Cultivar Dependent Impact on Yield and Its Components of Young Almond Trees under Sustained-Deficit Irrigation in Semi-Arid Environments

 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Irrigation Treatments
2.3. Plant Measurements
2.4. Experimental Design and Stadistical Analysis
3. Results and Discussion
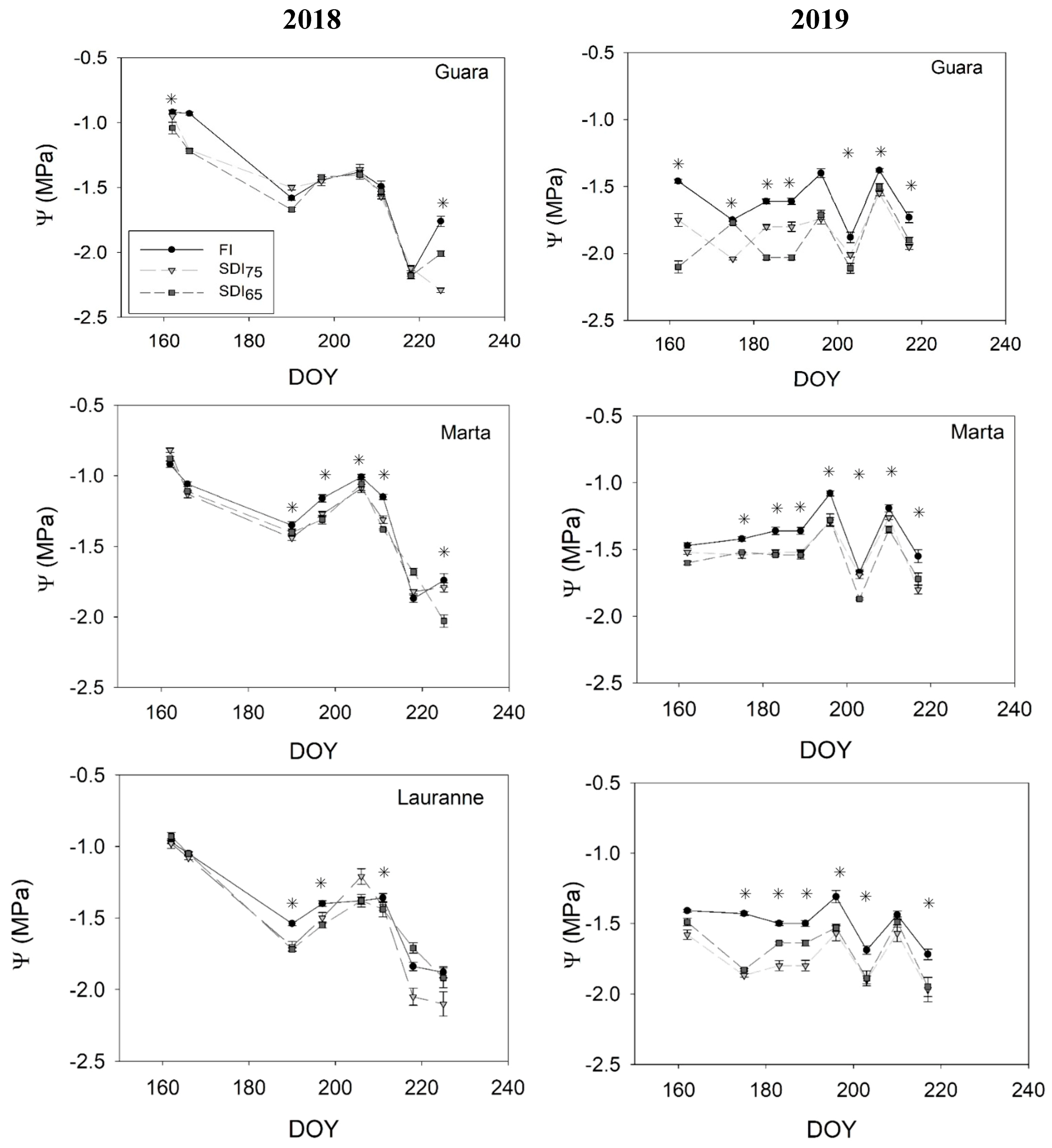
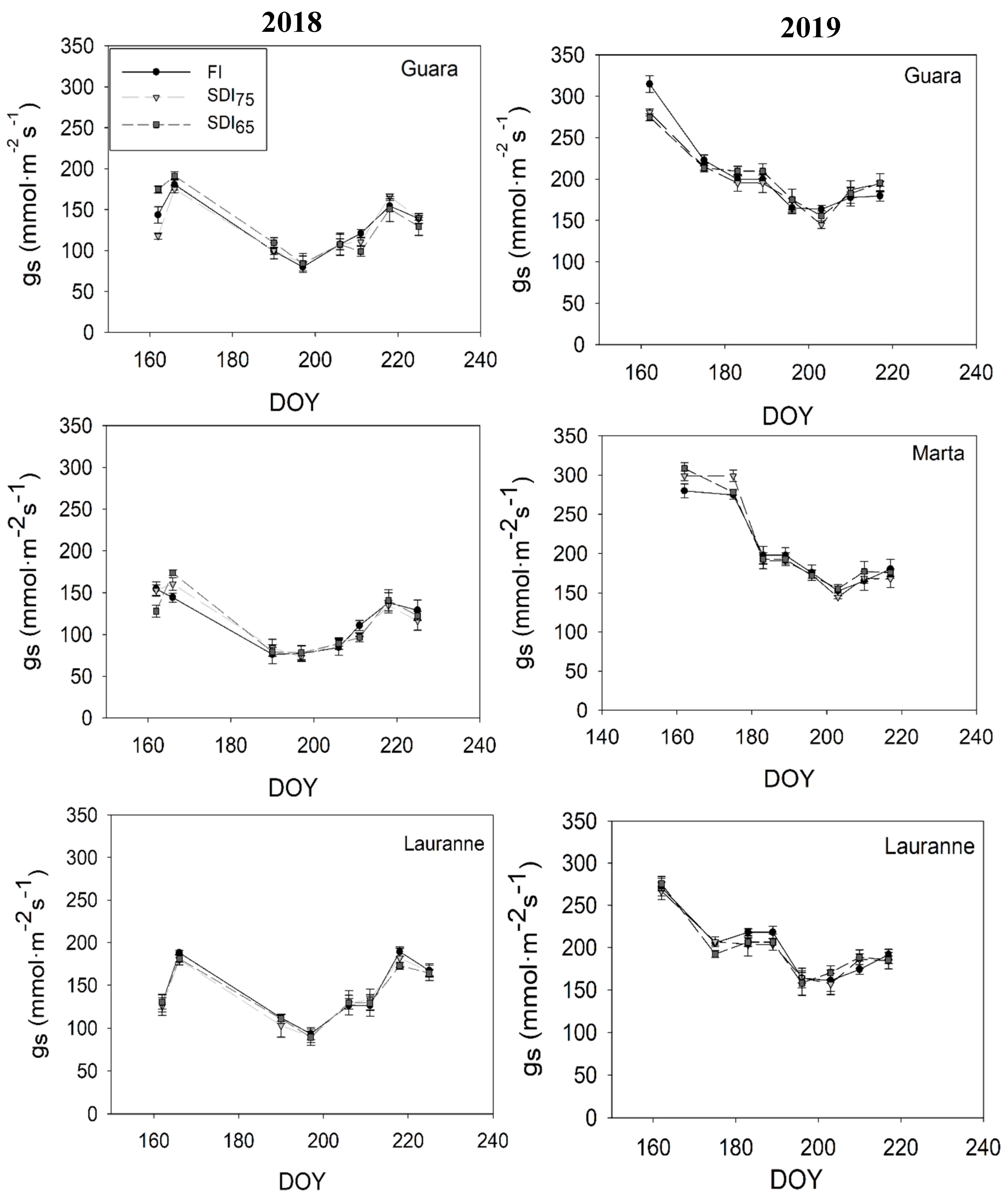
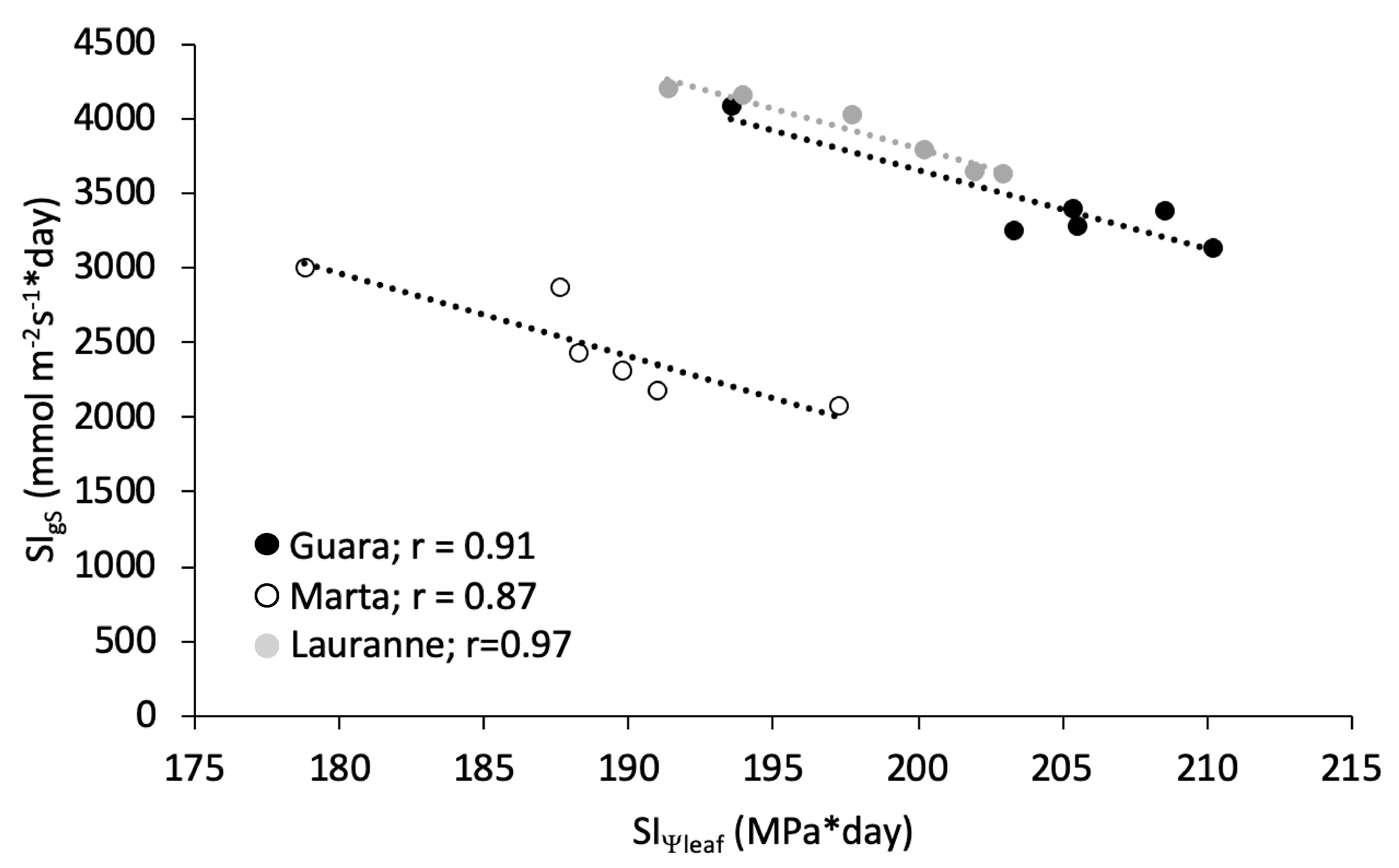
3.1. Physiological Response to Water Stress
3.2. Nut Yield and Irrigation-Water Productivity (IWP)
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fereres, E.; Soriano, M.A. Deficit irrigation for reducing agricultural water use. J. Exp. Bot. 2007, 58, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Sepaskhah, A.R.; Ahmadi, S.H. A review on partial root-zone drying irrigation. Int. J. Plant Prod. 2012, 4, 241–258. [Google Scholar]
- Expósito, A.; Berbel, J. Sustainability implications of deficit irrigation in a mature water economy: A case study in Southern Spain. Sustainability 2017, 9, 1144. [Google Scholar] [CrossRef]
- Kaya, S.; Evren, S.; Dasci, E.; Adiguzel, M.C.; Yilmaz, H. Effects of different irrigation regimes on vegetative growth, fruit yield and quality of drip-irrigated apricot trees. Afr. J. Biotechnol. 2010, 9, 5902–5907. [Google Scholar]
- Sánchez-Rodríguez, L.; Corell, M.; Hernández, F.; Sendra, E.; Moriana, A.; Carbonell-Barrachina, Á.A. Effect of Spanish-style processing on the quality attributes of HydroSOStainable green olives. J. Sci. Food Agric. 2019, 99, 1804–1811. [Google Scholar] [CrossRef] [PubMed]
- Torrecillas, A.; Ruiz-Sanchez, M.C.; Del Amor, F.; Leon, A. Seasonal variations on water relations of Amygdalus communis L. under drip irrigated and non irrigated conditions. Plant Soil 1988, 106, 215–220. [Google Scholar] [CrossRef]
- Naor, A.; Birger, R.; Peres, M.; Gal, Y.; Elhadi, F.A.; Haklay, A.; Assouline, S.; Schwartz, A. The effect of irrigation level in the kernel dry matter accumulation period on almond yield, kernel dry weight, fruit count, and canopy size. Irrig. Sci. 2018, 36, 1–8. [Google Scholar] [CrossRef]
- García Tejero, I.F.; Moriana, A.; Rodríguez Pleguezuelo, C.R.; Durán Zuazo, V.H.; Egea, G. Chapter 12-Sustainable Deficit-Irrigation Management in Almonds (Prunus dulcis L.): Different Strategies to Assess the Crop Water Status. In Water Scarcity and Sustainable Agriculture in Semiarid Environment; García Tejero, I.F., Durán Zuazo, V.H., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 271–298. [Google Scholar]
- Stewart, W.L.; Fulton, A.E.; Krueger, W.H.; Lampinen, B.D.; Shackel, K.A. Regulated deficit irrigation reduces water use of almonds without affecting yield. Calif. Agric. 2011, 65, 90–95. [Google Scholar] [CrossRef]
- Monks, D.P.; Taylor, C.; Sommer, K.; Treeby, M.T. Deficit irrigation of almond trees did not decrease yield. Acta Hortic. 2017, 251–260. [Google Scholar] [CrossRef]
- García-Tejero, I.F.; Gutiérrez Gordillo, S.; Souza, L.; Cuadros-Tavira, S.; Durán Zuazo, V.H. Fostering sustainable water use in almond (Prunus dulcis Mill.) orchards in a semiarid Mediterranean environment. Arch. Agron. Soil Sci. 2019, 65, 164–181. [Google Scholar] [CrossRef]
- Kang, S.; Zhang, J. Controlled alternate partial root-zone irrigation: Its physiological consequences and impact on water use efficiency. J. Exp. Bot. 2004, 55, 2437–2446. [Google Scholar] [CrossRef] [PubMed]
- Egea, G.; González-Real, M.M.; Baille, A.; Nortes, P.A.; Sánchez-Bel, P.; Domingo, R. The effects of contrasted deficit irrigation strategies on the fruit growth and kernel quality of mature almond trees. Irrig. Sci. 2009, 36, 1–8. [Google Scholar] [CrossRef]
- Goldhamer, D.A.; Viveros, M.; Salinas, M. Regulated deficit irrigation in almonds: Effects of variations in applied water and stress timing on yield and yield components. Irrig. Sci. 2006, 24, 101–114. [Google Scholar] [CrossRef]
- Romero, P.; Botia, P.; Garcia, F. Effects of regulated deficit irrigation under subsurface drip irrigation conditions on vegetative development and yield of mature almond trees. Plant Soil 2004, 260, 169–181. [Google Scholar] [CrossRef]
- Girona, J.; Mata, M.; Marsal, J. Regulated deficit irrigation during the kernel-filling period and optimal irrigation rates in almond. Agric. Water Manag. 2005, 75, 152–167. [Google Scholar] [CrossRef]
- Egea, G.; Nortes, P.A.; Domingo, R.; Baille, A.; Pérez-Pastor, A.; González-Real, M.M. Almond agronomic response to long-term deficit irrigation applied since orchard establishment. Irrig. Sci. 2013, 31, 445–454. [Google Scholar] [CrossRef]
- Phogat, V.; Skewes, M.A.; Mahadevan, M.; Cox, J.W. Evaluation of soil plant system response to pulsed drip irrigation of an almond tree under sustained stress conditions. Agric. Water Manag. 2013, 118, 1–11. [Google Scholar] [CrossRef]
- Alcon, F.; Egea, G.; Nortes, P.A. Financial feasibility of implementing regulated and sustained deficit irrigation in almond orchards. Irrig. Sci. 2013, 31, 931–941. [Google Scholar] [CrossRef]
- Karimi, S.; Yadollahi, A.; Arzani, K.; Imani, A.; Aghaalikhani, M. Gas-exchange response of almond genotypes to water stress. Photosynthetica 2015, 53, 29–34. [Google Scholar] [CrossRef]
- Palasciano, M.; Logoluso, V.; Lipari, E. Differences in drought tolerance in almond cultivars grown in apulia region (Southeast Italy). Acta Hortic. 2014, 319–324. [Google Scholar] [CrossRef]
- Gomes-Laranjo, J.; Coutinho, J.P.; Galhano, V.; Cordeiro, V. Responses of five almond cultivars to irrigation: Photosynthesis and leaf water potential. Agric. Water Manag. 2006, 83, 261–265. [Google Scholar] [CrossRef]
- Oliveira, I.; Meyer, A.; Afonso, S.; Gonçalves, B. Compared leaf anatomy and water relations of commercial and traditional Prunus dulcis (Mill.) cultivars under rain-fed conditions. Sci. Hortic. 2018, 229, 226–232. [Google Scholar] [CrossRef]
- Yadollahi, A.; Arzani, K.; Ebadi, A.; Wirthensohn, M.; Karimi, S. The response of different almond genotypes to moderate and severe water stress in order to screen for drought tolerance. Sci. Hortic. 2011, 129, 403–413. [Google Scholar] [CrossRef]
- Barzegar, K.; Yadollahi, A.; Imani, A.; Ahmadi, N. Influences of severe water stress on photosynthesis, water use efficiency and proline content of almond cultivars. J. Appl. Hortic. 2012, 14, 33–39. [Google Scholar] [CrossRef]
- Gutiérrez-Gordillo, S.; Durán-Zuazo, V.H.; García-Tejero, I. Response of three almond cultivars subjected to different irrigation regimes in Guadalquivir river basin. Agric. Water Manag. 2019, 222, 72–81. [Google Scholar] [CrossRef]
- RIA. Red de Información Agroclimática de Andalucía. Instituto Andaluz de Investigación y Formación Agraria y Pesquera. Consejería de Agricultura, Ganadería, Pesca y Desarrollo Sostenible. Junta de Andalucía. Available online: https://ifapa.junta-andalucia.es/agriculturaypesca/ifapa/ria/servlet/FrontController?action=Static&url=coordenadas.jsp&c_provincia=41&c_estacion=19 (accessed on 27 March 2020).
- López-López, M.; Espadador, M.; Testi, L.; Lorite, I.J.; Orgaz, F.; Fereres, E. Water use of irrigated almond trees when subjected to water deficits. Agric. Water Manag. 2018, 195, 84–93. [Google Scholar] [CrossRef]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M.; W, B. Crop evapotranspiration-Guidelines for computing crop water requirements-FAO Irrigation and drainage paper 56. Irrig. Drain. 1998, 1–15. [Google Scholar]
- García-Tejero, I.F.; Hernandez, A.; Rodriguez, V.M.; Ponce, J.R.; Ramos, V.; Muriel, J.L.; Zuazo, V.H.D. Estimating Almond Crop Coefficients and Physiological Response to Water Stress in Semiarid Environments (SW Spain). J. Agric. Sci. Technol. 2015, 17, 1255–1266. [Google Scholar]
- Myers, B.J. Water stress integral—a link between short-term stress and long-term growth. Tree Physiol. 1988, 4, 315–323. [Google Scholar] [CrossRef]
- García-Tejero, I.F.; Gutiérrez-Gordillo, S.; Ortega-Arévalo, C.; Iglesias-Contreras, M.; Moreno, J.M.; Souza-Ferreira, L.; Durán-Zuazo, V.H. Thermal imaging to monitor the crop-water status in almonds by using the non-water stress baselines. Sci. Hortic. 2018, 238, 91–97. [Google Scholar] [CrossRef]
- Hernandez-Santana, V.; Fernández, J.E.; Rodriguez-Dominguez, C.M.; Romero, R.; Diaz-Espejo, A. The dynamics of radial sap flux density reflects changes in stomatal conductance in response to soil and air water deficit. Agric. For. Meteorol. 2016, 218–219, 92–101. [Google Scholar] [CrossRef]
- Fernández, J.E. Understanding olive adaptation to abiotic stresses as a tool to increase crop performance. Environ. Exp. Bot. 2014, 103, 158–179. [Google Scholar] [CrossRef]
- Fu, X.; Meinzer, F.C. Metrics and proxies for stringency of regulation of plant water status (iso/anisohydry): A global data set reveals coordination and trade-offs among water transport traits. Tree Physiol. 2018, 39, 122–134. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Santana, V.; Rodriguez-Dominguez, C.M.; Fernández, J.E.; Diaz-Espejo, A. Role of leaf hydraulic conductance in the regulation of stomatal conductance in almond and olive in response to water stress. Tree Physiol. 2016, 36, 725–735. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Dominguez, C.M.; Buckley, T.N.; Egea, G.; de Cires, A.; Hernandez-Santana, V.; Martorell, S.; Diaz-Espejo, A. Most stomatal closure in woody species under moderate drought can be explained by stomatal responses to leaf turgor. Plant Cell Environ. 2016, 39, 2014–2026. [Google Scholar] [CrossRef]
- Espadafor, M.; Orgaz, F.; Testi, L.; Lorite, I.J.; González-Dugo, V.; Fereres, E. Responses of transpiration and transpiration efficiency of almond trees to moderate water deficits. Sci. Hortic. (Amst.) 2017, 225, 6–14. [Google Scholar] [CrossRef]
- Egea, G.; Nortes, P.A.; González-Real, M.M.; Baille, A.; Domingo, R. Agronomic response and water productivity of almond trees under contrasted deficit irrigation regimes. Agric. Water Manag. 2010, 97, 171–181. [Google Scholar] [CrossRef]
- Alegre Castellví, S.; Miarnau i Prim, X.; Romero Romero, M.; Vargas García, F. Potencial productivo de seis variedades de almendro. Frutic. Prof. 2007, 169, 23–29. [Google Scholar]
- Miarnau, X.; Torguet, L.; Batlle, I.; Romero, A.; Rovira, M.; Alegre, S. Comportamiento agronómico y productivo de las nuevas variedades de almendro. Fruticultura 2016, 49, 42–59. [Google Scholar]
- Miarnau, X.; Torguet, L.; Batlle, I.; Romero, A.; Rovira, M.; Alegre, S. Resultados agronómicos y productivos de los ensayos IRTA de nuevas variedades de almendro. Irta Jorn. Almendro Ix Ed. 2017, 43–63. [Google Scholar]
- Miarnau, X.; Torguet, L.; Batlle, I.; Romero, A.; Rovira, M.; Alegre, S. La revolución del almendro: Nuevas variedades y nuevos modelos productivos. Simp. Nac. Almendro Otros Frutos Secos 2015, 6–27. [Google Scholar]
- Lipan, L.; Cano-Lamadrid, M.; Corell, M.; Sendra, E.; Hernández, F.; Stan, L.; Vodnar, D.C.; Vázquez-Araújo, L.; Carbonell-Barrachina, Á.A. Sensory profile and acceptability of hydrosostainable almonds. Foods 2019, 8, 64. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| First Year (2018) | Second Year (2019) | |||
|---|---|---|---|---|
| SIleaf | SIgs | SIleaf | SIgs | |
| ANOVA | (MPa·day) | (mmol m−2 s−1·day) | (MPa·day) | (mmol m−2 s−1·day) |
| Irrigation | ns | ns | ns | * |
| Cultivar | * | * | * | ** |
| Irrigation × cultivar | ns | ns | * | * |
| Tukey multiple range test | ||||
| Irrigation | ||||
| FI | 198.7a | 3069a | 188.7a | 3136a |
| SDI75 | 199.4a | 3071a | 197.9a | 3426ab |
| SDI65 | 197.9a | 3152a | 199.6a | 3758b |
| Cultivar | ||||
| Marta | 189.6b | 2301b | 187.8b | 2651b |
| Lauranne | 201.6a | 3685a | 194.3ab | 4127a |
| Guara | 204.7a | 3306a | 204.1a | 3532a |
| Irrigation × cultivar | ||||
| cv. Marta | ||||
| FI | 190.9a | 2174a | 178.8d | 3005b |
| SDI75 | 189.8a | 2306a | 187.6c | 2874b |
| SDI65 | 188.2a | 2422a | 197.2ab | 2075c |
| cv. Lauranne | ||||
| FI | 201.9b | 3784b | 193.9bc | 4151a |
| SDI75 | 202.9b | 3633b | 197.7ab | 4023a |
| SDI65 | 200.2b | 3639b | 191.3c | 4207a |
| cv. Guara | ||||
| FI | 203.3b | 3249b | 193.5bc | 4089a |
| SDI75 | 205.5b | 3276b | 208.5a | 3381b |
| SDI65 | 205.4b | 3395b | 210.2a | 3127b |
| Cultivar | First Year (2018) | Second Year (2019) | ||||
|---|---|---|---|---|---|---|
| FI | SDI75 | SDI65 | FI | SDI75 | SDI65 | |
| Kernel yield (kg ha−1) | ||||||
| Guara | 1929a | 1659b | 1704b | 2254a | 2081ab | 1871b |
| Marta | 1933a | 1677b | 1775b | 2218a | 2209a | 2243a |
| Lauranne | 2349a | 2343a | 2241a | 2326a | 2105a | 2196a |
| Ratio (kernel/nut) | ||||||
| Guara | 0.41a | 0.41a | 0.41a | 0.34a | 0.39b | 0.37b |
| Marta | 0.32a | 0.31a | 0.33a | 0.34a | 0.34a | 0.34a |
| Lauranne | 0.36a | 0.36a | 0.36a | 0.33a | 0.33a | 0.32a |
| Kernel unit weight (g) | ||||||
| Guara | 1.40a | 1.40a | 1.41a | 0.99b | 1.00b | 1.22a |
| Marta | 1.31a | 1.37b | 1.33a | 1.21a | 1.18a | 1.18a |
| Lauranne | 1.12a | 1.13a | 1.13a | 1.03a | 1.05a | 1.08a |
| Fruits number tree−1 | ||||||
| Guara | 6611a | 5688b | 5801b | 10930a | 9989ab | 7363b |
| Marta | 7083a | 5875b | 6407ab | 8799a | 8984a | 9125a |
| Lauranne | 10068a | 9952a | 9519a | 10828a | 9621b | 9758b |
| Cultivar | First Year (2018) | Second Year (2019) | ||||
|---|---|---|---|---|---|---|
| (kg·m−3) | ||||||
| FI | SDI75 | SDI65 | FI | SDI75 | SDI65 | |
| Guara | 0.39c | 0.45b | 0.51a | 0.29b | 0.36a | 0.36a |
| Marta | 0.39c | 0.45b | 0.53a | 0.29b | 0.34b | 0.44a |
| Lauranne | 0.47b | 0.63a | 0.67a | 0.30b | 0.37ab | 0.43a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gutiérrez-Gordillo, S.; Durán Zuazo, V.H.; Hernández-Santana, V.; Ferrera Gil, F.; García Escalera, A.; Amores-Agüera, J.J.; García-Tejero, I.F. Cultivar Dependent Impact on Yield and Its Components of Young Almond Trees under Sustained-Deficit Irrigation in Semi-Arid Environments. Agronomy 2020, 10, 733. https://doi.org/10.3390/agronomy10050733
Gutiérrez-Gordillo S, Durán Zuazo VH, Hernández-Santana V, Ferrera Gil F, García Escalera A, Amores-Agüera JJ, García-Tejero IF. Cultivar Dependent Impact on Yield and Its Components of Young Almond Trees under Sustained-Deficit Irrigation in Semi-Arid Environments. Agronomy. 2020; 10(5):733. https://doi.org/10.3390/agronomy10050733
Chicago/Turabian StyleGutiérrez-Gordillo, Saray, Víctor Hugo Durán Zuazo, Virginia Hernández-Santana, Fernando Ferrera Gil, Amelia García Escalera, José Juan Amores-Agüera, and Iván Francisco García-Tejero. 2020. "Cultivar Dependent Impact on Yield and Its Components of Young Almond Trees under Sustained-Deficit Irrigation in Semi-Arid Environments" Agronomy 10, no. 5: 733. https://doi.org/10.3390/agronomy10050733
APA StyleGutiérrez-Gordillo, S., Durán Zuazo, V. H., Hernández-Santana, V., Ferrera Gil, F., García Escalera, A., Amores-Agüera, J. J., & García-Tejero, I. F. (2020). Cultivar Dependent Impact on Yield and Its Components of Young Almond Trees under Sustained-Deficit Irrigation in Semi-Arid Environments. Agronomy, 10(5), 733. https://doi.org/10.3390/agronomy10050733