxidant activities of crop plants under abiotic-induced oxidative stress: A review. Front Plant Sci. 2017, 8, 510. [Google Scholar] [CrossRef]
- Silva, S.; de Oliveira, J.M.P.F.; Dias, M.C.; Silva, A.M.S.; Santos, C. Antioxidant mechanisms to counteract TiO2-nanoparticles toxicity in wheat leaves and roots are organ dependent. J. Hazard. Mat. 2019, 380, 120889. [Google Scholar] [CrossRef] [PubMed]
- Tavallali, V. Effects of iron nano-complex and Fe-EDDHA on bioactive compounds and nutrient status of purslane plants. Int. Agrophys. 2018, 32, 411–419. [Google Scholar] [CrossRef]
- Sharifi-Rad, A.; Sharifi-Rad, M.; Teixeira da Silva, J.A. Morphological, physiological and biochemical responses of crops (Zea mays L., Phaseolus vulgaris L.), medicinal plants (Hyssopus officinalis L., Nigella sativa L.), and weeds (Amaranthus retroflexus L., Taraxacum officinale F. H. Wigg) exposed to SiO2 nanoparticles. J. Agric. Sci. Tech. 2016, 18, 1027–1040. [Google Scholar]
- Ma, C.; Chhikara, S.; Xing, B.; Musante, C.; White, J.C.; Dhankher, O.P. Physiological and molecular response of Arabidopsis thaliana (L.) to nanoparticle cerium and indium oxide exposure. ACS Sustain. Chem. Eng. 2013, 1, 768–778. [Google Scholar] [CrossRef]
- Farhangi-Abriz, S.; Torabian, S. Nano-silicon alters antioxidant activities of soybean seedlings under salt toxicity. Protoplasma 2018, 255, 953–962. [Google Scholar] [CrossRef] [PubMed]
- Vega, I.; Nikolic, M.; Pontigo, S.; Godoy, K.; de La Luz Mora, M.; Cartes, P. Silicon improves the production of high antioxidant or structural phenolic compounds in barley cultivars under aluminum stress. Agronomy 2019, 9, 388. [Google Scholar] [CrossRef]
- Ghorbanpour, M. Major essential oil constituents, total phenolics and flavonoids content and antioxidant activity of Salvia officinalis plant in response to nano-titanium dioxide. Ind. J. Plant Physiol. 2015, 20, 249–256. [Google Scholar] [CrossRef]
- Torabzadeh, D.; Hassanpour, H.; Asgarpanah, J.; Rezayian, M. Nanoparticles induced antioxidative compounds in Matricaria chamomilla. Iran. J. Plant Physiol. 2019, 9, 2955–2961. [Google Scholar]
- Zhang, Z.; He, X.; Zhang, H.; Ma, Y.; Zhang, P.; Ding, Y.; Zhao, Y. Uptake and distribution of ceria nanoparticles in cucumber plants. Metallomics 2011, 3, 816–822. [Google Scholar] [CrossRef]
- Le, V.N.; Rui, Y.; Gui, X.; Li, X.; Liu, S.; Ha, Y. Uptake, transport, distribution and bio-effects of SiO2 nanoparticles in Bt-transgenic cotton. J. Nanobiotechnol. 2014, 12, 50. [Google Scholar] [CrossRef]
- Jiang, F.; Shen, Y.; Ma, C.; Zhang, X.; Cao, W.; Rui, Y. Effects of TiO2 nanoparticles on wheat (Triticum aestivum L.) seedlings cultivated under super-elevated and normal CO2 conditions. PLoS ONE 2017, 12, e0178088. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MO-NPs and Concentration | Fresh Weight (g per shoot) | Dry Weight (g 100 g−1 FW) | Total Dry Weight (g per shoot) | Chlorophyll a (mg g−1 FW) | Chlorophyll b (mg g−1 FW) | Chlorophyll a + b (mg g−1 FW) | Chl a:Chl b Ratio | Car:Chls Ratio | |
|---|---|---|---|---|---|---|---|---|---|
| CeO2 | 1.5% | 2.850 ± 0.262 | 3.309 ± 0.226 | 0.094 ± 0.011 | 0.322 ± 0.036 | 0.134 ± 0.011 | 0.456 ± 0.041 | 2.403 ± 0.290 | 0.149 ± 0.016 |
| 3% | 2.851 ± 0.263 | 3.596 ± 0.090 | 0.103 ± 0.012 | 0.301 ± 0.033 | 0.125 ± 0.019 | 0.426 ± 0.046 | 2.408 ± 0.282 | 0.143 ± 0.017 | |
| Fe2O3 | 0.75% | 2.821 ± 0.315 | 3.957 ± 0.196 *a | 0.111 ± 0.008 | 0.272 ± 0.009 a | 0.105 ± 0.007 a | 0.377 ± 0.007 a | 2.591 ± 0.231 | 0.152 ± 0.019 |
| 1.5% | 2.629 ± 0.476 | 3.999 ± 0.073 *a | 0.105 ± 0.017 | 0.290 ± 0.006 a | 0.117 ± 0.002 ab | 0.407 ± 0.005 a | 2.479 ± 0.096 | 0.149 ± 0.005 | |
| 3% | 2.208 ± 0.386 * | 4.567 ± 0.276 *b | 0.100 ± 0.015 | 0.333 ± 0.029 *b | 0.125 ± 0.009 b | 0.458 ± 0.038 *b | 2.664 ± 0.053 | 0.149 ± 0.004 | |
| SiO2 | 1.5% | 3.305 ± 0.407 | 3.471 ± 0.015 | 0.115 ± 0.015 | 0.431 ± 0.025 *b | 0.137 ± 0.008 *b | 0.568 ± 0.034 *b | 3.146 ± 0.048 * | 0.195 ± 0.016 *b |
| 3% | 3.325 ± 0.057 | 3.603 ± 0.045 | 0.120 ± 0.001 | 0.335 ± 0.023 *a | 0.107 ± 0.006 a | 0.442 ± 0.029 a | 3.131 ± 0.076 * | 0.177 ± 0.002 *a | |
| 6% | 3.169 ± 0.325 | 3.637 ± 0.233 | 0.116 ± 0.019 | 0.431 ± 0.043 *b | 0.136 ± 0.014 *b | 0.567 ± 0.057 *b | 3.169 ± 0.075 * | 0.178 ± 0.003 *a | |
| SnO2 | 1.5% | 3.069 ± 0.121 | 3.305 ± 0.099 | 0.102 ± 0.007 | 0.304 ± 0.037 | 0.117 ± 0.008 | 0.421 ± 0.044 | 2.598 ± 0.167 | 0.154 ± 0.009 |
| 3% | 2.993 ± 0.325 | 3.229 ± 0.008 | 0.097 ± 0.008 | 0.285 ± 0.046 | 0.113 ± 0.013 | 0.398 ± 0.059 | 2.522 ± 0.118 | 0.154 ± 0.006 | |
| TiO2 | 1.5% | 3.294 ± 0.558 | 3.530 ± 0.104 | 0.116 ± 0.017 | 0.384 ± 0.002 | 0.135 ± 0.020 | 0.519 ± 0.018 | 2.844 ± 0.407 | 0.172 ± 0.002 * |
| 3% | 3.413 ± 0.581 | 3.602 ± 0.196 | 0.123 ± 0.022 | 0.424 ± 0.064 * | 0.137 ± 0.021 | 0.561 ± 0.084 * | 3.095 ± 0.087 * | 0.173 ± 0.010 * | |
| 6% | 3.131 ± 0.370 | 3.314 ± 0.194 | 0.104 ± 0.016 | 0.380 ± 0.117 | 0.125 ± 0.042 | 0.505 ± 0.159 | 3.040 ± 0.111 * | 0.167 ± 0.004 * | |
| Control | 3.027 ± 0.213 | 3.347 ± 0.234 | 0.101 ± 0.004 | 0.270 ± 0.018 | 0.110 ± 0.015 | 0.380 ± 0.032 | 2.455 ± 0.191 | 0.143 ± 0.007 | |
| MO-NPs and Concentration | Element Content | |
|---|---|---|
| Ce | ||
| Control | 0.13 ± 0.02 | |
| CeO2 | 1.5% | 512.74 ± 49.49 *a |
| CeO2 | 3% | 712.94 ± 128.10 *b |
| Fe | ||
| Control | 33.16 ± 28.52 | |
| Fe2O3 | 0.75% | 936.42 ± 117.34 a |
| Fe2O3 | 1.5% | 1566.10 ± 195.63 *a |
| Fe2O3 | 3% | 5642.71 ± 1390.65 *b |
| Si | ||
| Control | 59.13 ± 12.37 | |
| SiO2 | 1.5% | 62.42 ± 0.40 |
| SiO2 | 3% | 92.09 ± 51.54 |
| SiO2 | 6% | 97.68 ± 1.69 |
| Sn | ||
| Control | 0.97 ± 0.13 | |
| SnO2 | 1.5% | 208.87 ± 2.50 *a |
| SnO2 | 3% | 323.08 ± 94.94 *b |
| Ti | ||
| Control | 1.44 ± 0.20 | |
| TiO2 | 1.5% | 208.44 ± 36.39 a |
| TiO2 | 3% | 460.80 ± 89.45 *a |
| TiO2 | 6% | 932.06 ± 445.18 *b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jurkow, R.; Sękara, A.; Pokluda, R.; Smoleń, S.; Kalisz, A. Biochemical Response of Oakleaf Lettuce Seedlings to Different Concentrations of Some Metal(oid) Oxide Nanoparticles. Agronomy 2020, 10, 997. https://doi.org/10.3390/agronomy10070997
Jurkow R, Sękara A, Pokluda R, Smoleń S, Kalisz A. Biochemical Response of Oakleaf Lettuce Seedlings to Different Concentrations of Some Metal(oid) Oxide Nanoparticles. Agronomy. 2020; 10(7):997. https://doi.org/10.3390/agronomy10070997
Chicago/Turabian StyleJurkow, Rita, Agnieszka Sękara, Robert Pokluda, Sylwester Smoleń, and Andrzej Kalisz. 2020. "Biochemical Response of Oakleaf Lettuce Seedlings to Different Concentrations of Some Metal(oid) Oxide Nanoparticles" Agronomy 10, no. 7: 997. https://doi.org/10.3390/agronomy10070997
APA StyleJurkow, R., Sękara, A., Pokluda, R., Smoleń, S., & Kalisz, A. (2020). Biochemical Response of Oakleaf Lettuce Seedlings to Different Concentrations of Some Metal(oid) Oxide Nanoparticles. Agronomy, 10(7), 997. https://doi.org/10.3390/agronomy10070997