Interactive Effect of Nitrogen and Azoxystrobin on Yield, Quality, Nitrogen and Water Use Efficiency of Wild Rocket in Southern Italy

 ,
, 
 ,
,  ,
, 
Abstract
1. Introduction
2. Material and Methods
2.1. Experimental Site Characteristics
2.2. Treatments, Experimental Design, and Crop Management
2.3. Biomass, Yield, and Plant Biometric Characteristics
2.4. N Uptake, Nitrogen and Water Use Efficiency
2.5. Qualitative Features
2.6. Statistical Analysis
3. Results
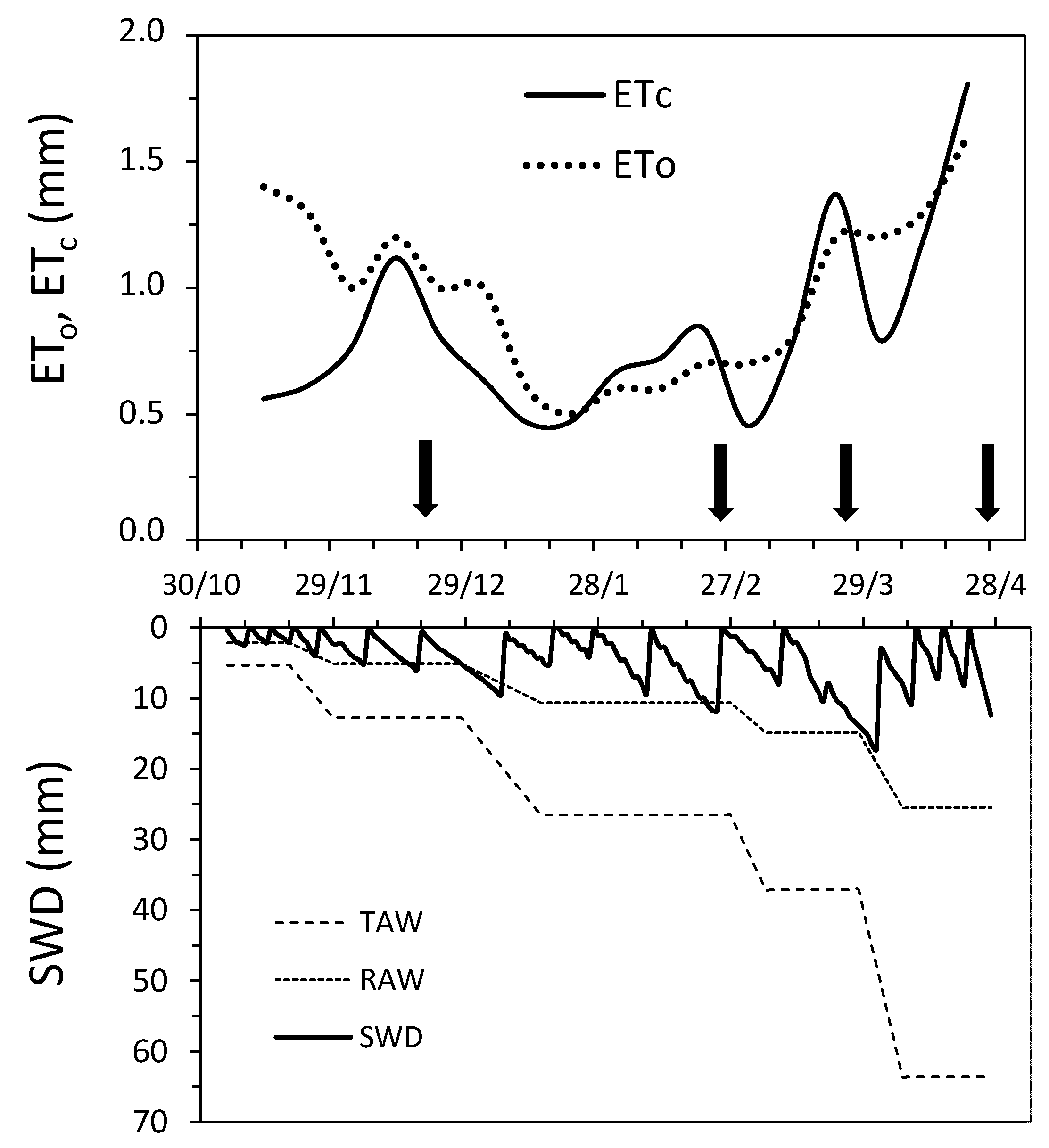
3.1. Evapotranspiration, Soil Water Regime, and Irrigation Supply
3.2. Yield and Quality Parameters
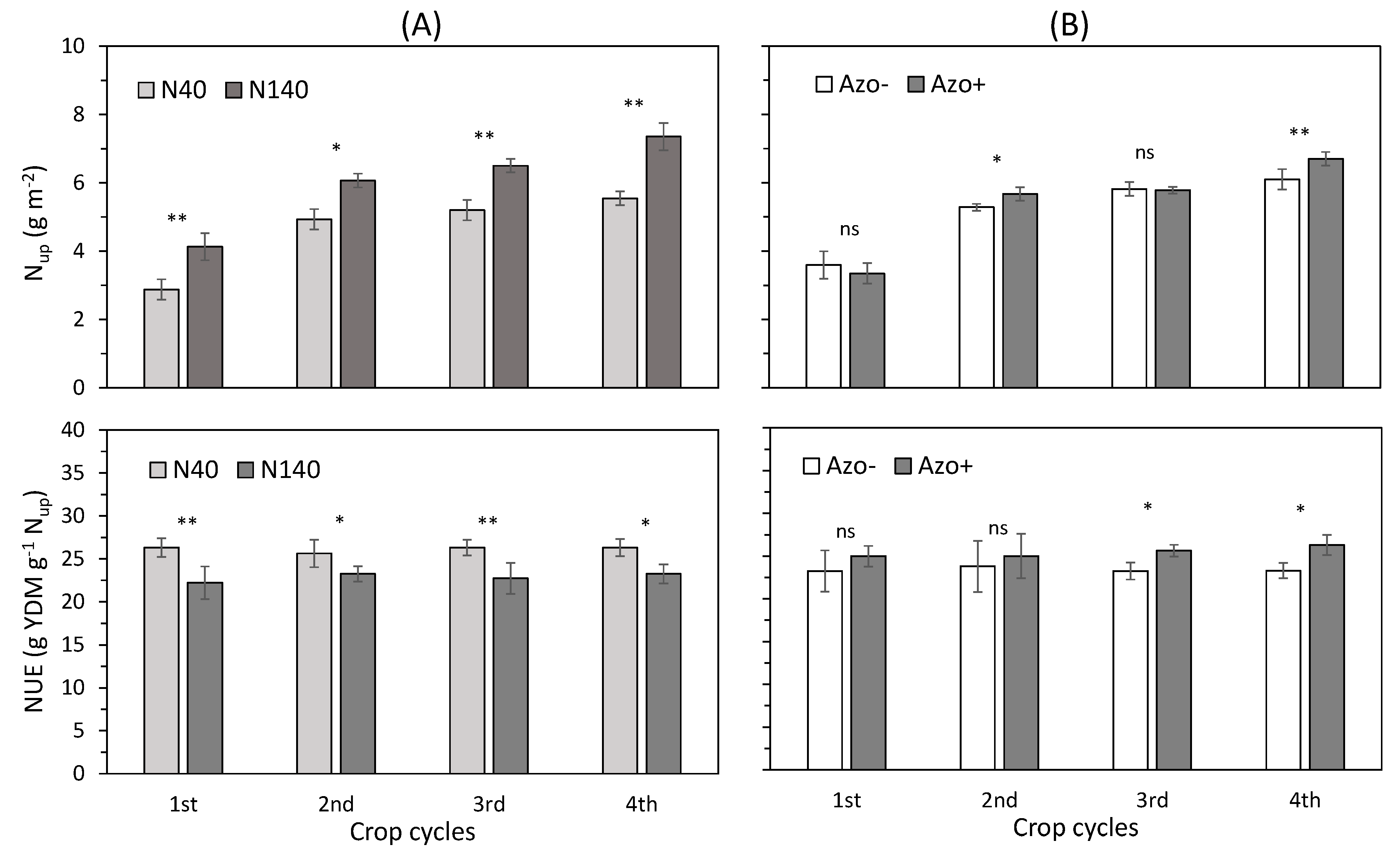
3.3. N Uptake, NUE, and WUE
4. Discussion
4.1. Yield and Qualitative Parameters
4.2. N Uptake, NUE and WUE
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nicoletti, R.; Raimo, F.; Miccio, G. Diplotaxis tenuifolia: Biology, production and properties. Eur. J. Plant Sci. Biotechnol. 2007, 1, 36–43. [Google Scholar]
- Caruso, G.; Stoleru, V.; De Pascale, S.; Cozzolino, E.; Pannico, A.; Giordano, M.; Teliban, G.; Cuciniello, A.; Rouphael, Y. Production, leaf quality and antioxidants of perennial wall rocket as affected by crop cycle and mulching type. Agronomy 2019, 9, 194. [Google Scholar] [CrossRef]
- Caruso, G.; Parrella, G.; Giorgini, M.; Nicoletti, R. Crop systems, quality and protection of Diplotaxis tenuifolia. Agriculture 2018, 8, 55. [Google Scholar] [CrossRef]
- Schiattone, M.I.; Candido, V.; Cantore, V.; Montesano, F.F.; Boari, F. Water use and crop performance of two wild rocket genotypes under salinity conditions. Agric. Water Manag. 2017, 194, 214–221. [Google Scholar] [CrossRef]
- Erice, G.; Sanz-Sáez, A.; Aranjuelo, I.; Irigoyen, J.J.; Aguirreolea, J.; Avice, J.-C.; Sánchez-Díaz, M. Photosynthesis, N2 fixation and taproot reserves during the cutting regrowth cycle of alfalfa under elevated CO2 and temperature. J. Plant Physiol. 2011, 168, 2007–2014. [Google Scholar] [CrossRef]
- Schiattone, M.I. Interventi Agronomici per Migliorare la Produzione, la Qualità e L’efficienza d’uso Dell’acqua e dell’azoto Della Ruchetta [Diplotaxis tenuiflia (L.) DC.]. [Agronomic Interventions to Improve Yield, Quality and Water and Nitrogen use Efficiency of Rocket (Diplotaxis tenuiflia L. DC.)]. Ph.D. Tesis, Università degli Studi della Basilicata, Potenza, Italy, 2018; 191p. [Google Scholar]
- Cavaiuolo, M.; Ferrante, A. Nitrates and glucosinolates as strong determinants of the nutritional quality in rocket leafy salads. Nutrients 2014, 6, 1519–1538. [Google Scholar] [CrossRef]
- D’Antuono, L.F.; Elementi, S.; Neri, R. Exploring new potential health promoting vegetables: Glucosinolates and sensory attributes of rocket salads and related Diplotaxis and Eruca species. J. Sci. Food Agric. 2009, 89, 713–722. [Google Scholar] [CrossRef]
- Di Venere, D.; Calabrese, N.; Linsalata, V.; Cardinali, A.; Bianco, V.V. Influence of sowing time on phenolic composition of rocket. Acta Hortic. 2000, 533, 343–350. [Google Scholar] [CrossRef]
- Ramos-Bueno, R.P.; Rincón-Cervera, M.A.; González-Fernández, M.J.; Guil-Guerrero, J.L. Phytochemical composition and antitumor activities of new salad greens: Rucola (Diplotaxis tenuifolia) and corn salad (Valerianella locusta). Plant Foods Hum. Nutr. 2016, 71, 197–203. [Google Scholar] [CrossRef]
- Koukounaras, A.; Bantis, F.; Karatolos, N.; Melissas, C.; Vezyroglou, A. Influence of pre-harvest factors on postharvest quality of fresh-cut and baby leafy vegetables. Agronomy 2020, 10, 172. [Google Scholar] [CrossRef]
- Colelli, G.; Elia, A. I prodotti ortofrutticoli di IV gamma: Aspetti fisiologici e tecnologici. [Physiological and technological aspects of fresh-cut horticultural products]. Italus Hortus 2009, 16, 55–78. [Google Scholar]
- Kader, A.A. Flavour quality of fruits and vegetables. J. Sci. Food Agric. 2008, 88, 1863–1868. [Google Scholar] [CrossRef]
- Mahlangu, R.I.S.; Maboko, M.M.; Mudau, F.N. Growth, yield and mineral content of basil and cultivated rocket due to plant density and nitrogen level. Int. J. Veget. Sci. 2019, 25, 1–15. [Google Scholar] [CrossRef]
- Habermeyer, M.; Roth, A.; Guth, S.; Diel, P.; Engel, K.-H.; Epe, B.; Fürst, P.; Heinz, V.; Humpf, H.-U.; Joost, H.-G.; et al. Nitrate and nitrite in the diet: How to assess their benefit and risk for human health. Mol. Nutr. Food Res. 2015, 59, 106–128. [Google Scholar] [CrossRef]
- Franzoni, G.; Cocetta, G.; Trivellini, A.; Ferrante, A. Transcriptional regulation in rocket leaves as affected by salinity. Plants 2020, 9, 20. [Google Scholar] [CrossRef]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Amaro, A.C.E.; Baron, D.; Ono, E.O.; Rodrigues, J.D. Physiological effects of strobilurin and carboxamides on plants: An overview. Acta Physiol. Plant. 2020, 42, 4. [Google Scholar] [CrossRef]
- Boari, F.; Cantore, V.; Di Venere, D.; Sergio, L.; Candido, V.; Schiattone, M.I. Pyraclostrobin can mitigate salinity stress in tomato crop. Agric. Water Manag. 2019, 222, 254–264. [Google Scholar] [CrossRef]
- Venancio, W.S.; Tavares Rodrigues, M.A.; Begliomini, E.; de Souza, N.L. Physiological effects of strobilurin fungicides on plants. UEPG Ciência Exatas E Terra Ciênc. Agrar. E Eng. Ponta Grossa.
- Bartlett, D.W.; Clough, J.M.; Godwin, J.R.; Hall, A.A.; Hamer, M.; Parr-Dobrzanski, B. Review: The strobilurin fungicides. Pest Manag. Sci. 2002, 58, 649–662. [Google Scholar] [CrossRef]
- Balba, H. Review of strobilurin fungicide chemicals. J. Environ. Sci. Health Part B 2007, 42, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Xu, X.; Lu, Z. Effect of Azoxystrobin fungicide on the physiological and biochemical indices and ginsenoside contents of ginseng leaves. J. Ginseng Res. 2018, 42, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Lopes, A.M.; Schumacher, P.V.; Martínez, A.T.P.; da Costa Netto, A.P.; Chalfun-Junior, A. Insights into the positive effect of Pyraclostrobin on sugarcane productivity. Agronomy 2018, 8, 122. [Google Scholar] [CrossRef]
- Boari, F.; Todorovic, M.; Albrizio, R.; Sellami, M.H.; Schiattone, M.I.; Cantore, V. Le strobilurine su pomodoro migliorano le rese e l’uso dell’acqua. [Strobilurins on tomato improve yield and water use]. L’Inftore Agrar. 2017, 73, 42–45. [Google Scholar]
- Cantore, V.; Lechkar, O.; Karabulut, E.; Sellami, M.H.; Albrizio, R.; Boari, F.; Stellacci, A.M.; Todorovic, M. Combined effect of deficit irrigation and strobilurin application on yield, fruit quality and water use efficiency of ‘cherry’ tomato (Solanum lycopersicum L.). Agric. Water Manag. 2016, 167, 53–61. [Google Scholar] [CrossRef]
- Giuliani, M.M.; Carucci, F.; Nardella, E.; Francavilla, M.; Ricciardi, L.; Lotti, C.; Gatta, G. Combined effects of deficit irrigation and strobilurin application on gas exchange, yield and water use efficiency in tomato (Solanum lycopersicum L.). Sci. Hortic. 2018, 233, 149–158. [Google Scholar] [CrossRef]
- van Dingenen, J.; Antoniou, C.; Filippou, P.; Pollier, J.; Gonzalez, N.; Dhondt, S.; Goossens, A.; Fotopoulos, V.; Inzé, D. Strobilurins as growth-promoting compounds: How Stroby regulates Arabidopsis leaf growth. Plant Cell Environ. 2017, 40, 1748–1760. [Google Scholar] [CrossRef]
- Amaro, A.C.E.; Ramos, A.R.P.; Macedo, A.C.; Ono, E.O.; Rodrigues, J.D. Effects of the fungicides Azoxystrobin, Pyraclostrobin and Boscalid on the physiology of Japanese cucumber. Sci. Hortic. 2018, 228, 66–75. [Google Scholar] [CrossRef]
- Joshi, J.; Sharma, S.; Guruprasad, K.N. Foliar application of Pyraclostrobin fungicide enhances the growth, rhizobial-nodule formation and nitrogenase activity in soybean (var. JS-335). Pestic. Biochem. Physiol. 2014, 114, 61–66. [Google Scholar] [CrossRef]
- Bonasia, A.; Conversa, G.; Lazzizzera, C.; Elia, A. Pre-harvest nitrogen and Azoxystrobin application enhances postharvest shelf-life in Butterhead lettuce. Postharvest Biol. Technol. 2013, 85, 67–76. [Google Scholar] [CrossRef]
- Conversa, G.; Bonaria, A.; Lazzizera, C.; Elia, A. Pre-harvest nitrogen and Azoxystrobin application enhances raw product quality and post-harvest shelf-life of baby spinach (Spinacia oleracea L.). J. Sci. Food Agric. 2014, 94, 3263–6272. [Google Scholar]
- Cassi, F.; Viviano, L. I Suoli della Basilicata-Carta pedologica della Regione Basilicata in scala 1:250.000. [The soils of Basilicata-Soil map of the Basilicata region in scale 1: 250,000]; Reg. Basilicata-Dip. Agric. Svilup. Rurale. Dir. Gen.: Potenza, Italy, 2006; 343p. [Google Scholar]
- Cantore, V.; Iovino, F.; Pontecorvo, G. Aspetti climatici e zone fitoclimatiche della Basilicata. [Climatic aspects and phytoclimatic zones of Basilicata]; CNR-IEIF: Cosenza, Italy, 1987; Volume 2, 49p. [Google Scholar]
- Regione Basilicata. I disciplinari di produzione integrata della Regione Basilicata 2014. [The integrated production disciplinary of the Basilicata Region 2014]; Regione Basilicata-CREA: Basilicata, Italy, 2014; 775p. [Google Scholar]
- Todorovic, M. An Excel-based tool for real time irrigation management at field scale. In Proceedings of the International Symposium on ‘Water and Land Management for Sustainable Irrigated Agriculture’, Cukurova Univ., Adana, Turkey, 4–8 April 2006. [Google Scholar]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration; FAO Irrig. and Drain. Paper 56; FAO: Rome, Italy, 1998; 300p. [Google Scholar]
- Gavilàn, P.; Castillo-Llanque, F. Estimating reference evapotranspiration with atmometers in a semiarid environment. Agric. Water Manag. 2009, 96, 465–472. [Google Scholar] [CrossRef]
- Cantore, V.; Boari, F.; Rubino, P. Consumi idrici ed efficienza d’uso dell’acqua della ruchetta selvatica (Diplotaxis tenuifolia L. DC.). [Water consumption and water use efficiency of wild rocket (Diplotaxis tenuifolia L. DC.)]. In Proceedings of the VI Giornate Scientifiche SOI, Spoleto, Italy, 22–25 April 2002; pp. 529–530. [Google Scholar]
- Qiu, Y.; Fu, B.; Wang, J.; Chen, L. Soil moisture variation in relation to topography and land use in a hillslope catchment of the Loess Plateau, China. J. Hydrol. 2001, 240, 243–263. [Google Scholar] [CrossRef]
- Dobermann, A. Nutrient use efficiency–measurement and management. IFA International Workshop on Fertilizer Best Management Practices, Brussels, Belgium, 7–9 March 2007; pp. 1–28. [Google Scholar]
- Santamaria, P.; Elia, A.; Serio, F. Effect of solution nitrogen concentration on yield, leaf element content, and water and nitrogen use efficiency of three hydroponically-grown rocket salad genotypes. J. Plant Nutr. 2002, 25, 245–258. [Google Scholar] [CrossRef]
- Howell, T.A.; Cuenca, R.H.; Solomon, K.H. Crop yield response. In Management of Farm Irrigation Systems; Hoffman, Ed.; ASAE: Washington, DC, USA, 1995; 312p. [Google Scholar]
- Bonasia, A.; Conversa, G.; Gonnella, M.; Serio, F.; Santamaria, P. Effects of ammonium and nitrate nutrition on yield and quality in endive. J. Hortic. Sci. Biotech. 2008, 83, 64–70. [Google Scholar] [CrossRef]
- Gatto, M.A.; Sergio, L.; Ippolito, A.; Di Venere, D. Phenolic extracts from wild edible plants to control postharvest diseases of sweet cherry fruit. Postharv. Biol. Technol. 2016, 120, 180–187. [Google Scholar] [CrossRef]
- Sergio, L.; Gonnella, M.; Renna, M.; Linsalata, V.; Gatto, M.A.; Boari, F.; Di Venere, D. Biochemical traits of asparagus cultivars and quality changes in two differently colored genotypes during cold storage. LWT-Food Sci. Technol. 2019, 101, 427–434. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophyll a and chlorophyll b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Schiattone, M.I.; Viggiani, R.; Di Venere, D.; Sergio, L.; Cantore, V.; Todorovic, M.; Perniola, M.; Candido, V. Impact of irrigation regime and nitrogen rate on yield, quality and water use efficiency of wild rocket under greenhouse conditions. Sci. Hortic. 2018, 229, 182–192. [Google Scholar] [CrossRef]
- Pimpini, F.; Giannini, M.; Lazzarin, R. Ortaggi da foglia da taglio. [Fresh-cut lafy vegetables]; Veneto Agricoltura: Piazzola sul Brenta, Italiy, 2005; 118p. [Google Scholar]
- Cantore, V.; Pace, B.; Calabrese, N.; Boari, F.; Schiattone, M.I. Effects of non-woven fabric and fertilizer on air and soil temperature, leaf gas exchange, yield and quality of wild rocket grown in organic farming. Acta Hortic. 2013, 1005, 479–486. [Google Scholar] [CrossRef]
- Hall, M.K.D.; Jobling, J.J.; Rogers, G.S. Some perspectives on rocket as a vegetable crop: A review. Veg. Crop. Res. Bull. 2012, 76, 21–41. [Google Scholar] [CrossRef]
- Bianco, V.V.; Boari, F. Up-to-date developments on wild rocket cultivation. In Workshop ‘Rocket: A Mediterranean Crop for the World’, 1996 Legnaro (Padova); IPGRI: Rome, Italy, 1997; pp. 41–49. [Google Scholar]
- Boari, F.; Cantore, V.; De Palma, E. Influenza del regime irriguo sulla ruchetta selvatica, Diplotaxis tenuifolia (L.) DC. [Influence of the irrigation regime on wild rocket, Diplotaxis tenuifolia (L.) DC.]. Proceedings of IV Giornate Scientifiche SOI, Sanremo, Italy, 1–3 April 1998; pp. 355–356. [Google Scholar]
- Sharma, L.K.; Bali, S.K. A review of methods to improve nitrogen use efficiency in agriculture. Sustainability 2018, 10, 51. [Google Scholar] [CrossRef]
- Wang, Z.; Li, S. Effects of nitrogen and phosphorus fertilization on plant growth and nitrate accumulation in vegetables. J. Plant Nutr. 2004, 27, 539–556. [Google Scholar] [CrossRef]
- Zhang, J.; Sha, Z.; Zhang, Y.; Bei, Z.; Cao, L. The effects of different water and nitrogen levels on yield, water and nitrogen utilization efficiencies of spinach (Spinacia oleracea L.). Can. J. Plant Sci. 2015, 95, 671–679. [Google Scholar] [CrossRef]
- Grossmann, K.; Kwiatkowski, J.; Casper, G. Regulation of phytohormone levels, leaf senescence and transpiration by the strobilurin Kresoxim-methyl in wheat (Triticum aestivum). J. Plant. Physiol. 1999, 154, 805–808. [Google Scholar] [CrossRef]
- Giuliani, M.M.; Gagliardi, A.; Nardella, E.; Carucci, F.; Amodio, M.L.; Gatta, G. The effect of strobilurin on ethylene production in flowers, yield and quality parameters of processing tomato grown under a moderate water stress condition in Mediterranean area. Sci. Hortic. 2019, 249, 155–161. [Google Scholar] [CrossRef]
- Abeles, G.B.; Morgan, P.W.; Saltveit, M.E. Ethylene in Plant Biology; Academic Press: San Diego, CA, USA, 1992. [Google Scholar]
- Marek, J.; de Azevedo, D.; Ono, E.O.; Rodrigues, J.D.; Faria, C.M.D.R. Photosynthetic and productive increase in tomato plants treated with strobilurins and carboxamides for the control of Alternaria solani. Sci. Hortic. 2018, 242, 76–89. [Google Scholar] [CrossRef]
- Ramos, A.R.P.; Amaro, A.C.E.; Macedo, A.C.; Souza, E.R.; Rodrigues, J.D.; Ono, E.O. Acúmulo de carboidratos no desenvolvimento de tomateiro tratado com produtos químicos. Semina. [Accumulation of carbohydrates in the development of tomato treated with chemical products]. Ciênc. Agrár. 2015, 36, 705–718. [Google Scholar] [CrossRef]
- Viana, E.M.; Kiehl, J.C. Doses de nitrogênio e potássio no crescimento do trigo. [Nitrogen and potassium doses in wheat growth]. Bragantia 2010, 69, 975–982. [Google Scholar] [CrossRef]
- Kluge, R.A.; Tezotto-Uliana, J.V.; Silva, P.P. Aspectos fisiológicos e ambientais da fotossíntese. [Physiological and environmental aspects of photosynthesis]. Rev. Virtual Química 2014, 7, 56–73. [Google Scholar]
- Kopsell, D.A.; Kopsell, D.E.; Curran-Celentano, J. Carotenoid pigments in kale are influenced by nitrogen concentration and form. J. Sci. Food Agric. 2007, 87, 900–907. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology; The Benjamin/Cummings Publishing Company, Inc.: Redwood City, CA, USA; Berlin, Germany, 2002; 675p. [Google Scholar]
- Koyama, R.; Itoh, H.; Kimura, S.; Morioka, A.; Uno, Y. Augmentation of antioxidant constituents by drought stress to roots in leafy vegetables. HortTechnology 2012, 22, 121–125. [Google Scholar] [CrossRef]
- Luoh, J.W.; Begg, C.B.; Symonds, R.C.; Ledesma, D.; Yang, R.-Y. Nutritional yield of african indigenous vegetables in water-deficient and water-sufficient conditions. Food Nutr. Sci. 2014, 5, 812–822. [Google Scholar] [CrossRef]
- EC. Regulation (EU) No 1258/2011 of 2 December 2011 amending Regulation (EC) No. 1881/2006 as regards maximum levels for nitrates in foodstuffs. Off. J. Eur. Union L 2011, 320, 15–17. [Google Scholar]
- Bonasia, A.; Lazzizera, C.; Elia, A.; Conversa, G. Nutritional, biophysical and physiological characteristics of wild rocket genotypes as affected by soilless cultivation system, salinity level of nutrient solution and growing period. Front. Plant Sci. 2017, 8, 1–15. [Google Scholar] [CrossRef]
- Signore, A.; Bell, L.; Santamaria, P.; Wagstaff, C.; Van Labeke, M.-C. Red light is effective in reducing nitrate concentration in rocket by increasing nitrate reductase activity, and contributes to increased total glucosinolates content. Front. Plant Sci. 2020, 11, 604. [Google Scholar] [CrossRef]
- Weightman, R.M.; Huckle, A.J.; Roques, S.E.; Ginsburg, D.; Dyer, C.J. Factors influencing tissue nitrate concentration in field-grown wild rocket (Diplotaxis tenuifolia) in southern England. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2012, 29, 1425–1435. [Google Scholar] [CrossRef]
- Chen, B.-M.; Wang, Z.-H.; Li, S.-X.; Wang, G.-X.; Song, H.-X.; Wang, X.-N. Effects of nitrate supply on plant growth, nitrate accumulation, metabolic nitrate concentration and nitrate reductase activity in three leafy vegetables. Plant Sci. 2004, 167, 635–643. [Google Scholar] [CrossRef]
- Devienne-Barret, F.; Justes, E.; Machet, J.M.; Mary, B. Integrated control of nitrate uptake by crop growth rate and soil nitrate availability under field conditions. Ann. Bot. 2000, 86, 995–1005. [Google Scholar] [CrossRef]
- Ceylan, S.; Mordogan, N.; Cakici, H.; Yoldas, F. Effects of different nitrogen levels on the yield and nitrogen accumulation in the rocket. Asian J. Plant Sci. 2002, 1, 482–483. [Google Scholar]
- Kristensen, H.L.; Stavridou, E. Deep root growth and nitrogen uptake by rocket (Diplotaxis tenuifolia L.) as affected by nitrogen fertilizer, plant density and leaf harvesting on a coarse sandy soil. Soil Use Manag. 2017, 33, 62–71. [Google Scholar] [CrossRef]
- Benincasa, P.; Guiducci, M.; Tei, F. The nitrogen use efficiency: Meaning and sources of variation-case studies on three vegetable crops in central Italy. HortTechnology 2011, 21, 266–273. [Google Scholar] [CrossRef]
- Rubino, P.; Cantore, V.; Mastro, M.A. Studio dell’efficienza dell’uso dell’acqua di alcune specie erbacee in un ambiente dell’Italia meridionale. [Study of the water use efficiency of some herbaceous species in an environment of Southern Italy]. Irrig. Dren. 1999, 46, 39–46. [Google Scholar]
- Ueno, O.; Bang, S.W.; Wada, Y.; Kondo, A.; Ishihara, K.; Kaneko, Y.; Matsuzawa, Y. Structural and biochemical dissection of photorespiration in hybrids differing in genome constitution between Diplotaxis tenuifolia (C3–C4) and radish (C3). Plant Physiol. 2003, 132, 1550–1559. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, A.; Sayadi, F.; Dashti, H.; Tajabadi Pour, A. Effects of water and nitrogen supply on growth, water-use efficiency and mucilage yield of isabgol (Plantago ovata Forsk). J. Soil Sci. Plant Nutr. 2013, 13, 341–354. [Google Scholar] [CrossRef]
- Giuliani, M.M.; Nardella, E.; Gatta, G.; Quintadamo, M.; De Caro, A. Processing tomato cultivated under water deficit conditions: The effect of Azoxystrobin. Acta Hortic. 2011, 914, 287–294. [Google Scholar] [CrossRef]
- Nason, M.A.; Farrar, J.; Bartlett, D. Strobilurin fungicides induce changes in photosynthetic gas exchange that do not improve water use efficiency of plants grown under conditions of water stress. Pest Manag. Sci. 2007, 63, 1191–1200. [Google Scholar] [CrossRef]
- Gilardi, G.; Demarchi, S.; Gullino, M.L.; Garibaldi, A. Management of leaf spot of wild rocket using fungicides, resistance inducers and a biocontrol agent, under greenhouse conditions. Crop Prot. 2015, 71, 39–44. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Treatments | Marketable Yield | LAI | Plant Mean Weight | Leaves | Aboveground Dry Biomass | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Number | Total Length (LT) | Bade Length (LB) | LB/LT | Width | Incision Index | |||||
| (kg m−2) | (g plant−1) | (n. plant−1) | (cm) | (cm) | (%) | (cm) | (score 1-5) | (kg m−2) | ||
| 1st crop cycle | ||||||||||
| Azoxystrobin (Azo) | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| Azo- | 1.16 | 2.85 | 2.19 | 5.08 | 16.1 | 5.9 | 36.5 | 1.89 | 3.0 | 0.15 |
| Azo+ | 1.13 | 2.72 | 2.03 | 4.97 | 16.2 | 5.8 | 35.8 | 1.88 | 2.9 | 0.16 |
| N level (NL) | * | * | ns | * | * | ns | ns | ns | ns | * |
| N40 | 1.05 | 2.48 | 2.09 | 4.80 | 15.9 | 5.8 | 36.8 | 1.90 | 3.0 | 0.14 |
| N140 | 1.24 | 3.09 | 2.14 | 5.25 | 16.4 | 5.9 | 35.5 | 1.88 | 2.9 | 0.17 |
| Azo x NL | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| 2nd crop cycle | ||||||||||
| Azoxystrobin (Azo) | * | * | * | * | * | * | ns | ns | ns | * |
| Azo- | 1.48 | 2.12 | 3.68 | 9.3 | 17.4 | 9.0 | 51.5 | 1.08 | 4.7 | 0.23 |
| Azo+ | 1.63 | 2.60 | 3.95 | 9.9 | 19.6 | 10.0 | 50.9 | 1.07 | 4.7 | 0.25 |
| N level (NL) | * | ** | * | * | ns | ns | ns | ns | * | * |
| N40 | 1.47 | 2.18 | 3.60 | 9.2 | 18.7 | 9.7 | 51.6 | 1.06 | 4.9 | 0.24 |
| N140 | 1.64 | 2.55 | 4.02 | 10.0 | 18.3 | 9.3 | 50.8 | 1.09 | 4.5 | 0.26 |
| Azo x NL | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| 3rd crop cycle | ||||||||||
| Azoxystrobin (Azo) | * | * | * | ns | ns | ns | ns | ns | ns | * |
| Azo- | 1.65 | 3.35 | 6.14 | 10.7 | 19.3 | 11.3 | 58.7 | 1.02 | 5.0 | 0.28 |
| Azo+ | 1.83 | 3.95 | 6.85 | 11.3 | 20.0 | 11.6 | 58.0 | 1.01 | 5.0 | 0.29 |
| N level (NL) | ** | * | ** | * | ns | ns | * | ns | ns | * |
| N40 | 1.61 | 3.30 | 5.79 | 10.2 | 19.8 | 11.2 | 56.7 | 1.04 | 5.0 | 0.28 |
| N140 | 1.87 | 4.01 | 7.19 | 11.8 | 19.5 | 11.7 | 60.1 | 0.98 | 5.0 | 0.29 |
| Azo x NL | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| 4th crop cycle | ||||||||||
| Azoxystrobin (Azo) | * | * | * | ns | ns | ns | ns | ns | ns | * |
| Azo- | 1.50 | 3.06 | 7.62 | 14.8 | 23.4 | 12.2 | 52.7 | 1.33 | 5.0 | 0.37 |
| Azo+ | 1.81 | 3.57 | 8.34 | 16.2 | 20.7 | 10.0 | 48.7 | 1.10 | 5.0 | 0.41 |
| N level (NL) | * | * | ** | * | ns | ns | ns | ns | ns | * |
| N40 | 1.52 | 3.03 | 7.15 | 14.2 | 20.8 | 11.0 | 53.0 | 1.24 | 5.0 | 0.37 |
| N140 | 1.80 | 3.59 | 8.80 | 16.8 | 23.2 | 11.2 | 48.5 | 1.19 | 5.0 | 0.41 |
| Azo x NL | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| Treatments | DM | Ni | TP | TAA | Chla+b | TC |
|---|---|---|---|---|---|---|
| (g 100 g−1 FW) | (mg kg−1 FW) | (mg CAE g−1 DM) | (g Trolox 100 g−1 DM) | (μg g−1 DM) | (μg g−1 DM) | |
| 1st crop cycle | ||||||
| Azoxystrobin (Azo) | ns | ns | ns | * | ns | * |
| Azo- | 7.2 | 4838.5 | 11.7 | 0.37 | 836.9 | 155.5 |
| Azo+ | 7.4 | 4536.2 | 12.6 | 0.45 | 887.0 | 164.8 |
| N level (NL) | ns | * | * | * | * | * |
| N40 | 7.2 | 4432.9 | 13.2 | 0.48 | 787.5 | 170.1 |
| N140 | 7.4 | 4942.0 | 11.1 | 0.34 | 936.4 | 150.2 |
| Azo x NL | ns | ns | ns | ns | ns | ns |
| 2nd crop cycle | ||||||
| Azoxystrobin (Azo) | ns | ns | ns | ns | * | ** |
| Azo- | 8.5 | 4334.7 | 13.6 | 0.44 | 790.7 | 149.8 |
| Azo+ | 8.7 | 4438.1 | 13.8 | 0.47 | 966.7 | 173.1 |
| N level (NL) | ns | ns | * | * | * | ** |
| N40 | 8.6 | 4398.0 | 14.5 | 0.50 | 825.6 | 175.2 |
| N140 | 8.6 | 4374.4 | 12.9 | 0.41 | 931.8 | 147.7 |
| Azo x NL | ns | ns | ns | ns | ns | ns |
| 3rd crop cycle | ||||||
| Azoxystrobin (Azo) | ns | ns | * | * | * | * |
| Azo- | 8.2 | 3265.5 | 13.6 | 0.42 | 835.4 | 169.7 |
| Azo+ | 8.1 | 2946.8 | 14.6 | 0.52 | 948.1 | 187.9 |
| N level (NL) | * | * | ns | * | * | * |
| N40 | 8.5 | 2764.2 | 14.4 | 0.53 | 936.6 | 185.4 |
| N140 | 7.9 | 3448.3 | 13.8 | 0.41 | 846.9 | 172.2 |
| Azo x NL | ns | ns | ns | ns | ns | ns |
| 4rt crop cycle | ||||||
| Azoxystrobin (Azo) | ns | ns | ** | ** | * | * |
| Azo- | 9.4 | 2017.0 | 14.8 | 0.57 | 651.5 | 189.7 |
| Azo+ | 9.7 | 1959.5 | 17.9 | 0.67 | 774.2 | 201.4 |
| N level (NL) | ns | ns | * | ** | ** | ** |
| N40 | 9.6 | 1996.0 | 17.1 | 0.75 | 787.9 | 207.5 |
| N140 | 9.5 | 1980.6 | 15.5 | 0.49 | 637.8 | 183.6 |
| Azo x NL | ns | ns | ns | ns | ns | ns |
| Treatments | 1st Crop Cycle | 2nd Crop Cycle | 3rd Crop Cycle | 4rt Crop Cycle | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Y_WUE | B_WUE | IY_WUE | Y_WUE | B_WUE | IY_WUE | Y_WUE | B_WUE | IY_WUE | Y_WUE | B_WUE | IY_WUE | |
| Azoxystrobin (Azo) | ns | ns | ns | * | ns | * | * | ns | * | ** | * | ** |
| Azo- | 33.1 | 4.3 | 37.7 | 35.1 | 5.5 | 35.3 | 36.9 | 6.2 | 36.9 | 31.2 | 7.6 | 31.5 |
| Azo+ | 32.3 | 4.4 | 36.8 | 38.6 | 6.0 | 38.7 | 41.0 | 6.6 | 41.0 | 37.6 | 8.5 | 38.0 |
| N level (NL) | ** | * | ** | * | ns | * | ** | ns | ** | * | ns | * |
| N40 | 30.0 | 4.1 | 34.1 | 34.9 | 5.6 | 35.0 | 36.1 | 6.4 | 36.1 | 31.5 | 7.7 | 31.8 |
| N140 | 35.4 | 4.7 | 40.4 | 38.8 | 5.9 | 39.0 | 41.8 | 6.4 | 41.8 | 37.3 | 8.4 | 37.7 |
| Azo x NL | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Candido, V.; Boari, F.; Cantore, V.; Castronuovo, D.; Di Venere, D.; Perniola, M.; Sergio, L.; Viggiani, R.; Schiattone, M.I. Interactive Effect of Nitrogen and Azoxystrobin on Yield, Quality, Nitrogen and Water Use Efficiency of Wild Rocket in Southern Italy. Agronomy 2020, 10, 849. https://doi.org/10.3390/agronomy10060849
Candido V, Boari F, Cantore V, Castronuovo D, Di Venere D, Perniola M, Sergio L, Viggiani R, Schiattone MI. Interactive Effect of Nitrogen and Azoxystrobin on Yield, Quality, Nitrogen and Water Use Efficiency of Wild Rocket in Southern Italy. Agronomy. 2020; 10(6):849. https://doi.org/10.3390/agronomy10060849
Chicago/Turabian StyleCandido, Vincenzo, Francesca Boari, Vito Cantore, Donato Castronuovo, Donato Di Venere, Michele Perniola, Lucrezia Sergio, Roberto Viggiani, and Maria Immacolata Schiattone. 2020. "Interactive Effect of Nitrogen and Azoxystrobin on Yield, Quality, Nitrogen and Water Use Efficiency of Wild Rocket in Southern Italy" Agronomy 10, no. 6: 849. https://doi.org/10.3390/agronomy10060849
APA StyleCandido, V., Boari, F., Cantore, V., Castronuovo, D., Di Venere, D., Perniola, M., Sergio, L., Viggiani, R., & Schiattone, M. I. (2020). Interactive Effect of Nitrogen and Azoxystrobin on Yield, Quality, Nitrogen and Water Use Efficiency of Wild Rocket in Southern Italy. Agronomy, 10(6), 849. https://doi.org/10.3390/agronomy10060849