Allelopathic Potential of Teff Varieties and Effect on Weed Growth

 ,
,
Abstract
1. Introduction
- There are differences in allelopathic activity between teff varieties.
- Emergence and allelopathic activity are the two most important traits for teff’s competitive ability against weeds.
2. Materials and Methods
2.1. Plant Materials
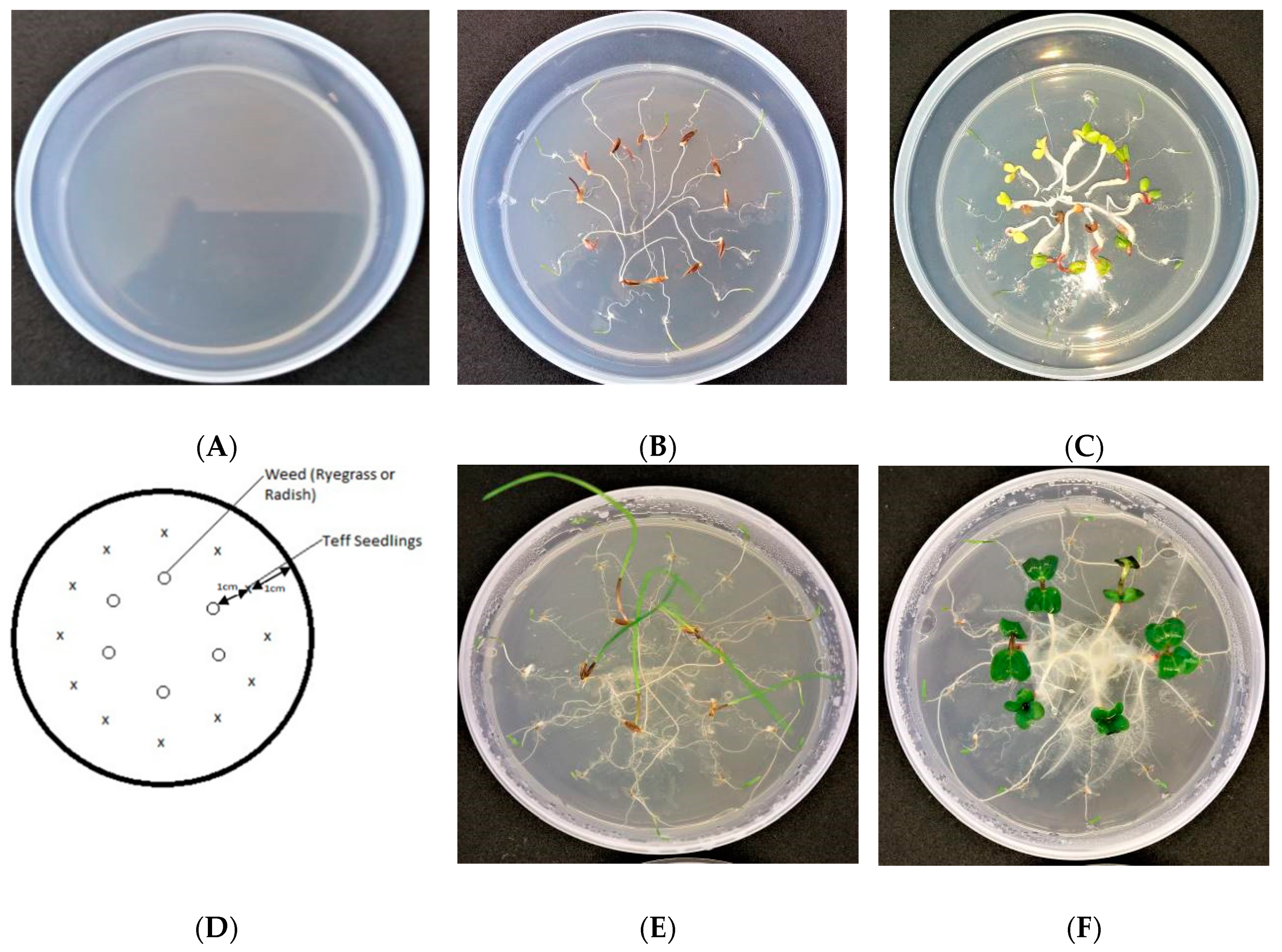
2.2. Part I: Detecting the Potential Allelopathic Activity (PAA) of Teff Varieties (Bioassay Experiment)
2.2.1. Seed Germination of Teff Varieties and Model Weeds
2.2.2. Transplanting of Seedlings of Teff and Model Weeds into Water Agar and Their Incubation in the Growth Chamber (Bioassay)
2.2.3. Root Analysis and Weight Measurements
2.3. Part II: Contribution of PAA and Other Teff Agronomic Traits on Weed Growth Dynamics
2.4. Data Analysis
3. Results
3.1. Part I: Detecting the Potential Allelopathic Activity (PAA) of Teff Varieties (Bioassay Experiment)
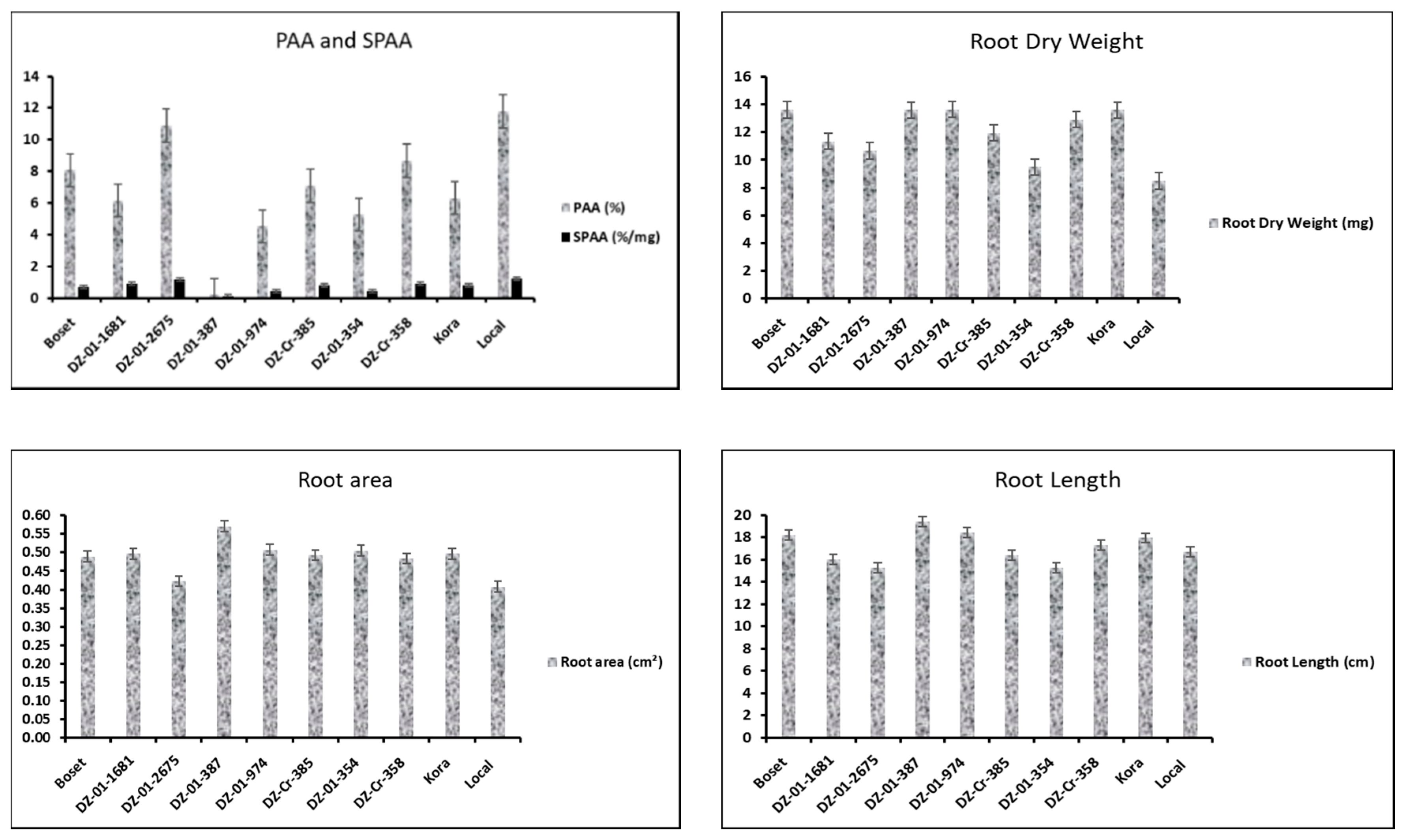
3.1.1. Allelopathic Effect on Ryegrass
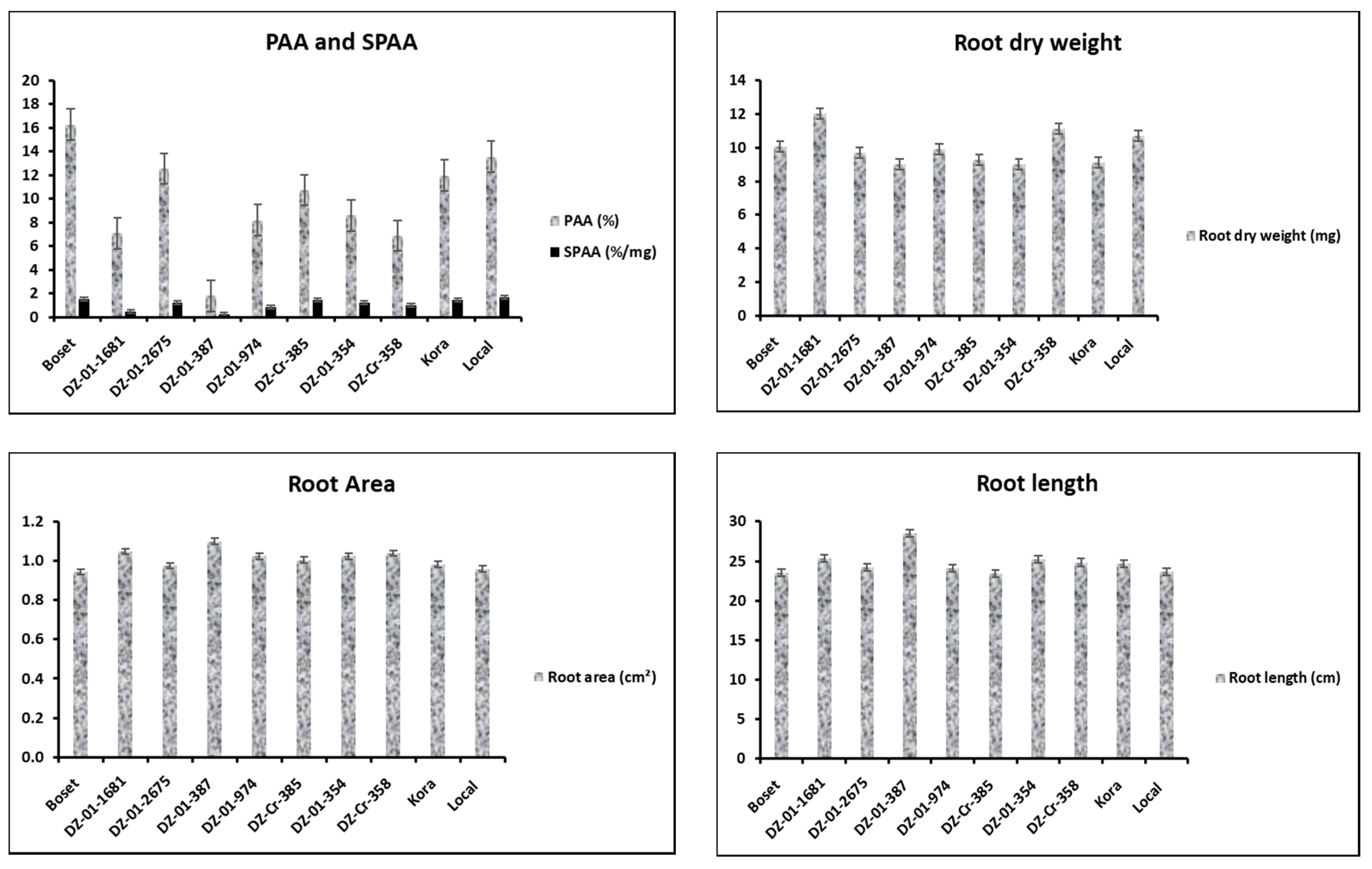
3.1.2. Allelopathic Effect on Radish
3.2. Part II: Contribution of PAA and Other Teff Agronomic Traits on Weed Growth Dynamics
Relationships between Traits of Teff Varieties and Weed Assessments
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Root Length (cm) | Root Area (cm2) | Root Diameter (mm) | Root Volume (cm3) | Root Dry Weight (mg) | |
|---|---|---|---|---|---|
| Root length (cm) | 1 | 0.9239 | −0.5848 | 0.6483 | 0.57582 |
| 0.0001 | 0.0758 | 0.0426 | 0.0815 | ||
| Root area (cm2) | 0.92394 | 1 | −0.3229 | 0.7592 | 0.48459 |
| 0.0001 | 0.3627 | 0.0109 | 0.1558 | ||
| Root diameter (mm) | −0.58483 | −0.323 | 1 | −0.2644 | −0.16693 |
| 0.0758 | 0.3627 | 0.4603 | 0.6448 | ||
| Root volume (cm3) | 0.64831 | 0.7592 | −0.2644 | 1 | 0.44691 |
| 0.0426 | 0.0109 | 0.4603 | 0.1954 | ||
| Root (dry weight (mg) | 0.57582 | 0.4846 | −0.1669 | 0.4469 | 1 |
| 0.0815 | 0.1558 | 0.6448 | 0.1954 |
| Root Length (cm) | Root Area (cm2) | Root Diameter (mm) | Root Volume (cm3) | Root (Dry Weight (mg) | |
|---|---|---|---|---|---|
| Root length (cm) | 1 | 0.8388 | −0.8121 | 0.3109 | −0.1913 |
| 0.0024 | 0.0043 | 0.3819 | 0.5966 | ||
| Root area (cm2) | 0.83882 | 1 | −0.3893 | 0.7674 | 0.01436 |
| 0.0024 | 0.2662 | 0.0096 | 0.9686 | ||
| Root diameter (mm) | −0.81207 | −0.389 | 1 | 0.2546 | 0.43235 |
| 0.0043 | 0.2662 | 0.4777 | 0.2121 | ||
| Root volume (cm3) | 0.31092 | 0.7674 | 0.2546 | 1 | 0.19908 |
| 0.3819 | 0.0096 | 0.4777 | 0.5814 | ||
| Root dry weight (mg) | −0.19126 | 0.0144 | 0.4324 | 0.1991 | 1 |
| 0.5966 | 0.9686 | 0.2121 | 0.5814 |
| Emergence | Heading | Maturity | Plant Height | Tiller No. Per Plant | Biomass Yield (kg/ha) | PAArye | SPAArye | PAArad | SPAArad | |
|---|---|---|---|---|---|---|---|---|---|---|
| Emergence | 1 | 0.56923 | 0.58938 | 0.24164 | 0.56447 | −0.79402 | −0.03352 | −0.124 | −0.3327 | −0.00824 |
| 0.0859 | 0.073 | 0.5012 | 0.0891 | 0.0061 | 0.9268 | 0.7329 | 0.3475 | 0.982 | ||
| Heading | 0.56923 | 1 | 0.93377 | 0.83025 | 0.59615 | 0.00091 | −0.26368 | −0.23276 | −0.5653 | −0.149 |
| 0.0859 | <0.0001 | 0.0029 | 0.0689 | 0.998 | 0.4617 | 0.5175 | 0.0886 | 0.6812 | ||
| Maturity | 0.58938 | 0.93377 | 1 | 0.73715 | 0.65183 | −0.0289 | −0.08545 | 0.00143 | −0.4591 | −0.07745 |
| 0.073 | <0.0001 | 0.015 | 0.0411 | 0.9368 | 0.8144 | 0.9969 | 0.182 | 0.8316 | ||
| Plant height | 0.24164 | 0.83025 | 0.73715 | 1 | 0.43478 | 0.20518 | −0.00891 | −0.04849 | −0.2027 | 0.14782 |
| 0.5012 | 0.0029 | 0.015 | 0.2092 | 0.5696 | 0.9805 | 0.8942 | 0.5743 | 0.6836 | ||
| Tiller no. per plant | 0.56447 | 0.59615 | 0.65183 | 0.43478 | 1 | −0.32473 | −0.02873 | 0.05773 | −0.5719 | −0.41598 |
| 0.0891 | 0.0689 | 0.0411 | 0.2092 | 0.3599 | 0.9372 | 0.8741 | 0.0841 | 0.2318 | ||
| Biomass yield (kg/ha) | −0.79402 | 0.00091 | −0.0289 | 0.20518 | −0.32473 | 1 | −0.16876 | −0.01427 | 0.03512 | −0.07146 |
| 0.0061 | 0.998 | 0.9368 | 0.5696 | 0.3599 | 0.6412 | 0.9688 | 0.9233 | 0.8445 | ||
| PAArye | −0.03352 | −0.26368 | −0.08545 | −0.00891 | −0.02873 | −0.16876 | 1 | 0.94648 | 0.75023 | 0.70771 |
| 0.9268 | 0.4617 | 0.8144 | 0.9805 | 0.9372 | 0.6412 | <0.0001 | 0.0124 | 0.022 | ||
| SPAArye | −0.124 | −0.23276 | 0.00143 | −0.04849 | 0.05773 | −0.01427 | 0.94648 | 1 | 0.61775 | 0.5442 |
| 0.7329 | 0.5175 | 0.9969 | 0.8942 | 0.8741 | 0.9688 | <0.0001 | 0.057 | 0.1039 | ||
| PAArad | −0.33272 | −0.5653 | −0.45908 | −0.20272 | −0.5719 | 0.03512 | 0.75023 | 0.61775 | 1 | 0.8805 |
| 0.3475 | 0.0886 | 0.182 | 0.5743 | 0.0841 | 0.9233 | 0.0124 | 0.057 | 0.0008 | ||
| SPAArad | −0.00824 | −0.149 | −0.07745 | 0.14782 | −0.41598 | −0.07146 | 0.70771 | 0.5442 | 0.8805 | 1 |
| 0.982 | 0.6812 | 0.8316 | 0.6836 | 0.2318 | 0.8445 | 0.022 | 0.1039 | 0.0008 |
References
- Ketema, S. Tef (Eragrostis tef (Zucc.) Trotter): Promoting the Conservation and Use of Underutilized and Neglected Crops; Institute of Plant Genetics and Crop Plant Research, Gatersleben/International Plant Genetic Resources Institute: Rome, Italy, 1997; Volume 12. [Google Scholar]
- CSA. Agricutlrual Sample Survey 2015/2016 (2008E.C): Report on Area, Production and Farm Management Practice on Belg Season Crops for Private Peasant Holdings; CENTRAL STATISTICAL AGENCY: Addis Ababa, Ethiopia, 2016; Volume 5, p. 131.
- Assefa, K.; Cannarozzi, G.; Girma, D.; Kamies, R.; Chanyalew, S.; Plaza-Wüthrich, S.; Blösch, R.; Rindisbacher, A.; Rafudeen, S.; Tadele, Z. Genetic diversity in tef [Eragrostis tef (Zucc.) Trotter]. Front. Plant Sci. 2015, 6, 177. [Google Scholar] [CrossRef] [PubMed]
- Baye, K. Teff: Nutrient Composition and Health Benefits; International Food Policy Research Institute (IFPRI): Washington, DC, USA, 2014. [Google Scholar]
- Daniel, T.; Teferi, A.; Tesfaye, W. Evaluation of Improved Varieties of Teff in West Belessa, Northern Ethiopia. Rev. Plant Sci. 2016, 3, 1–6. [Google Scholar] [CrossRef]
- IFPRI. Atlas of the Ethiopian Rural Economy; International Food Policy Research Institute and Central Statistical Agency: Addis Ababa, Ethiopia, 2006; pp. 1–108. [Google Scholar] [CrossRef]
- Teklu, Y.; Tefera, H. Genetic improvement in grain yield potential and associated agronomic traits of tef (Eragrostis tef). Euphytica 2005, 141, 247–254. [Google Scholar] [CrossRef]
- Assefa, A.; Liben, M.; Tadesse, T.; Yeshalem, B. The effect of tillage frequency and weed control on yield of tef (Eragrostis tef) in Yielmana-Densa Area, northwestern Ethiopia. East Afr. J. Sci. 2008, 2, 35–40. [Google Scholar] [CrossRef][Green Version]
- Assefa, K.; Tadele, Z.; Chanyalew, S. (Eds.) Achievements and Prospects of Tef Improvement: Proceedings of the Second International Workshop, November 7–9, 2011, Debre Zeit, Ethiopia; Universität Bern: Bern, Swiss, 2013. [Google Scholar]
- Balesh, T.; Aune, J.B.; Johnsen, F.H.; Vanlauwe, B. The prospects of reduced tillage in tef (Eragrostis tef Zucca) in Gare Arera, West Shawa Zone of Oromiya, Ethiopia. Soil Tillage Res. 2008, 99, 58–65. [Google Scholar] [CrossRef]
- Haftamu, G.G.; Aune, J.B.; Netland, J.; Eklo, O.M.; Torp, T.; Brandsaeter, L.O. Weed competitive ability of teff (Eragrostis tef (Zucc.) Trotter) varieties. Agronomy 2020, 10, 108. [Google Scholar] [CrossRef]
- van der Meulen, A.; Chauhan, B.S. A review of weed management in wheat using crop competition. Crop Prot. 2017, 95, 38–44. [Google Scholar] [CrossRef]
- Powles, S.B. Evolved glyphosate-resistant weeds around the world: Lessons to be learnt. Pest Manag. Sci. 2008, 64, 360–365. [Google Scholar] [CrossRef]
- Powles, S.B. Evolution in action: Glyphosate-resistant weeds threaten world crops. Outlooks Pest Manag. 2008, 19, 256–259. [Google Scholar] [CrossRef]
- Andrew, I.K.S.; Storkey, J.; Sparkes, D.L. A review of the potential for competitive cereal cultivars as a tool in integrated weed management. Weed Res. 2015, 55, 239–248. [Google Scholar] [CrossRef]
- Grace, J.; Tilman, D. Perspectives on plant competition: Some introductory remarks. In Perspectives on Plant Competition; Elsevier: Amsterdam, The Netherlands, 1990; pp. 3–7. [Google Scholar]
- Willis, R.J. The History of Allelopathy; Springer: Dordrecht, The Netherlands, 2007; p. 315. [Google Scholar]
- Weston, L.A.; Duke, S.O. Weed and Crop Allelopathy. Crit. Rev. Plant Sci. 2003, 22, 367–389. [Google Scholar] [CrossRef]
- Lisanework, N.; Michelsen, A. Allelopathy in agroforestry systems: The effects of leaf extracts of Cupressus lusitanica and threeEucalyptus spp. on four Ethiopian crops. Agrofor. Syst. 1993, 21, 63–74. [Google Scholar] [CrossRef]
- Tefera, T. Allelopathic effects of Parthenium hysterophorus extracts on seed germination and seedling growth of Eragrostis tef. J. Agron. Crop Sci. 2002, 188, 306–310. [Google Scholar] [CrossRef]
- Chou, C.-H. Roles of Allelopathy in Plant Biodiversity and Sustainable Agriculture. Crit. Rev. Plant Sci. 1999, 18, 609–636. [Google Scholar] [CrossRef]
- Qasem, J.R.; Foy, C.L. Weed Allelopathy, Its Ecological Impacts and Future Prospects. J. Crop Prod. 2001, 4, 43–119. [Google Scholar] [CrossRef]
- Singh, H.P.; Batish, D.R.; Kohli, R.K. Allelopathic Interactions and Allelochemicals: New Possibilities for Sustainable Weed Management. Crit. Rev. Plant Sci. 2003, 22, 239–311. [Google Scholar] [CrossRef]
- Bertholdsson, N.O. Early vigour and allelopathy – two useful traits for enhanced barley and wheat competitiveness against weeds. Weed Res. 2005, 45, 94–102. [Google Scholar] [CrossRef]
- Weih, M.; Didon, U.M.E.; Rönnberg-Wästljung, A.C.; Björkman, C. Integrated agricultural research and crop breeding: Allelopathic weed control in cereals and long-term productivity in perennial biomass crops. Agric. Syst. 2008, 97, 99–107. [Google Scholar] [CrossRef]
- Wu, H.; Pratley, J.; Lemerle, D.; Haig, T. Evaluation of seedling allelopathy in 453 wheat (Triticum aestivum) accessions against annual ryegrass (Lolium rigidum) by the equal-compartment-agar method. Aust. J. Agric. Res. 2000, 51, 937–944. [Google Scholar] [CrossRef]
- Campbell, L.G.; Snow, A.A.; Sweeney, P.M.; Ketner, J.M. Rapid evolution in crop-weed hybrids under artificial selection for divergent life histories. Evol. Appl. 2009, 2, 172–186. [Google Scholar] [CrossRef]
- Dennis, C.O.; Jose, V.F.; Nikol, H. Weed Control and Radish (Raphanus sativus) Response to S-metolachlor on Organic Soils. HortScience 2016, 51, 79–83. [Google Scholar]
- Haugland, E.; Brandsaeter, L.O. Experiments on bioassay sensitivity in the study of allelopathy. J. Chem. Ecol. 1996, 22, 1845–1859. [Google Scholar] [CrossRef] [PubMed]
- Duke, S.O. Proving Allelopathy in Crop–Weed Interactions. Weed Sci. 2015, 63, 121–132. [Google Scholar] [CrossRef]
- Bertholdsson, N.-O. Variation in allelopathic activity over 100 years of barley selection and breeding. Weed Res. 2004, 44, 78–86. [Google Scholar] [CrossRef]
- Inderjit; Dakshini, K.M.M.; Einhellig, F.A. Allelopathy; American Chemical Society: Washington, DC, USA, 1994; Volume 582, p. 396. [Google Scholar]
- Inderjit; Duke, S.O. Ecophysiological aspects of allelopathy. Planta 2003, 217, 529–539. [Google Scholar] [CrossRef]
- Walker, T.S.; Bais, H.P.; Grotewold, E.; Vivanco, J.M. Root Exudation and Rhizosphere Biology. Plant Physiol. 2003, 132, 44. [Google Scholar] [CrossRef]
- Artico, L.L.; Kömmling, G.; Clarindo, W.R.; Menezes, A.P.S. Cytotoxic, Genotoxic, Mutagenic, and Phytotoxic Effects of the Extracts from Eragrostis plana Nees, 1841 (Poaceae), Grown in a Coal-Contaminated Region. Water Air Soil Pollut. 2020, 231, 81. [Google Scholar] [CrossRef]
- Batish, D.R.; Singh, H.P.; Kaur, S. Crop Allelopathy and Its Role in Ecological Agriculture. J. Crop Prod. 2001, 4, 121–161. [Google Scholar] [CrossRef]
- Belz, R.G.; Reinhardt, C.F.; Foxcroft, L.C.; Hurle, K. Residue allelopathy in Parthenium hysterophorus L.—Does parthenin play a leading role? Crop Prot. 2007, 26, 237–245. [Google Scholar] [CrossRef]
- Goodall, J.; Witkowski, E.T.F.; Ammann, S.; Reinhardt, C. Does allelopathy explain the invasiveness of Campuloclinium macrocephalum (pompom weed) in the South African grassland biome? Biol. Invasions 2010, 12, 3497–3512. [Google Scholar] [CrossRef]
- Laan, M.v.d.; Reinhardt, C.; Belz, R.; Truter, W.; Foxcroft, L.; Hurle, K. Interference potential of the perennial grasses Eragrostis curvula, Panicum maximum and Digitaria eriantha with Parthenium hysterophorus. TG: Trop. Grassl. 2008, 42, 88. [Google Scholar]
- Latif, S. Performance and Secondary Chemistry of Selected Annual Pasture Legumes in Southeastern Australia. Ph.D. Thesis, Charles Sturt University Wagga Wagga, Wagga Wagga, NSW, Australia, 2019. [Google Scholar]
- Siyar, S.; Majeed, A.; Muhammad, Z.; Ali, H.; Inayat, N. Allelopathic effect of aqueous extracts of three weed species on the growth and leaf chlorophyll content of bread wheat. Acta Ecol. Sin. 2019, 39, 63–68. [Google Scholar] [CrossRef]
- Sponchiado, D. Estudo alelopático e fitoquímico de Eragrostis lugens Nees.(POACEAE). Master’s Thesis, Universidade Federal De Santa Maria, Santa Maria, Brasil, 2019. [Google Scholar]
- Yang, M.; Zhao, Y.; Yang, H.; Shen, Y.; Zhang, X. Suppression of weeds and weed seeds in the soil by stubbles and no-tillage in an arid maize-winter wheat-common vetch rotation on the Loess Plateau of China. J. Arid Land 2018, 10, 809–820. [Google Scholar] [CrossRef]
- EMAaRD. Crop Variety Issue No.12: Summary of Released/Registered Varieties of Crops; Ethiopian Ministry of Agriculture and Rural Development: Addis Ababa, Ethiopia, 2014; Volume 12. [Google Scholar]
- Kebebew, A.; Sherif, A.; Getachew, B.; Gizaw, M.; Hailu, T.; Mark, E.S. Quncho: The first popular tef variety in Ethiopia. Int. J. Agric. Sustain. 2011, 9, 25–34. [Google Scholar] [CrossRef]
- Christensen, S. Weed suppression ability of spring barley varieties. Weed Res. 1995, 35, 241–247. [Google Scholar] [CrossRef]
- Lemerle, D.; Verbeek, B.; Cousens, R.D.; Coombes, N.E. The potential for selecting wheat varieties strongly competitive against weeds. Weed Res. 1996, 36, 505–513. [Google Scholar] [CrossRef]
- Korres, N.E.; Froud-Williams, R.J. Effects of winter wheat cultivars and seed rate on the biological characteristics of naturally occurring weed flora. Weed Res. 2002, 42, 417–428. [Google Scholar] [CrossRef]
- Bajwa, A.A.; Walsh, M.; Chauhan, B.S. Weed management using crop competition in Australia. Crop Prot. 2017, 95, 8–13. [Google Scholar] [CrossRef]
- Belz, R.G. Allelopathy in crop/weed interactions—an update. Pest Manag. Sci. 2007, 63, 308–326. [Google Scholar] [CrossRef]
- Esayas, T.; Abraham, G.H.; Mashila, D. Weed Interference in Cotton (Gossypium hirsutum L.) in the Middle Awash, Ethiopia. Ethio. J. Appl. Sci. Technol. 2013, 4, 41–49. [Google Scholar]





| Weed Biomass | Weed Density | Weed Cover | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Crop Traits | Axum | Mekelle | Axum | Mekelle | Axum | Mekelle | ||||||
| 2015 | 2016 | 2015 | 2016 | 2015 | 2016 | 2015 | 2016 | 2015 | 2016 | 2015 | 2016 | |
| Days to emergence | 0.959 *** | −0.2065 ns | 0.3689 ns | −0.359 ns | 0.793 ** | −0.2797 ns | 0.3964 ns | 0.3198 ns | 0.8146 *** | 0.7438 ** | 0.7719 ** | 0.4026 ns |
| Days to heading | 0.2416 ns | 0.3453 ns | 0.3468 ns | −0.0485 ns | 0.3302 ns | 0.5559 * | 0.6263 * | 0.1626 ns | −0.0193 ns | 0.2005 ns | 0.6968 * | −0.2673 ns |
| Days to maturity | 0.4421 ns | 0.1602 ns | 0.1601 ns | −0.1499 ns | 0.3798 ns | −0.3127 ns | 0.1395 ns | 0.1959 ns | 0.4056 ns | 0.404 ns | 0.2148 ns | −0.2251 ns |
| Plant height | 0.2716 ns | 0.1603 ns | 0.0471 ns | −0.155 ns | 0.4075 ns | −0.5705 * | −0.1936 ns | 0.0465 ns | −0.0045 ns | −0.157 ns | −0.0735 ns | −0.3527 ns |
| Total tiller no. per plant | 0.2716 ns | 0.181 ns | −0.2087 ns | −0.3489 ns | −0.1073 ns | −0.524 * | 0.2831 ns | −0.538 * | −0.0447 ns | 0.0028 ns | 0.0263 ns | −0.5266 * |
| Crop biomass yield | −0.8784 *** | 0.421 ns | 0.0258 ns | −0.5686 * | −0.7859 ** | 0.1844 ns | 0.2452 ns | −0.4408 ns | −0.8632 *** | −0.8417 *** | −0.1496 ns | −0.5355 * |
| PAA Monocot | 0.1196 ns | 0.0196 ns | −0.5306 * | −0.0223 ns | −0.0298 ns | 0.0053 ns | −0.0533 ns | −0.0502 ns | 0.0418 ns | 0.3731 ns | 0.0751 ns | 0.464 ns |
| PAA Dicot | −0.2807 ns | −0.2001 ns | 0.155 ns | 0.3154 ns | −0.322 | 0.1101 ns | −0.0845 ns | −0.504 * | −0.2412 ns | −0.0657 ns | −0.1511 ns | 0.0797 ns |
| n | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 |
| Multiple Regression Models | Weed Biomass | Weed Cover | ||||||
|---|---|---|---|---|---|---|---|---|
| Whole Model | Axum | Mekelle | Axum | Mekelle | ||||
| 2015 | 2016 | 2015 | 2016 | 2015 | 2016 | 2015 | 2016 | |
| R2 (%) | 98.65 | 77.05 | 70.29 | 98.24 | 99.69 | 95.82 | 89.52 | 83.59 |
| p-value | <0.0001 | 0.1799 | 0.0691 | 0.0463 | 0.0108 | 0.0012 | 0.0024 | 0.0091 |
| p-values of independent variables | ||||||||
| Days to emergence | <0.0001 (179.2) | 0.0524 (3.4) | 0.0925 (55) | - | 0.018 (−3.3) | 0.0005 (0.4) | 0.0004 (4.3) | - |
| Days to heading | 0.0068 (29) | - | - | 0.0039 (11) | - | - | - | - |
| Days to maturity | 0.0061 (−13) | - | - | - | 0.0096 (0.5) | 0.0047 (−0.6) | - | 0.017 (−0.3) |
| Plant height | - | 0.0741 (−1.2) | - | 0.0033 (−6.3) | 0.0072 (−0.65) | - | 0.0107 (−0.4) | - |
| Tiller no./plant | - | 0.062 (9) | - | 0.0186 (−33.8) | 0.0973 (−2.1) | - | - | 0.0034 (4.3) |
| Crop Biomass | - | 0.0309 (0.01) | - | 0.0225 (0.02) | 0.0057 (−0.004) | - | - | 0.0114 (0.002) |
| PAA Monocot | 0.051 (15.4) | - | 0.0445 (38.2) | 0.0691 (−2.8) | 0.0319 (1.4) | 0.0016 (0.6) | 0.0831 (0.6) | - |
| PAA Dicot | - | 0.0591 (2.5) | - | 0.0047 (7.4) | 0.0493 (−0.66) | 0.01 (−0.3) | - | - |
| n | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 |
| Multiple Regression Models | Weed Density | |||
|---|---|---|---|---|
| Whole Model | Axum | Mekelle | ||
| 2015 | 2016 | 2015 | 2016 | |
| R2 (%) | 62.88 | 92.62 | 39.22 | 99.02 |
| p-value | 0.0062 | 0.0221 | 0.0527 | 0.0042 |
| p-values of independent variables | ||||
| Days to emergence | 0.0062 (50.4) | - | - | - |
| Days to heading | - | 0.0043 (−20.7) | 0.0527 (10.2) | 0.0042 (18) |
| Days to maturity | - | 0.0081 (24.1) | - | 0.0038 (−21.8) |
| Plant height | - | - | - | - |
| Tiller no./plant | - | 0.0463 (−19) | - | 0.0061 (−49.7) |
| Crop Biomass | - | 0.0448 (0.01) | - | 0.0037 (−0.06) |
| PAA Monocot | - | - | - | 0.0134 (5.9) |
| PAA Dicot | - | 0.0075 (−9.6) | - | 0.0013 (−11) |
| n | 10 | 10 | 10 | 10 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gebrehiwot, H.G.; Aune, J.B.; Eklo, O.M.; Torp, T.; Brandsæter, L.O. Allelopathic Potential of Teff Varieties and Effect on Weed Growth. Agronomy 2020, 10, 854. https://doi.org/10.3390/agronomy10060854
Gebrehiwot HG, Aune JB, Eklo OM, Torp T, Brandsæter LO. Allelopathic Potential of Teff Varieties and Effect on Weed Growth. Agronomy. 2020; 10(6):854. https://doi.org/10.3390/agronomy10060854
Chicago/Turabian StyleGebrehiwot, Haftamu Gebretsadik, Jens Bernt Aune, Ole Martin Eklo, Torfinn Torp, and Lars Olav Brandsæter. 2020. "Allelopathic Potential of Teff Varieties and Effect on Weed Growth" Agronomy 10, no. 6: 854. https://doi.org/10.3390/agronomy10060854
APA StyleGebrehiwot, H. G., Aune, J. B., Eklo, O. M., Torp, T., & Brandsæter, L. O. (2020). Allelopathic Potential of Teff Varieties and Effect on Weed Growth. Agronomy, 10(6), 854. https://doi.org/10.3390/agronomy10060854