The Effects of Hydrogen Peroxide on Plant Growth, Mineral Accumulation, as Well as Biological and Chemical Properties of Ficus deltoidea


Abstract
1. Introduction
2. Materials and Methodology
2.1. Plant Material and Experimental Site
2.2. Plant Physiology and Growth Parameters
2.3. Mineral Accumulation
2.4. Biologic and Chemical Properties
2.4.1. Sample Preparation Procedure
2.4.2. DPPH• Radical -Scavenging Activity, in Vitro α-Glucosidase Inhibitory, Total Phenolic and Total Flavonoid Content Assay
2.4.3. High Performance Thin Layer Chromatography Fingerprint (HPTLC)
2.5. Statistical Analysis
3. Results
3.1. Effects of Hydrogen Peroxide (H2O2) on Morphologic and Physiological Characteristics
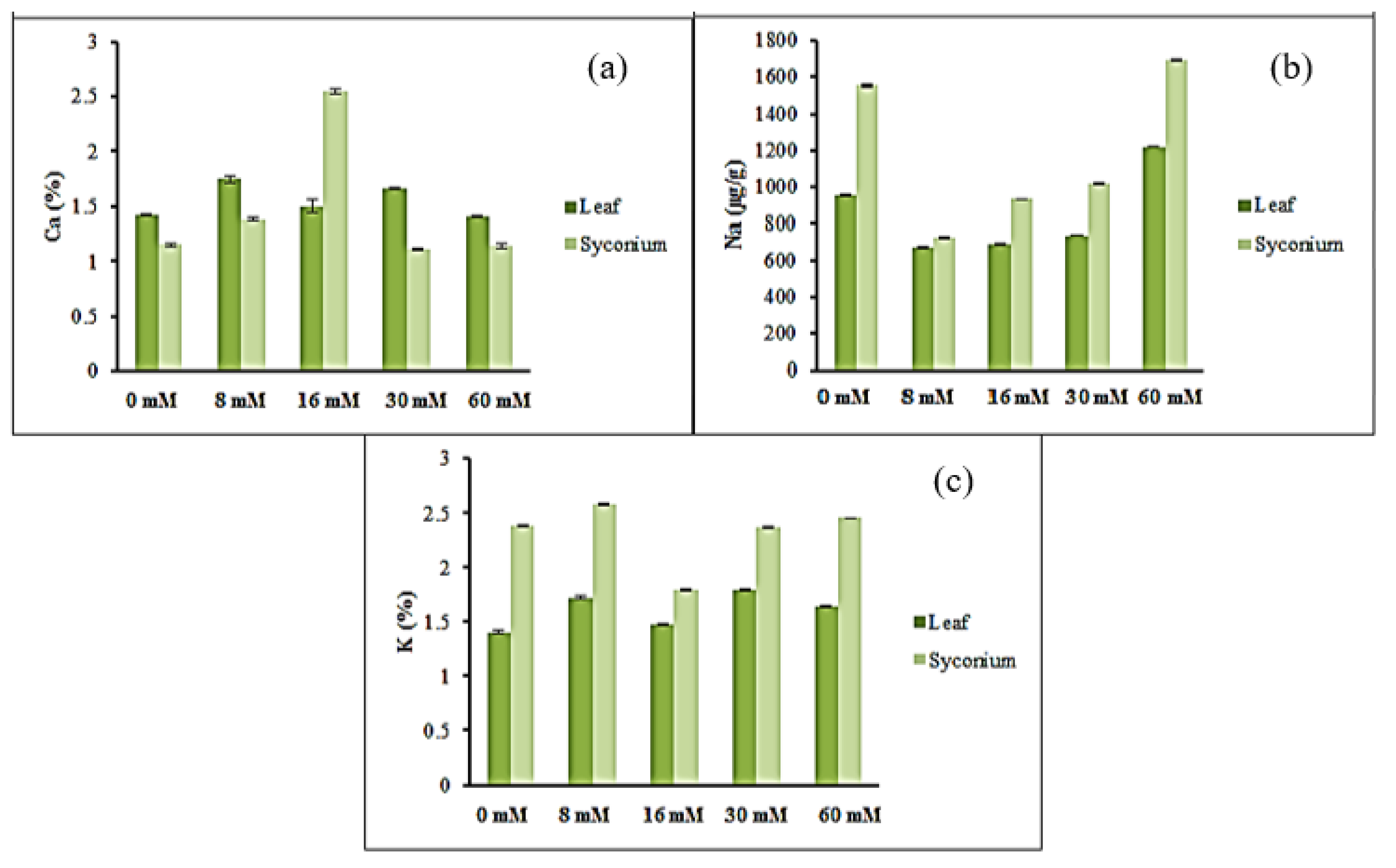
3.2. Mineral Accumulation
3.3. Effect of Hydrogen Peroxide (H2O2) on Biologic and Chemical Properties
3.3.1. DPPH• Scavenging Activity
3.3.2. In Vitro α-Glucosidase Assay
3.3.3. Total Phenolic Content
3.3.4. Total Flavonoid Content
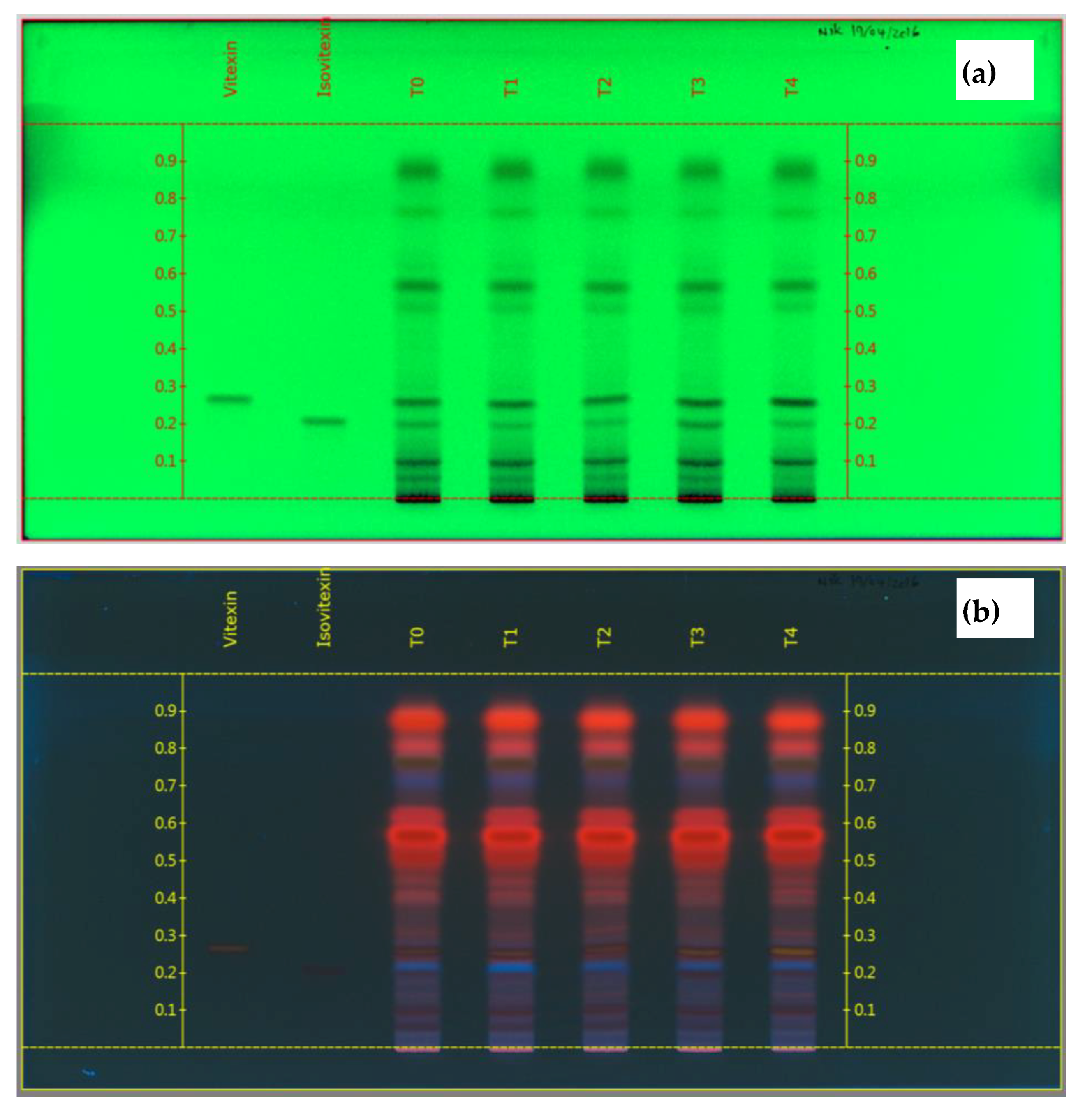
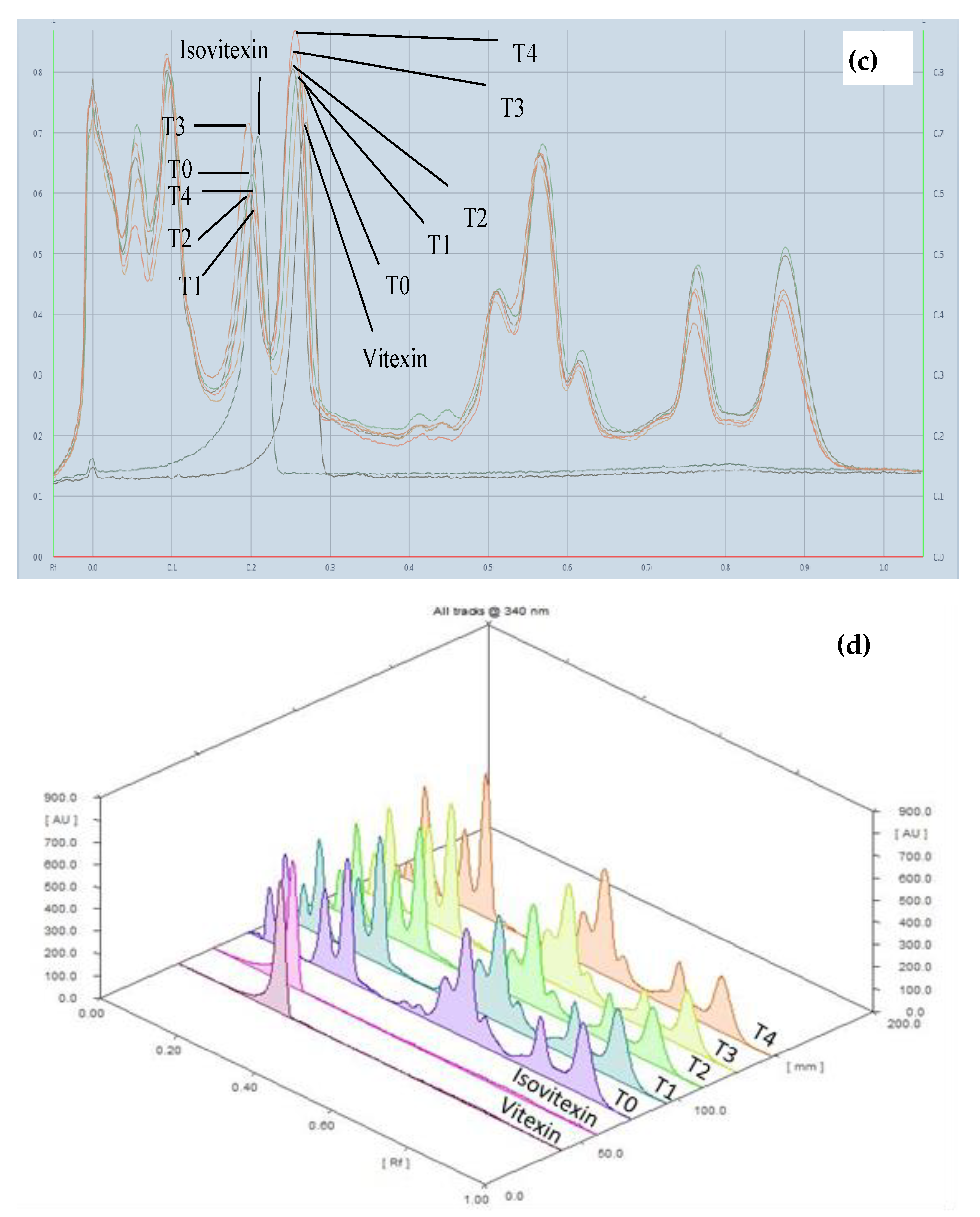
3.3.5. High Performance Thin Layer Chromatography (HPTLC) Fingerprint
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hasham, R.; Choi, H.K.; Sarmidi, M.R.; Park, C.S. Protective effects of a Ficus deltoidea extract against UVB-induced photoageing in skin cells. Biotechnol. Bioprocess Eng. 2013, 18, 185–193. [Google Scholar] [CrossRef]
- Starr, F.; Starr, K.; Loope, L. Ficus deltoidea; United States Geological Survey—Biological Resources Division, Haleakala Field Station: Maui, HI, USA, 2003. [Google Scholar]
- Badron, U.H.; Talip, N.; Mohamad, A.L.; Affenddi, A.E.A.; Ahmad Juhari, A.A. Studies on leaf venation in selected taxa of the genus Ficus L. (Moraceae) in peninsular Malaysia. Trop. Life Sci. Res. 2014, 25, 111–125. [Google Scholar] [PubMed]
- Musa, Y. Variability in morphology and agronomy of mas cotek accessions found in Kelantan and Terengganu. Bul. Teknol. Tanam. 2005, 2, 35–48. [Google Scholar]
- Misbah, H.; Abdul Aziz, A.; Aminudin, N. Antidiabetic and antioxidant properties of F. deltoidea fruit extracts and fractions. BMC Complement. Altern. Med. 2013, 13, 118. [Google Scholar] [CrossRef] [PubMed]
- Draman, S.; Aris, M.A.M.; Razman, S.F.; Akter, U.; Azlina, H.; Norazlina, A.R.; Norazlansha, M. Mas cotek (Ficus deltoidea): A possible supplement for type II diabetes: (A pilot study). Pertanika J. Trop. Agric. Sci. 2012, 35, 93–102. [Google Scholar]
- Sulaiman, M.R.; Hussain, M.K.; Zakaria, Z.A. Evaluation of the antinociceptive activity of Ficus deltoidea aqueous extract. Fitoterapia 2008, 79, 557–561. [Google Scholar] [CrossRef]
- Haida, Z.; Syahida, A.; Ariff, S.M.; Hakiman, M. Factors Affecting Cell Biomass and Flavonoid Production of Ficus deltoidea var. kunstleri in Cell Suspension Culture System. Sci. Rep. 2009, 9, 9533. [Google Scholar] [CrossRef]
- Ashraf, K.; Halim, H.; Lim, S.M.; Ramasamy, K.; Sultan, S. In vitro antioxidant, antimicrobial and antiproliferative studies of four different extracts of Orthosiphon stamineus, Gynura procumbens and Ficus deltoidea. Saudi J. Biol. Sci. 2020, 27, 417–432. [Google Scholar] [CrossRef]
- Abolmaesoomi, M.; Abdul Aziz, A.; Mat Junit, S.; Mohd Ali, J. Ficus deltoidea: Effects of solvent polarity on antioxidant and anti-proliferative activities in breast and colon cancer cells. Eur. J. Integr. Med. 2019, 28, 57–67. [Google Scholar] [CrossRef]
- Nazarni Che Isa, M.; Ajit, A.; Naila, A.; Sulaiman, A.Z. Effect of microwave assisted hydro-distillation extraction on extracts of Ficus deltoidea. Mater. Today Proc. 2018, 5, 21772–21779. [Google Scholar] [CrossRef]
- Fasihuddin, B.A.; Din, L.B. Medicinal plants used by various Ethnic Groups in Sabah. In Proceedings of the The French Malaysian-Symposium on Natural Products, Kuala Lumpur, Malaysia, 24–25 July 2002; 85p. [Google Scholar]
- Armaghan, S.; Nahdzatul, S.M.; Zeyad, D.N.; Abdalrahim, F.A.A.; Amin, M.S.; Zhari, I. Antiangiogenic effect of Ficus deltoidea Jack standardised leaf extracts. Trop. J. Pharm. Res. 2014, 13, 761–768. [Google Scholar]
- Aris, S.R.S.; Mustafa, S.; Ahmat, N.; Jaafar, F.M.; Ahmad, R. Phenolic content and antioxidant activity of fruits of Ficus deltoidea var Angustifolia sp. Malay. J. Anal. Sci. 2009, 13, 146–150. [Google Scholar]
- Nurdiana, S.; Goh, Y.M.; Hafandi, A.; Dom, S.M.; Nur Syimal’ain, A.; Noor Syaffinaz, N.M.; Ebrahimi, M. Improvement of spatial learning and memory, cortical gyrification patterns and brain oxidative stress markers in diabetic rats treated with Ficus deltoidea leaf extract and vitexin. J. Trad. Complement. Med. 2018, 8, 190–202. [Google Scholar] [CrossRef]
- Oh, M.J.; Abdul, H.M.; Ngadiran, S.; Seo, Y.K.; Sarmidi, M.R.; Park, C.S. Ficus deltoidea extract exerted anti-melanogenic activity by preventing tyrosinase activity in vitro and by suppressing tyrosinase gene expression in B16F1 melanoma cells. Arch. Dermat. Res. 2011, 303, 161–170. [Google Scholar] [CrossRef]
- Cakmak, I.; Marschner, H. Effect of zinc nutritional-status on activities of superoxide radical and hydrogen-peroxide scavenging enzymes in bean-leaves. Plant Soil. 1993, 155, 127–130. [Google Scholar] [CrossRef]
- Almeida, J.M.; Fidalgo, F.; Confraria, A.; Santos, A.; Pires, H.; Santos, I. Effect of hydrogen peroxide on catalase gene expression, isoform activities and levels in leaves of potato sprayed with homobrassinolide and ultrastructural changes in mesophyll cells. Funct. Plant Biol. 2005, 32, 707–720. [Google Scholar] [CrossRef]
- Orabi, S.A.; Dawood, M.G.; Salman, S.R. Comparative study between the physiological role of hydrogen peroxide and salicylic acid in alleviating the harmful effect of low temperature on tomato plants grown under sand-ponic culture. Sci. Agric. 2015, 9, 49–59. [Google Scholar]
- Khandaker, M.M.; Boyce, A.N.; Osman, N. The influence of hydrogen peroxide on the growth, development and quality of wax apple (Syzygium samarangense, [Blume] Merrill & L.M. Perry var. jambu madu) fruits. Plant Physiol. Biochem. 2012, 53, 101–110. [Google Scholar] [CrossRef]
- Xia, X.-J.; Huang, L.-F.; Zhou, Y.-H.; Mao, W.-H.; Shi, K.; Wu, J.-X.; Asami, T.; Chen, Z.; Yu, J.-Q. Brassinosteroids promote photosynthesis and growth by enhancing activation of Rubisco and expression of photosynthetic genes in Cucumis sativus. Planta 2009, 230, 1185–1196. [Google Scholar] [CrossRef]
- Nashriyah, M.; Zaini, H.; Mazleha, M.; Abdul, K.W. Mineral uptake by taro (Colocasia esculenta) in swamp agroecosystem following gramaxone® (paraquat). J. Nuclear Relat. Technol. 2006, 3, 59–68. [Google Scholar]
- Akter, M.S.; Ahmed, M.; Eun, J.B. Solvent effects on antioxidant properties of persimmon (Diospyros kaki, L. cv Daebong) seeds. Int. J. Food. Sci. Technol. 2010, 45, 2258–2264. [Google Scholar] [CrossRef]
- Choo, C.Y.; Sulong, N.Y.; Man, F.; Wong, T.W. Vitexin and isovitexin from the Leaves of Ficus deltoidea with in-vivo -glucosidase inhibition. J. Ethnopharmacol. 2012, 142, 776–781. [Google Scholar] [CrossRef]
- Singleton, V.; Rossi, J.A.J. Colorimetry of total phenolic with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Viticult. 1965, 16, 144–158. [Google Scholar]
- Woisky, R.G.; Salatino, A. Analysis of propolis: Some parameters and procedures for chemical quality control. J. Apicult. Res. 1998, 37, 99–105. [Google Scholar] [CrossRef]
- Mohd, K.S.; Azemin, A.; Hamil, M.S.R.; Bakar, A.R.A.; Dharmaraj, S.; Hamdan, M.R.; Ismail, Z. Application of high performance thin layer chromatography and fourier transform Infrared profiling coupled with chemometrics for the differentiation of the varieties of Ficus deltoidea Jack. Asian J. Pharma Clin. Res. 2014, 7, 110–116. [Google Scholar]
- Kato, M.; Shimizu, S. Chlorophyll metabolism in higher plants. VII. Chlorophyll degradation in senescing tobacco leaves: Phenolic-dependent peroxidative degradation. Can. J. Bot. 1987, 65, 729–735. [Google Scholar] [CrossRef]
- Li, S.; Xue, L.; Xu, S.; Feng, H.; An, L. Hydrogen peroxide involvement in formation and development of adventitious roots in cucumber. Plant Growth Regul. 2007, 52, 173–180. [Google Scholar] [CrossRef]
- Deng, X.P.; Cheng, Y.J.; Wu, X.B.; Kwak, S.S.; Chen, W.; Egrinya, A. Exogenous hydrogen peroxide positively influences root growth and metabolism in leaves of sweet potato seedlings. Aust. J. Crop Sci. 2012, 6, 1572–1578. [Google Scholar]
- Rodriguez, A.A.; Grunberg, K.A.; Taleisnik, E.L. Reactive oxygen species in the elongation zone of maize meves are necessary for leaf extension. Plant Physiol. 2014, 129, 1627–1632. [Google Scholar] [CrossRef] [PubMed]
- Uchida, A.; Jagendorf, A.T.; Hibino, T.; Takabe, T.; Takabe, T. Effects of hydrogen peroxide and nitric oxide on both salt and heat stress tolerance in rice. Plant Sci. 2002, 163, 515–523. [Google Scholar] [CrossRef]
- Niu, L.; Liao, W. Hydrogen peroxide signaling in plant development and abiotic responses: Crosstalk with nitric oxide and calcium. Front. Plant Sci. 2016, 7, 230. [Google Scholar] [CrossRef] [PubMed]
- Hameed, A.; Farooq, S. Influence of exogenous application of hydrogen peroxide on root and seedling growth on wheat (Triticum aestivum L.). Int. J. Agric. Biol. 2004, 6, 366–369. [Google Scholar]
- Stefanowicz, A.M.; Stanek, M.; Woch, M.W.; Kapusta, P. High concentration of heavy metals in beech forest understory plants growing in waste heaps leaft by Zn-Pb ore mining. Geochem. Explor. 2016, 59, 157–162. [Google Scholar] [CrossRef]
- Han, Y.H.; Yang, G.M.; Fu, J.W.; Guan, D.X.; Chen, Y.; Ma, L.Q. Arsenic-induced plant growth of arsenic-hyper accumulator in Pteris vittata: Impact of arsenic and phosphate rock. Chemosphere 2016, 149, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Lv, J.; Liu, K.; Zhao, X.; Cao, Y. Major controlling factors and prediction models for arsenic uptake from soil to wheat plants. J. Exotoxicol. Environ. Saf. 2016, 130, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Islam, F.; Ali, B.; Najeeb, U.; Mao, B.; Gill, R.A.; Yan, G.; Siddique, K.H.M.; Zhou, W. Arsenic toxicity in plants: Cellular and molecular mechanisms of its transport and metabolism. Environ. Exp. Bot. 2016, 132, 42–52. [Google Scholar] [CrossRef]
- Pandey, N.; Bhatt, R. Role of soil associated Exiguobacterium in reducing arsenic toxicity and promoting plant growth in Vigna radiata. Eur. J. Soil Biol. 2016, 75, 142–150. [Google Scholar] [CrossRef]
- Pierart, A.; Shahid, M.; Séjalon-Delmas, N.; Dumat, C. Antimony bioavailability: Knowledge and research perspectives for sustainable agricultures. J. Hazard. Mat. 2015, 289, 219–234. [Google Scholar] [CrossRef]
- Pérez-Sirvent, C.; Martínez-Sánchez, M.J.; Martínez-López, S.; Bech, J.; Bolan, N. Distribution and bioaccumulation of arsenic and antimony in Dittrichia viscosa growing in mining-affected semiarid soils in southeast Spain. J. Geochem. Explor. 2012, 123, 128–135. [Google Scholar] [CrossRef]
- Briat, J.F.; Dubos, C.; Gaymard, F. Iron nutrition, biomass production, and plant product quality. Trends Plant Sci. 2015, 20, 33–40. [Google Scholar] [CrossRef]
- Tatagiba, S.D.; Rodrigues, F.A. Magnesium decreases the symptoms of leaf scald on rice leaves. Trop. Plant Pathol. 2016, 41, 132–137. [Google Scholar] [CrossRef]
- Jia, Y.; Kong, X.; Weiser, M.D.; Lv, Y.; Akbar, S.; Jia, X.; Tian, X. Sodium limits litter decomposition rates in a subtropical forest: Additional tests of the sodium ecosystem respiration hypothesis. Appl. Soil Ecol. 2015, 93, 98–104. [Google Scholar] [CrossRef]
- Zhang, X.L.; Jia, X.F.; Yu, B.; Gao, Y.; Bai, J.G. Exogenous hydrogen peroxide influences antioxidant enzyme activity and lipid peroxidation in cucumber leaves at low light. Sci. Hortic. 2011, 129, 656–662. [Google Scholar] [CrossRef]
- Hu, W.H.; Song, X.S.; Shi, K.; Xia, X.J.; Zhou, Y.H.; Yu, J.Q. Changes in electron transport, superoxide dismutase and ascorbate peroxidase isoenzymes in chloroplasts and mitochondria of cucumber leaves as influenced by chilling. Photosynthetica. 2008, 46, 581–588. [Google Scholar] [CrossRef]
- Sagrillo, M.R.; Garcia, L.F.M.; De Souza Filho, O.C.; Duarte, M.M.M.F.; Ribeiro, E.E.; Cadon, F.C.; Da Cruz, I.B.M. Tucum fruit extracts (Astrocaryum aculeatum Meyer) decrease cytotoxic effects of hydrogen peroxide on human lymphocytes. Food Chem. 2015, 173, 741–748. [Google Scholar] [CrossRef]
- Kraujalienė, V.; Pukalskas, A.; Kraujalis, P.; Venskutonis, P.R. Biorefining of Bergenia crassifolia L. roots and leaves by high pressure extraction methods and evaluation of antioxidant properties and main phytochemicals in extracts and plant material. Ind. Crop Prod. 2016, 89, 390–398. [Google Scholar] [CrossRef]
- Adam, Z.; Khamis, S.; Ismail, A.; Hamid, M. Ficus deltoidea: A potential alternative medicine for diabetes mellitus. Evid. Based Complementary Altern. Med. 2012, 2012, 632763. [Google Scholar] [CrossRef]
- Bi, X.; Zhang, J.; Chen, C.; Zhang, D.; Li, P.; Ma, F. Anthocyanin contributes more to hydrogen peroxide scavenging than other phenolics in apple peel. Food Chem. 2014, 152, 205–209. [Google Scholar] [CrossRef]
- Jaakola, L.; Määttä, K.; Pirttilä, A.M.; Törrönen, R.; Kärenlampi, S.; Hohtola, A. Expression of genes involved in anthocyanin biosynthesis in relation to anthocyanin, proanthocyanidin, and flavonol levels during bilberry fruit development. Plant Physiol. 2002, 130, 729–739. [Google Scholar] [CrossRef]
- Zhang, R.Q.; Zhu, H.H.; Zhao, H.Q.; Yao, Q. Arbuscular mycorrhizal fungal inoculation increases phenolic synthesis in clover roots via hydrogen peroxide, salicylic acid and nitric oxide signaling pathways. J. Plant Physiol. 2013, 170, 74–79. [Google Scholar] [CrossRef]
- Abdel-Rahman, R.F.; Ezzat, S.M.; Ogaly, H.A.; Abd-Elsalam, R.M.; Hessin, A.F.; Fekry, M.I.; Mansour, D.F.; Mohamed, S.O. Ficus deltoidea extract down-regulates protein tyrosine phosphatase 1 expression in a rat model of type 2 diabetes mellitus: A new insight into its antidiabetic mechanism. J. Nutr. Sci. 2020, 9, e2. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| H2O2 (mM) | Leaf Chlorophyll (CCI) | Net Photosynthesis (μmol/m2/s) | Stomatal Conductance (m2s/mol) | Quantum Yield (Fv/Fm) | Plant Height (cm) | Leaf Area (cm2) |
|---|---|---|---|---|---|---|
| 0 | 76 c | 2.5 b | 360 b | 0.77 b | 31 b | 7 a |
| 8 | 80 b | 2.4 b | 320 b | 0.81 a | 32 b | 7 a |
| 16 | 81 b | 3.6 a | 300 b | 0.80 a | 35 a | 8 a |
| 30 | 89 a | 4.5 a | 400 a | 0.80 a | 38 a | 9 a |
| 60 | 79 b | 2.2 b | 080 c | 0.79 a | 32 b | 8 a |
| H2O2 (mM) | IC50 (µg/mL) | ||
|---|---|---|---|
| Ethanolic Extract | Chloroform Extract | Aqueous Extract | |
| 0 | 76.33 ± 0.67 c | - | 95.67 ± 0.88 c |
| 8 | 86.33 ± 1.20 d | - | 95.33 ± 1.20 c |
| 16 | 25.00 ± 1.53 a | - | 116.33 ± 0.67 d |
| 30 | 61.67 ± 0.88 b | 201.67 ± 0.88 a | 45.00 ± 1.15 a |
| 60 | 75.00 ± 1.15 c | 290.77 ± 0.67 b | 80.00 ± 0.218 b |
| Gallic Acid | 3.50 ± 0.25 | 3.50 ± 0.25 | 3.50 ± 0.25 |
| Quercetin | 3.67 ± 0.08 | 3.67 ± 0.08 | 3.67 ± 0.08 |
| Ascorbic Acid | 4.92 ± 0.08 | 4.92 ± 0.08 | 4.92 ± 0.08 |
| H2O2 (mM) | IC50 (µg/mL) |
|---|---|
| α-Glucosidase Activity | |
| 0 | 25.0 ± 0.77 b |
| 8 | 60.0 ± 0.11 d |
| 16 | 100.0 ± 0.81 e |
| 30 | 40.0 ± 0.10 c |
| 60 Quercetin Acarbose | 57.5 ± 0.67 g 12.5 ± 0.15 a 235.0 ± 0.66 f |
| Regression Equation | Correlation Coefficient (r) | Coefficients of Determination (R2) |
|---|---|---|
| Relationship between antioxidant activity with total phenolic content | ||
| YEE = 1.4464x + 244.08 | r = 0.783 | R2 = 0.6138 |
| YCE = −0.062x + 55.475 | r = 0.848 | R2 = 0.7189 |
| YAE = 3.2971x + 20.955 | r = 0.695 | R2 = 0.4832 |
| Relationship between antioxidant activity with total flavonoid content | ||
| YEE = 0.5116x + 105.07 | r = 0.870 | R2 = 0.7573 |
| YCE = 0.0025x + 6.2605 | r = 0.886 | R2 = 0.7851 |
| YAE = 0.8122x + 65.338 | r = 0.708 | R2 = 0.5017 |
| Relationship between antioxidant activity with α-glucosidase activity | ||
| YEE = −0.2276x + 64.061 | r = 0.332 | R2 = 0.1105 |
| H2O2 (mM) | Vitexin (ng/ µg) | % in Extract | Isovitexin (ng/ µg) | % in Extract |
|---|---|---|---|---|
| 0 | 135.63 ± 0.09 e | 0.27 | 50.93 ± 0.20 b | 0.10 |
| 8 | 138.30 ± 0.40 d | 0.28 | 42.18 ± 0.22 d | 0.08 |
| 16 | 137.23 ± 0.23 c | 0.27 | 39.93 ± 0.23 e | 0.08 |
| 30 | 144.27 ± 0.17 b | 0.29 | 72.44 ± 0.17 a | 0.14 |
| 60 | 152.53 ± 0.19 a | 0.31 | 44.90 ± 0.19 c | 0.09 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nurnaeimah, N.; Mat, N.; Suryati Mohd, K.; Badaluddin, N.A.; Yusoff, N.; Sajili, M.H.; Mahmud, K.; Mohd Adnan, A.F.; Khandaker, M.M. The Effects of Hydrogen Peroxide on Plant Growth, Mineral Accumulation, as Well as Biological and Chemical Properties of Ficus deltoidea. Agronomy 2020, 10, 599. https://doi.org/10.3390/agronomy10040599
Nurnaeimah N, Mat N, Suryati Mohd K, Badaluddin NA, Yusoff N, Sajili MH, Mahmud K, Mohd Adnan AF, Khandaker MM. The Effects of Hydrogen Peroxide on Plant Growth, Mineral Accumulation, as Well as Biological and Chemical Properties of Ficus deltoidea. Agronomy. 2020; 10(4):599. https://doi.org/10.3390/agronomy10040599
Chicago/Turabian StyleNurnaeimah, Nik, Nashriyah Mat, Khamsah Suryati Mohd, Noor Afiza Badaluddin, Nornasuha Yusoff, Mohammad Hailmi Sajili, Khairil Mahmud, Ahmad Faris Mohd Adnan, and Mohammad Moneruzzaman Khandaker. 2020. "The Effects of Hydrogen Peroxide on Plant Growth, Mineral Accumulation, as Well as Biological and Chemical Properties of Ficus deltoidea" Agronomy 10, no. 4: 599. https://doi.org/10.3390/agronomy10040599
APA StyleNurnaeimah, N., Mat, N., Suryati Mohd, K., Badaluddin, N. A., Yusoff, N., Sajili, M. H., Mahmud, K., Mohd Adnan, A. F., & Khandaker, M. M. (2020). The Effects of Hydrogen Peroxide on Plant Growth, Mineral Accumulation, as Well as Biological and Chemical Properties of Ficus deltoidea. Agronomy, 10(4), 599. https://doi.org/10.3390/agronomy10040599