Influence of Electromagnetic Stimulation of Seeds on the Photosynthetic Indicators in Medicago sativa L. Leaves at Various Stages of Development

 ,
,  and
and 
Abstract
:1. Introduction
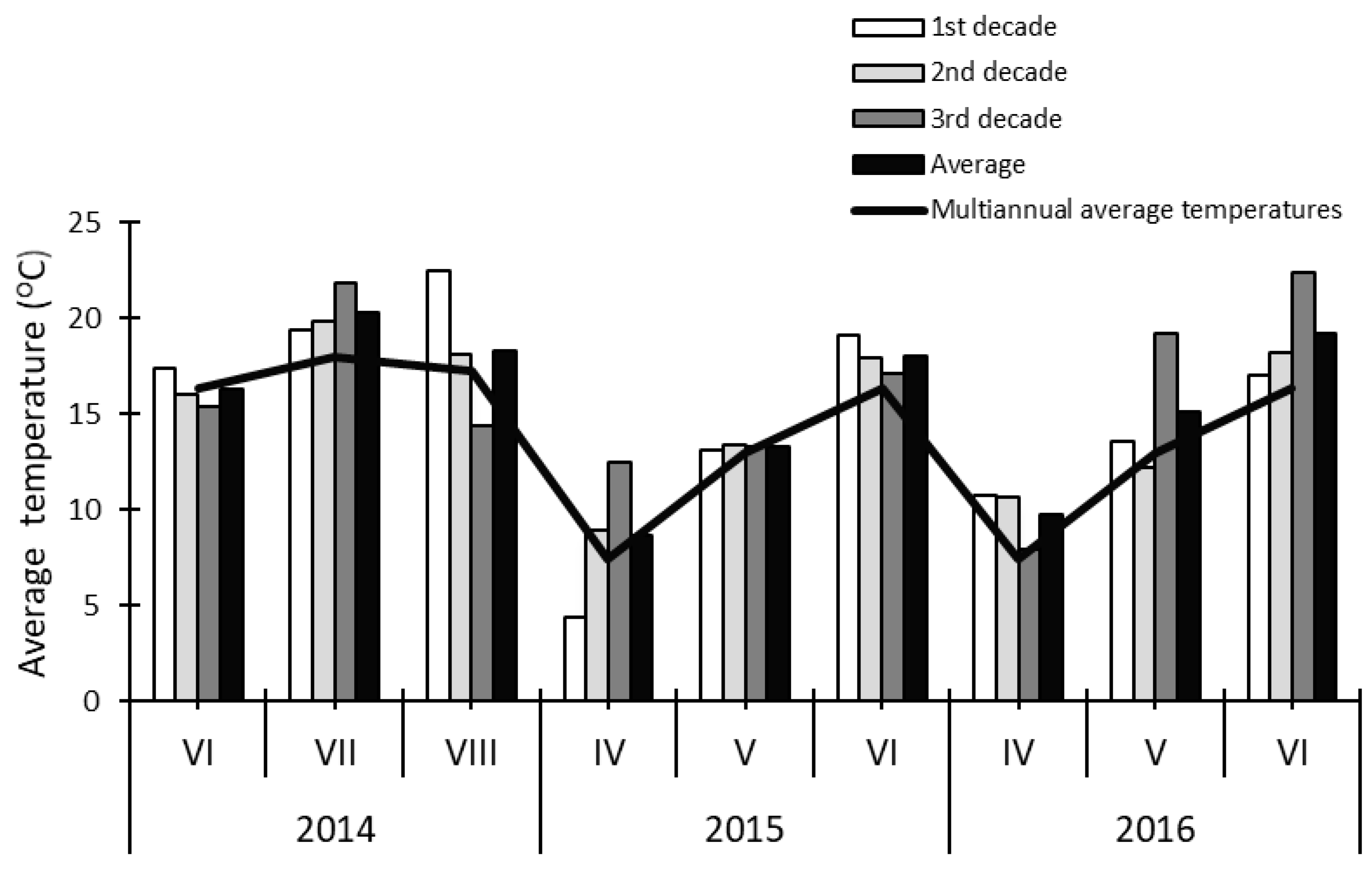
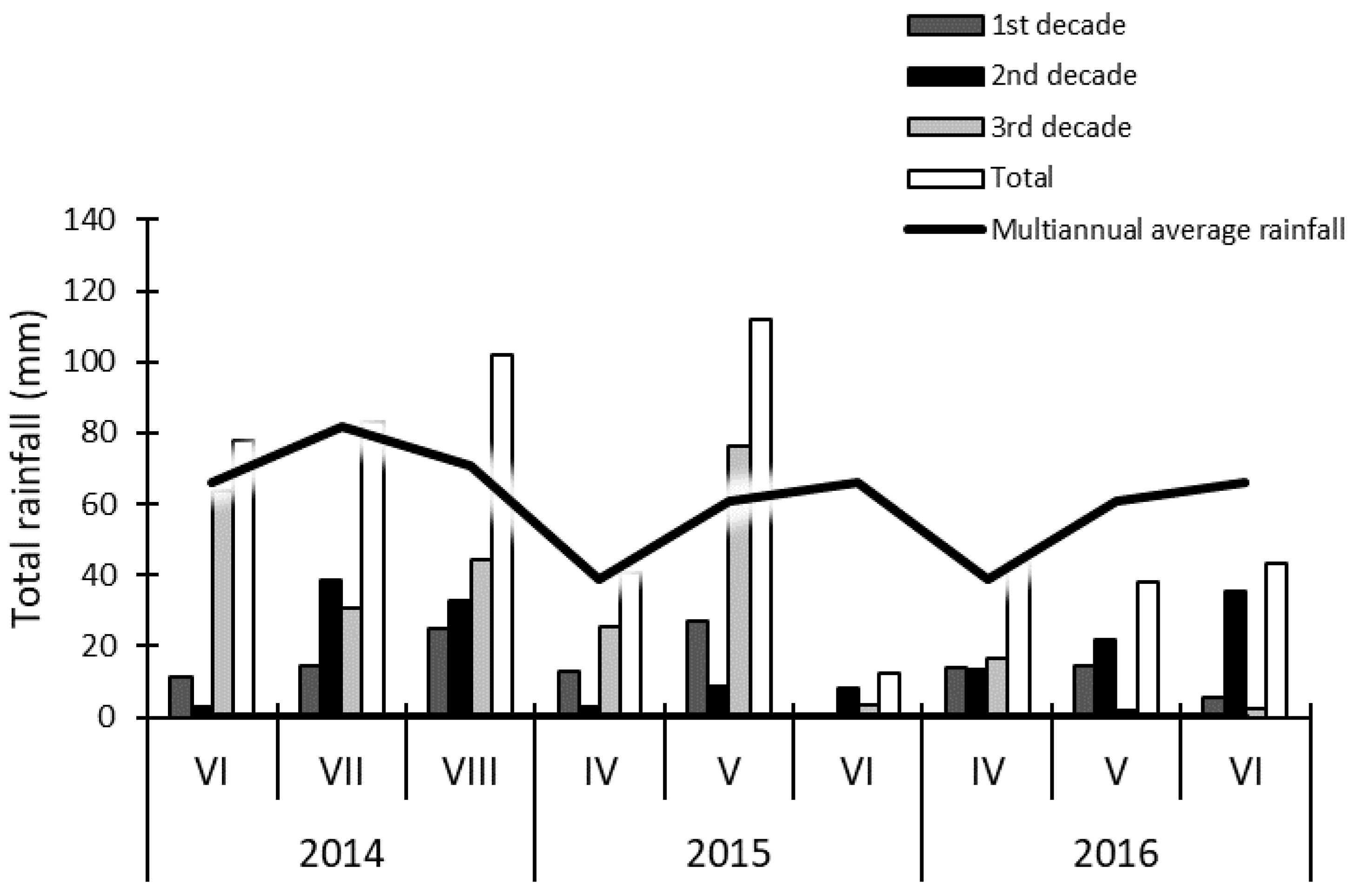
2. Materials and Methods
3. Results and Discussion
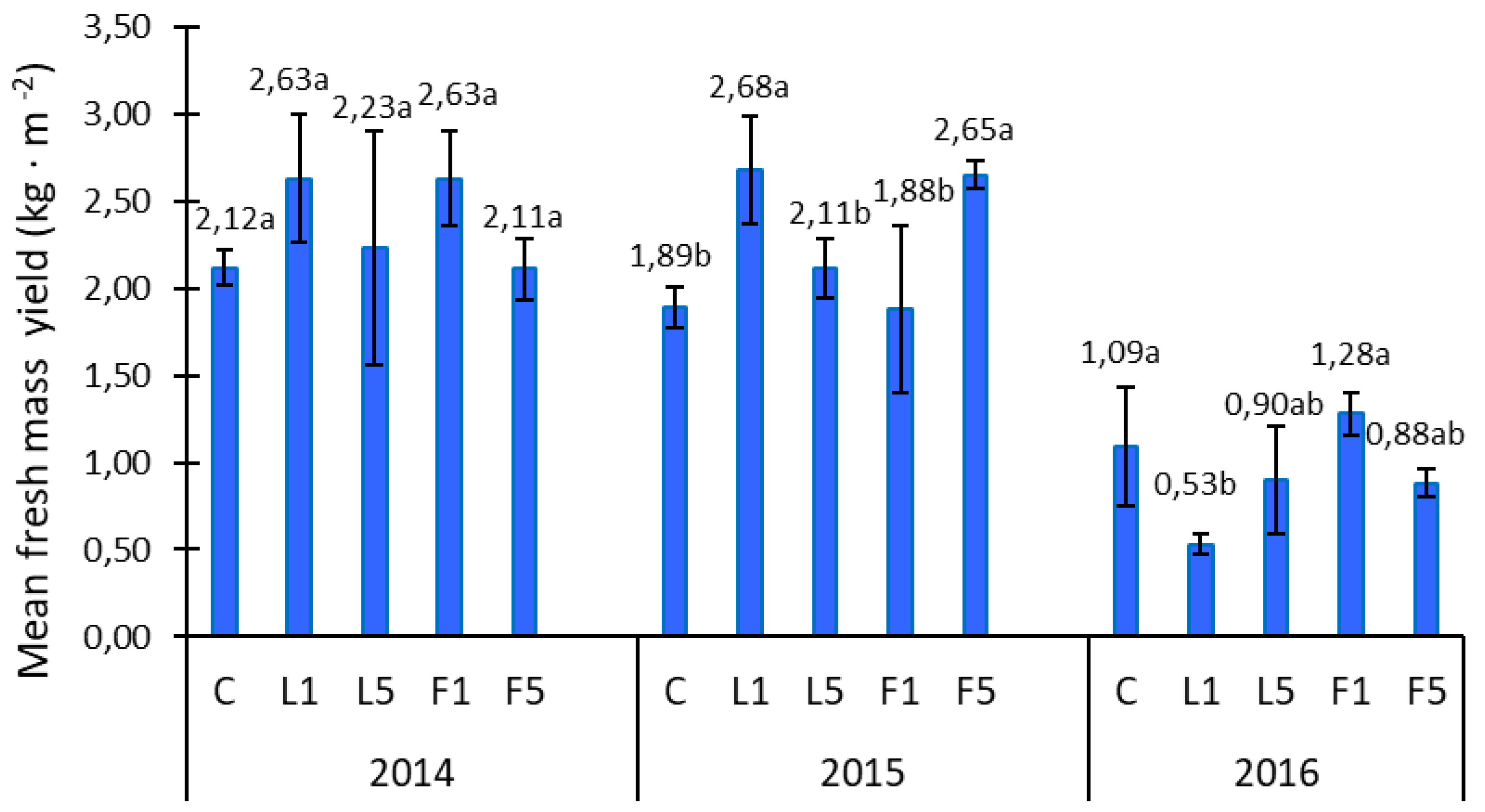
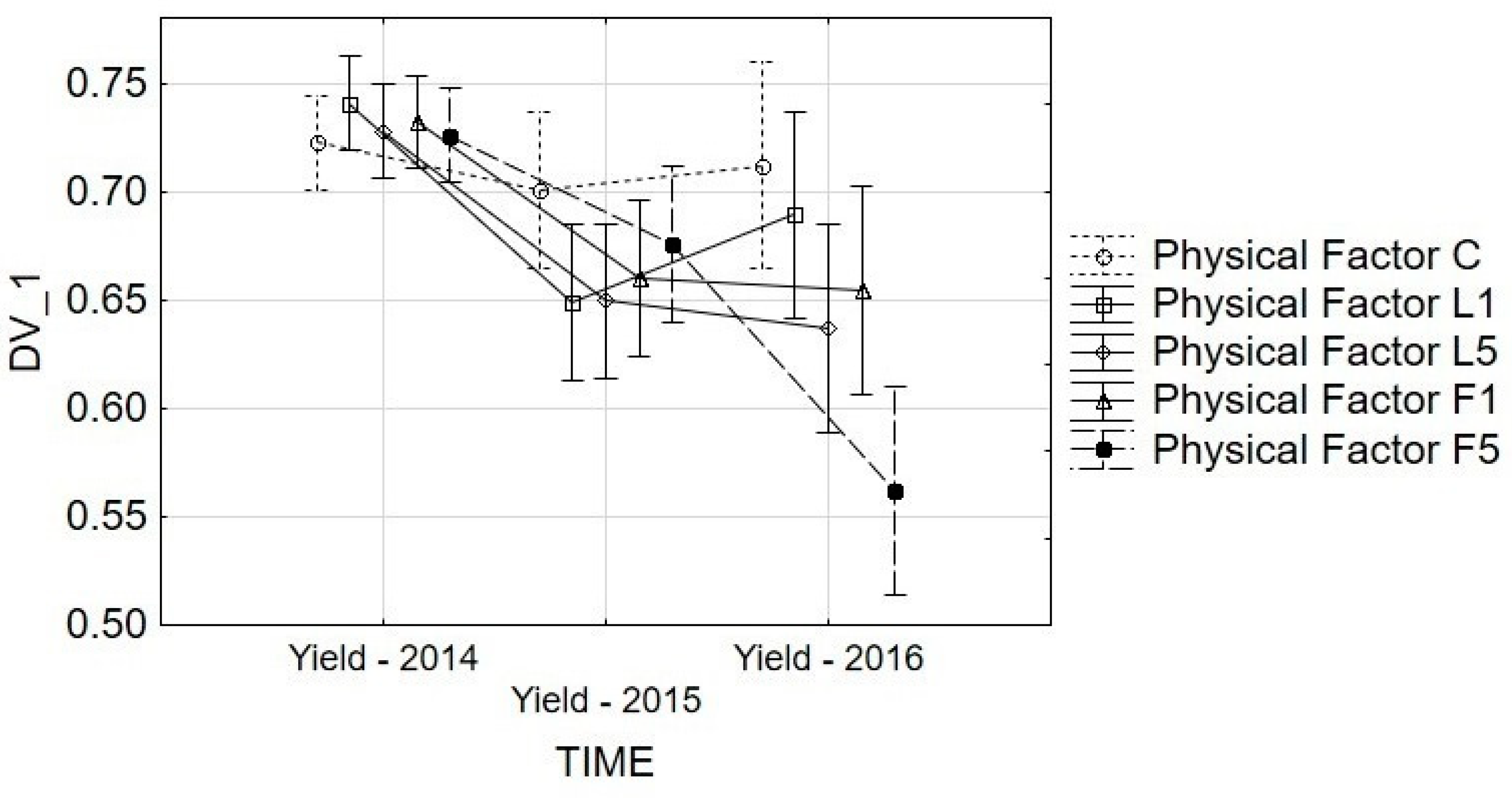
3.1. Yield and ETR Analysis
- H0—Yield (ETR) does not change in time;
- H0—Yield (ETR) does not depend on the physical factor;
- Yield (ETR) does not depend on the stage of vegetation.
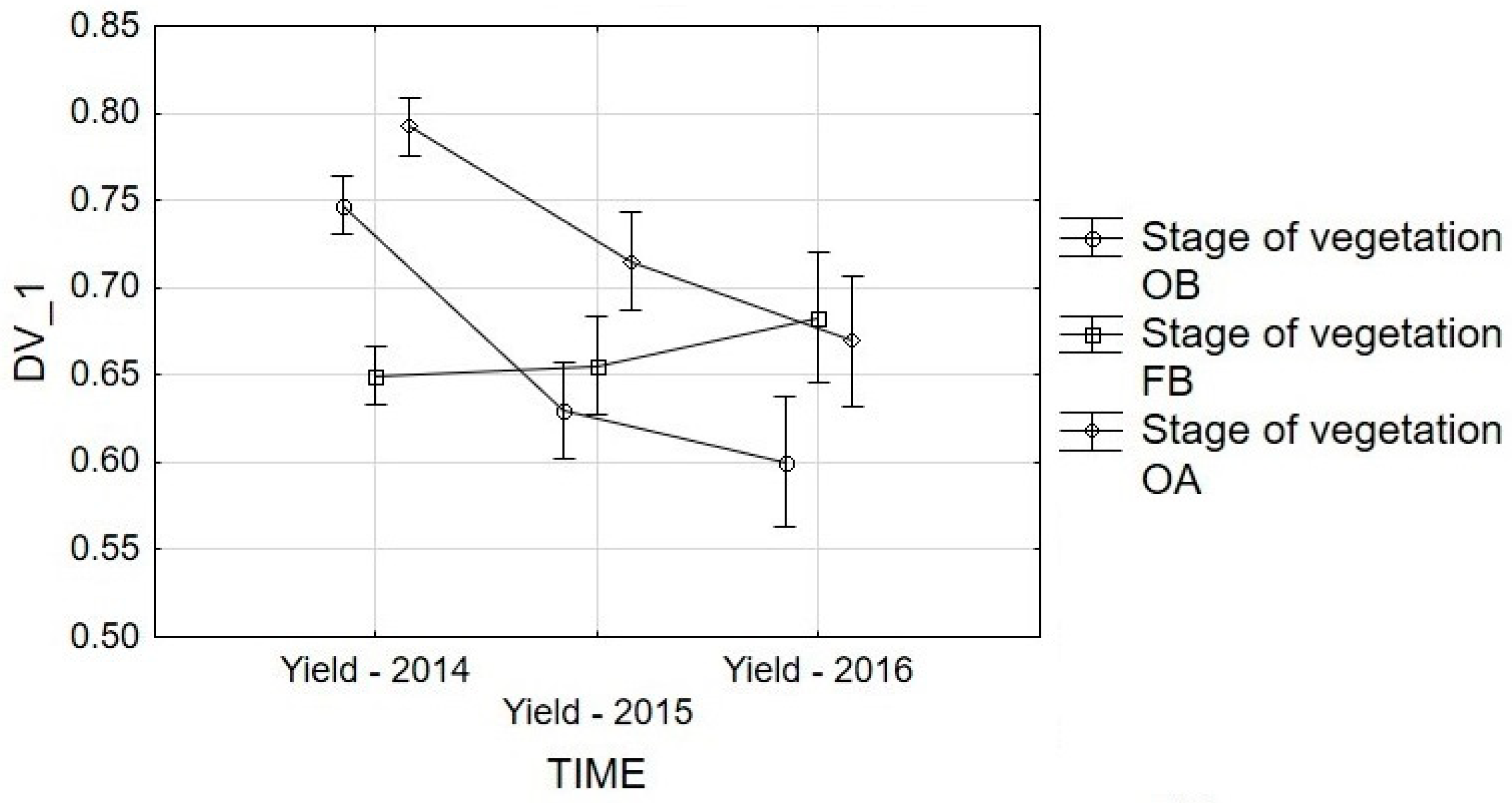
3.2. Yield Analysis
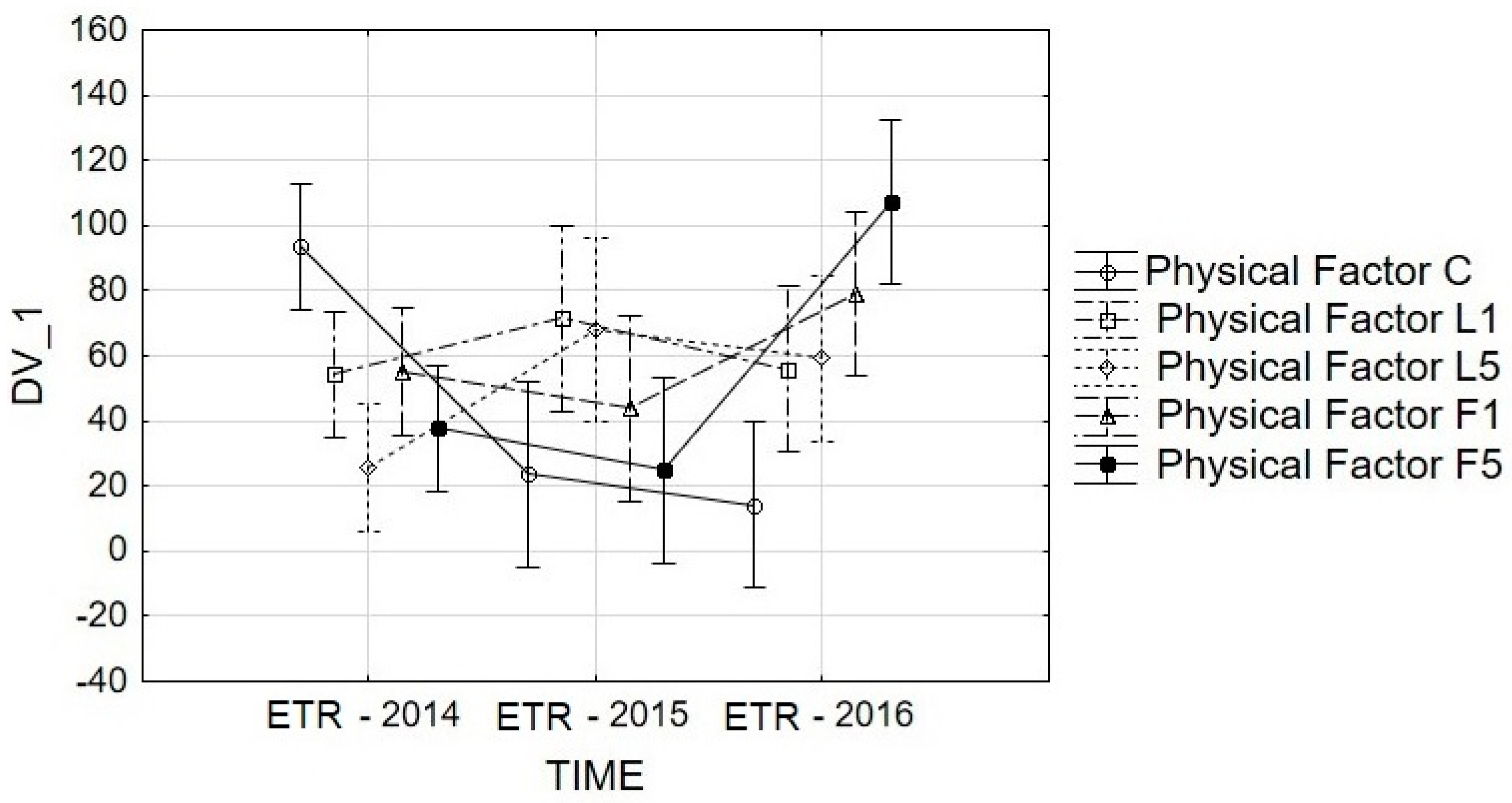
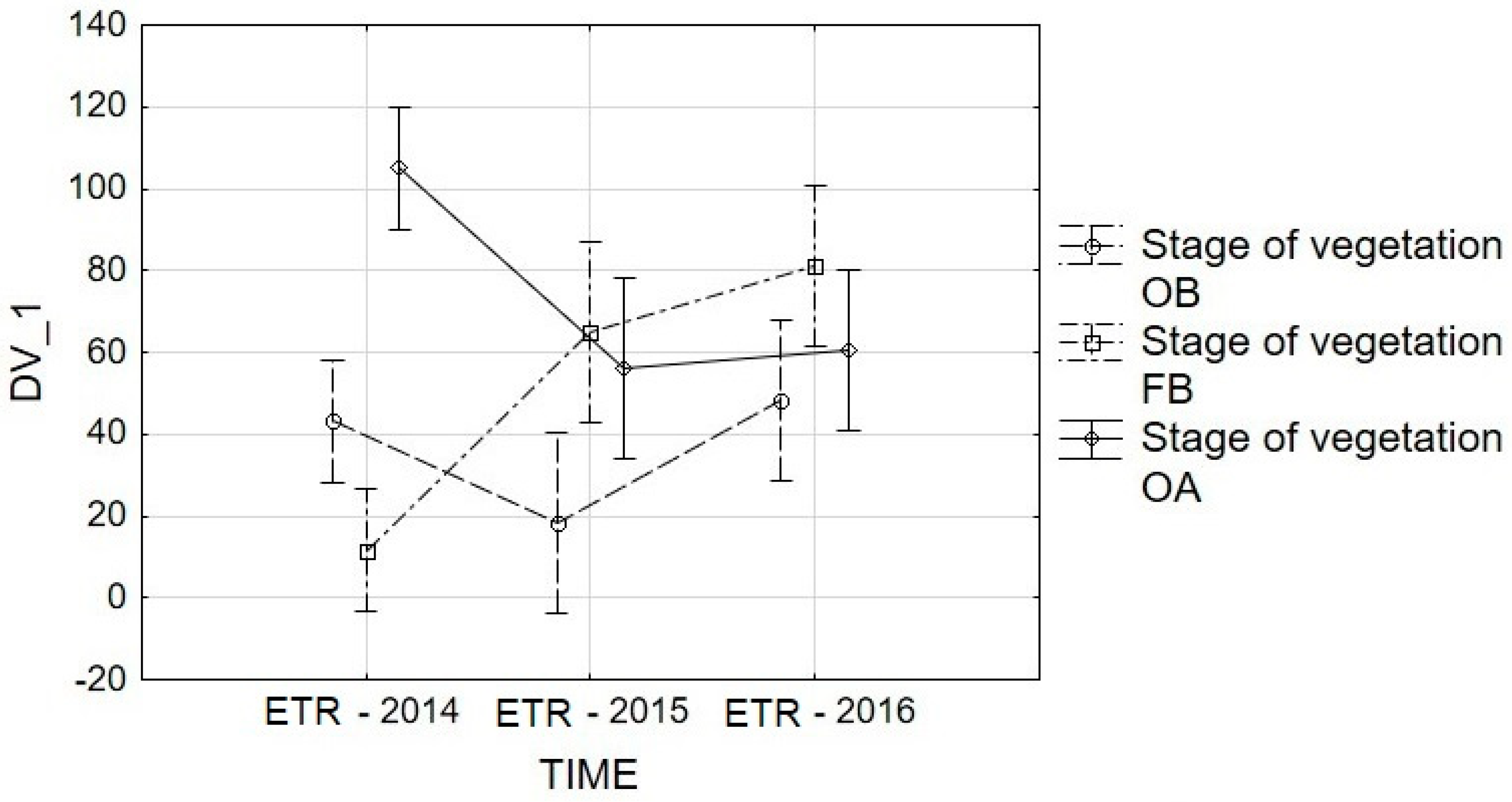
3.3. ETR Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Annicchiarico, P.; Pecetti, L.; Tava, A. Physiological and morphological traits associated with adaptation of lucerne (Medicago sativa) to severely drought-stressed and to irrigated environments. Ann. Appl. Biol. 2013, 162, 27–40. [Google Scholar] [CrossRef]
- Hayes, R.; Li, G.D.; Conyers, M.K.; Virgona, J.M.; Dear, B.S. Lime increases productivity and the capacity of Lucerne (Medicago sativa L.) and phalaris (Phalaris aquatic L.) to utilize stored soil water on an acidic soil in south-eastern Australia. Plant Soil 2016, 400, 29–43. [Google Scholar] [CrossRef]
- Le, X.H.; Franco, C.M.; Ballard, R.A.; Drew, E.A. Isolation and characterization of endophytic actinobacteria and their effect on the early growth and nodulation of lucerne (Medicago sativa L.). Plant Soil 2016, 405, 13–24. [Google Scholar] [CrossRef]
- He, S.; Liu, G.; Yang, H. Water Use Efficiency by Alfalfa: Mechanisms Involving Anti-Oxidation and Osmotic Adjustment under Drought. Russ. J. Plant Physiol. 2012, 59, 348–355. [Google Scholar] [CrossRef]
- Marley, C.L.; Fychan, R.; Theobald, V.J.; Cuttle, S.P.; Sanderson, R. Effects of a winter or spring sowing date on soil nitrogen utilization and yield of barley following a forage crop of red clover, lucerne or hybrid ryegrass. Agric. Ecosyst. Environ. 2013, 181, 213–222. [Google Scholar] [CrossRef]
- Bilodeau, S.E.; Wu, B.S.; Rufyikiri, A.S.; MacPherson, S.; Lefsrud, M. An update on plant photobiology and implications for cannabis production. Front. Plant Sci. 2019, 10, 296. [Google Scholar] [CrossRef]
- Hall, D.O.; Rao, K.K. Fotosynteza; WNT: Warszawa, Poland, 1999. (In Polish) [Google Scholar]
- Kalaji, M.H.; Łoboda, T. Fluorescencja Chlorofilu w Badaniach Stanu Fizjologicznego Roślin Wydawnictwo; SGGW: Warszawa, Poland, 2010. (In Polish) [Google Scholar]
- Alves, P.L.; Barja, P.R.; Magalhães, A.C.N. The phenomenon of photoinhibition of photosynthesis and its importance in reforestation. Bot. Rev. 2002, 68, 193–208. [Google Scholar] [CrossRef]
- Ptushenko, V.V.; Ptushenko, O.S.; Tikhonov, A.N. Chlorophyll fluorescence induction, chlorophyll content and chromaticity characteristics of leaves as in Arboreous plants. Biochemistry 2014, 79, 260–272. [Google Scholar]
- Sulkiewicz, M.; Ciereszko, I. Fluorescencja chlorofilu a–historia odkrycia i zastosowanie w badaniach roślin. KOSMOS Probl. Nauk Biol. 2016, 65, 103–115. (In Polish) [Google Scholar]
- Chen, Y.P.; Jia, J.F.; Yue, M. Effect of CO2 Laser Radiation on Physiological Tolerance of Wheat Seedlings Exposed to Chilling Stress. Photochem. Photobiol. 2010, 86, 600–605. [Google Scholar] [CrossRef]
- Hernandez, A.C.; Domínguez, P.A.; Cruz-Orea, A.; Ivanov, R.; Carballo, C.A.; Zepeda, B.R.; Galindo, S.L. Laser irradiation effects on field performance of maize seed genotypes. Int. Agrophys. 2009, 23, 327–332. [Google Scholar]
- Hernández, A.C.; Domínguez, P.A.; Cruz, O.A.; Ivanov, R.; Carballo, C.A.; Zepeda, B.R. Laser in agriculture. Int. Agrophys. 2010, 24, 407–422. [Google Scholar]
- Perveen, R.; Al, I.Q.; Ashraf, M.; Al-Qurainy, F.; Jamil, Y.; Ahmad, M.R. Effects of Different Doses of Low Power Continuous Wave He-Ne Laser Radiation on Some Seed Thermodynamic and Germination Parameters, and Potential Enzymes Involved in Seed Germination of Sunflower (Helianthus annuus L.). Photochem. Photobiol. 2010, 86, 1050–1055. [Google Scholar] [CrossRef] [PubMed]
- Soliman, A.S.H.; Harith, M.A. Effects of laser biostimulation on germination of Acacia Farnesiana (L.). Willd. Acta Hortic.(ISHS) 2010, 854, 41–50. [Google Scholar] [CrossRef]
- Dziwulska-Hunek, A.; Kornarzyński, K.; Matwijczuk, A.; Pietruszewski, S.; Szot, B. Effect of laser and variable magnetic field simulation on amaranth seeds germination. Int. Agrophys. 2009, 23, 229–235. [Google Scholar]
- Matwijczuk, A.; Kornarzyński, K.; Pietruszewski, S. Effect of magnetic field on seed germination and seedling growth of sunflower. Int. Agrophys. 2012, 26, 271–278. [Google Scholar] [CrossRef]
- Moon, J.D.; Chung, H.S. Acceleration of germination of tomato seed by applying AC electric and magnetic fields. J. Electrost. 2000, 48, 103–114. [Google Scholar] [CrossRef]
- Nechitailo, G.; Gordeev, A. The use of an electric field in increasing the resistance of plants to the action of unfavorable space flight factors. Adv. Space Res. 2004, 34, 1562–1565. [Google Scholar] [CrossRef]
- Cakmak, T.; Rahmi, D.; Serkan, E. Acceleration of germination and early growth of wheat and bean seedlings grown under various magnetic field and osmotic conditions. Bioelectromagnetics 2010, 31, 120–129. [Google Scholar] [CrossRef]
- Kataria, S.; Baghel, L.; Guruprasad, K.N. Pre-treatment of seeds with static magnetic field improves germination and early growth characteristics under salt stress in maize and soybean. Biocatal. Agric. Biotechnol. 2017, 10, 83–90. [Google Scholar] [CrossRef]
- Vashisth, A.; Nagarajan, S. Exposure of Seeds to Static Magnetic Field Enhances Germination and Early Growth Characteristics in Chickpea (Cicer arietinum L.). Bioelectromagnetics 2008, 29, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Yano, A.; Ohashi, Y.; Hirasaki, T.; Fujiwara, K. Effects of a 60 Hz Magnetic Field on Photosynthetic CO2 uptake and Early Growth of Radish Seedlings. Bioelectromagnetics 2004, 25, 572–581. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, P.A.; Hernandez, A.C.; Cruz, O.A.; Ivanov, R.; Carballo, C.A.; Zepeda, B.R.; Martínez, O.E. Influences of the electromagnetic field in maize seed vigor. Rev. Fitotec. Mex. 2010, 33, 183–188. (In Spanish) [Google Scholar]
- Galland, P.; Pazur, A. Magnetoreception in plants. J. Plant Res. 2005, 118, 371–389. [Google Scholar] [CrossRef]
- Hernandez, A.C.; Dominguez, P.A.; Carballo, C.A.; Cruz, O.A.; Ivanov, R.; López, B.J.L.; Valcarcel, M.J.P. Alternating magnetic field irradiation effects on three genotype maize seed field performance. Acta Agrophys. 2009, 170, 7–17. [Google Scholar]
- Zepeda, B.R.; Hernández, A.C.; Domínguez, P.A.; Cruz, O.A.; Godina, N.J.J.; Martínez, O.E. Electromagnetic field and seed vigour of corn hybrids. Int. Agrophys. 2010, 4, 329–332. [Google Scholar]
- Bujak, K.; Frant, M. Influence of pre-sowing seed stimulation with magnetic Field on spring wheat yielding. Acta Agrophys. 2009, 14, 19–29. (In Polish) [Google Scholar]
- Nimmi, V.; Madhu, G. Effect of pre-sowing treatment with permanent magnetic field on germination and growth of chilli (Capsicum annum. L.). Int. Agrophys. 2009, 23, 195–198. [Google Scholar]
- Pietruszewski, S.; Muszyński, S.; Dziwulska, A. Electromagnetic fields and electromagnetic radiation as non-invasive external simulations for seeds (selected methods and responses). Int. Agrophys. 2007, 21, 95–100. [Google Scholar]
- Greenebaum, B.; Barnes, F. Bioengineering and Biophysical Aspects of Electromagnetic Fields; CRC Press: Boca Raton, FL, USA, 2018; Available online: https://doi.org/10.1201/9781315186580 (accessed on 10 April 2020).
- Gutiérrez Cruz, D.; Zepeda Bautista, R.; Hernández Aguilar, C.; Domínguez Pacheoo, F.A.; Cruz Orea, A.; López Bonilla, J.L. Physical characteristic of grains of maize pre-sowing treated by electromagnetic fields. Acta Agrophys. 2011, 18, 17–31. [Google Scholar]
- Nobel, P. Physicochemical and Environmental Plant Physiology, 4th ed.; Akademic Press: Cambridge, MA, USA, 2009. [Google Scholar]
- Krawiec, M.; Dziwulska-Hunek, A.; Kornarzyński, K. The use of physical factors for seed quality improvement of horticultural plants. J. Hortic. Res. 2018, 26, 81–94. [Google Scholar] [CrossRef] [Green Version]
- Gładyszewska, B. Estimation of a laser biostimulation dose. Int. Agrophys. 2011, 25, 403–405. [Google Scholar]
- Vasilevski, G. Perspectives of the application of biophysical methods in sustainable agriculture. Bulg. J. Plant Physiol. 2003, 29, 179–186. [Google Scholar]
- Belyavskaya, N.A. Biological effects due to weak magnetic field on plants. Adv. Space Res. 2004, 34, 1566–1574. [Google Scholar] [CrossRef]
- Aksenov, S.I.; Buluchev, A.A.; Grunina, T.Y.; Turovetskii, V.B. Mechanisms of the action of a low-frequency magnetic field on the initial stages of germination of wheat seeds. Biophysics 1996, 41, 931–937. [Google Scholar]
- García Reina, F.; Arza Pascual, L.; Almanza Fundora, I. Influence of a stationary magnetic field on water relations in lettuce seeds. Part II: Experimental results. Bioelectromagnetics 2001, 22, 596–602. [Google Scholar] [CrossRef]
- Nasiri, A.A.; Mortazaeinezhad, F.; Taheri, R. Seed germination of medicinal sage is affected by gibberellic acid, magnetic field and laser irradiation. Electromagn. Biol. Med. 2018, 37, 50–56. [Google Scholar] [CrossRef]
- Ćwintal, M.; Dziwulska-Hunek, A. Effect of electromagnetic stimulation of alfalfa seeds. Int. Agrophys. 2013, 27, 391–401. [Google Scholar] [CrossRef] [Green Version]
- Ćwintal, M.; Dziwulska-Hunek, A.; Wilczek, M. Laser stimulation effect of seeds on quality of alfalfa. Int. Agrophys. 2010, 24, 15–19. [Google Scholar]
- Sujak, A.; Dziwulska-Hunek, A.; Reszczyńska, E. Effect of electromagnetic stimulation on selected Fabaceae Plants. Pol. J. Environ. Stud. 2013, 22, 893–898. [Google Scholar]
- Muszyński, S.; Gagoś, M.; Pietruszewski, S. Short-term pre-germination exposure to ELF magnetic field does not influence seedling growth in Durum Wheat (Triticum durum). Pol. J. Environ. Stud. 2009, 18, 1065–1072. [Google Scholar]
- Center, M.D.; Dąbrowski, P.; Samborska, I.A.; Łukasik, I.; Sowczyna, T.; Pietkiewicz, S.; Bąba, W.; Kalaji, H.M. Application of chlorophyll fluorescence measurements in environmental studies (in Polish). Kosmos Probl. Nauk Biol. 2016, 65, 197–205. [Google Scholar]
- Lichtenthalter, H.K.; Buschmann, C. Chlorophylls and Carotenoids: Measurement and Characterization by UV-Vis Spectroscopy. In Current Protocols in Food Analytical Chemistry; Wiley & Sons. Inc.: Hoboken, NJ, USA, 2001. [Google Scholar]
- Brereton, R.G. Chemometrics: Data Analysis for the Laboratory and Chemical Plant; John Wiley: Chichester, UK, 2003. [Google Scholar]
- Ni, Y.; Guo, Y.J.; Guo, Y.J.; Han, L.; Tang, H.; Conyers, M. Leaf cuticular waxes and physiological parameters in alfalfa leaves as influenced by drogught. Photosynthetica 2012, 50, 458–466. [Google Scholar] [CrossRef]
- Erice, G.; Louahlia, S.; Irigogen, J.J.; Sánchez-Díaz, M.; Alami, I.T.; Avice, J.C. Water use efficiency, transpiration and net CO2 exchange of four alfalfa genotypes submitted to progressive drought and subsequent recovery. Environ. Exp. Bot. 2011, 72, 123–130. [Google Scholar] [CrossRef]
- Dziwulska-Hunek, A.; Ćwintal, M.; Niemczynowicz, A.; Boroń, B.; Matwijczuk, A. Effect of stress caused by electromagnetic stimulation on the fluorescence lifetime of chlorophylls in alfalfa leaves. Pol. J. Environ. Stud. 2019, 28, 3133–3143. [Google Scholar] [CrossRef]
- Schurr, U.; Walter, A.; Rascher, U. Functional dynamic of plant growth and photosynthesis—From steady state to dynamics—From homogeneity to heterogeneity. Plant Cell Environ. 2006, 29, 340–352. [Google Scholar] [CrossRef]
- Baker, N.R.; Rosenqvist, E. Applications of chlorophyll fluorescence can improve crop production strategies: An examination of future possibilities. J. Exp. Bot. 2004, 55, 1607–1621. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Zhang, S.; Shan, L. Responsibility of non-stomatal limitations for the reduction of photosynthesis—Response of photosynthesis and antioxidant enzyme characteristics in alfalfa (Medicago sativa L.) seedlings to water stress and rehydration. Front. Agric. China 2007, 1, 255–264. [Google Scholar] [CrossRef]
- Smethurst, C.; Garnett, T.; Shabala, S. Nutritional and chlorophyll fluorescence responses of lucerne (Medicago sativa) to waterlogging and subsequent recovery. Plant Soil 2005, 270, 31–45. [Google Scholar] [CrossRef]
- Mouradi, M.; Bouizgaren, A.; Farissi, M.; Latrach, L.; Qaddoury, A.; Ghoulam, C. Seed osmopriming improves plant growth, nodulation, chlorophyll fluorescence and nutrient uptake in alfalfa (Medicago sativa L.)—Rhizobia symbiosis under drought stress. Sci. Hortic. 2016, 213, 232–242. [Google Scholar] [CrossRef]
- Hwang, S.F.; Wang, H.; Gossen, B.D.; Chang, K.F.; Turnbull, G.D.; Howard, R.J. Impact of foliar diseases on photosynthesis, protein content and seed yield of alfalfa and efficacy of fungicide application. Eur. J. Plant Pathol. 2006, 115, 389–399. [Google Scholar] [CrossRef]
- Mauro, R.P.; Occhipinti, A.; Longo, A.M.G.; Mauromicale, G. Effects of shading on chlorophyll content, chlorophyll fluorescence and photosynthesis of subterranean clover. J. Agron. Crop Sci. 2011, 197, 57–66. [Google Scholar] [CrossRef]
- Cetin, M. Change in Amount of chlorophyll in some interior ornamental plants. Kast. Univ. J. Eng. Sci. 2017, 3, 11–19. [Google Scholar]
- Singh, S.K.; Reddy, V.R.; Fleisher, D.H.; Timlin, D.J. Relationship between photosynthetic pigments and chlorophyll fluorescence in soybean under varying phosphorus nutrition at ambient and elevated CO2. Photosynthetica 2017, 55, 421–433. [Google Scholar] [CrossRef]
- Asghar, T.; Jamil, Y.; Iqbal, M.; Abbas, M. Laser light and magnetic field stimulation effect on biochemical, enzymes activities and chlorophyll contents in soybean seeds and seedlings during early growth stages. J. Photochem. Photobiol. B Biol. 2016, 165, 283–290. [Google Scholar] [CrossRef]
- Bielinis, E.; Jóźwiak, W.; Robakowski, P. Modeling of the relationship between the SPAD values and photosynthetic pigments content in Queraus petraea and Prunus serotina leaves. Dendrobiology 2015, 73, 125–134. [Google Scholar] [CrossRef] [Green Version]
- Khavari-Nejd, R.A.; Chaparzadeh, N. The effects of NaCl and CaCl2 on photosynthesis and growth of alfalfa plants. Photosynthetica 1998, 35, 461–466. [Google Scholar] [CrossRef]
- Owusu-Sekyere, A.; Kontturi, J.; Hajiboland, R.; Rahmat, S.; Aliasgharzad, N.; Hartikainen, H.; Seppänen, M.M. Influence of selenium (Se) on carbohydrate metabolism, nodulation and growth in alfalfa (Medicago sativa L.). Plant Soil 2013, 373, 541–552. [Google Scholar] [CrossRef]
- Lúcio, A.D.C.; Couto, M.R.M.; Lopes, S.J.; Storck, L. Transformação box-cox em experimentos com pimentão em ambiente protegido. Hortic. Bras. 2011, 29, 38–42. [Google Scholar] [CrossRef] [Green Version]
- Piepho, H.P. Data transformation in statistical analysis of field trials with changing treatment variance. Agron. J. 2009, 101, 865–869. [Google Scholar] [CrossRef]
- Sari, B.G.; Dal’Col Lúcio, A.; Santana, C.S.; Olivoto, T.; Diel, M.I.; Krysczun, D.K. Nonlinear growth models: An alternative to ANOVA in tomato trials evaluation. Eur. J. Agron. 2019, 104, 21–36. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Developmental Stage | BBCH Scale | Dates | Decade | Mean Decade Temperature (°C) | Total Rainfall in the Decade (mm) |
|---|---|---|---|---|---|---|
| 2014 | onset of budding | 51–54 | 26 June | 3 | 15.4 | 63.9 |
| full budding | 55–59 | 09 July | 1 | 19.4 | 14.2 | |
| onset of anthesis | 60–64 | 23 July | 3 | 21.8 | 30.6 | |
| 2015 | onset of budding | 51–54 | 01 May | 1 | 13.1 | 26.8 |
| full budding | 55–59 | 15 May | 2 | 13.4 | 8.9 | |
| onset of anthesis | 60–64 | 29 May | 3 | 13.3 | 76.2 | |
| 2016 | onset of budding | 51–54 | 26 April | 3 | 7.9 | 16.4 |
| full budding | 55–59 | 09 May | 1 | 13.6 | 14.2 | |
| onset of anthesis | 60–64 | 23 May | 3 | 19.2 | 2.1 |
| Specification | Photosynthetic Efficiency Yield (II) | ||||||
|---|---|---|---|---|---|---|---|
| C | L1 | L5 | F1 | F5 | Mean | ||
| Year | Developmental Stage | ||||||
| 2014 | onset of budding | 0.748a | 0.750a | 0.742a | 0.745a | 0.750a | 0.747B |
| full budding | 0.625a | 0.687a | 0.672a | 0.664a | 0.627a | 0.655C | |
| onset of anthesis | 0.795a | 0.801a | 0.769a | 0.798a | 0.800a | 0.793A | |
| 2015 | onset of budding | 0.634a | 0.618a | 0.659a | 0.611a | 0.614a | 0.627B |
| full budding | 0.705a | 0.616a | 0.627a | 0.674a | 0.643a | 0.653B | |
| onset of anthesis | 0.762a | 0.713a | 0.662a | 0.677a | 0.726a | 0.708A | |
| 2016 | onset of budding | 0.659a | 0.640a | 0.515b | 0.611a | 0.577a | 0.600A |
| full budding | 0.680a | 0.678a | 0.661a | 0.685a | 0.712a | 0.683A | |
| onset of anthesis | 0.797a | 0.751a | 0.735a | 0.668a | 0.398b | 0.670A | |
| Specification | ETR | ||||||
|---|---|---|---|---|---|---|---|
| C | L1 | L5 | F1 | F5 | Mean | ||
| Year | Developmental Stage | ||||||
| 2014 | onset of budding | 83.80a | 18.96b | 13.56b | 64.61a | 36.71b | 43.53B |
| full budding | 14.28a | 9.51b | 11.83a | 12.29a | 10.31b | 11.64B | |
| onset of anthesis | 182.69a | 141.58a | 51.80b | 83.02b | 66.23b | 105.06A | |
| 2015 | onset of budding | 25.22a | 17.48a | 14.35b | 14.48b | 18.74a | 18.05B |
| full budding | 24.30b | 138.22a | 70.91b | 60.98b | 35.14b | 65.91A | |
| onset of anthesis | 21.70a | 58.62a | 118.79b | 60.75a | 62.26a | 64.42A | |
| 2016 | onset of budding | 19.47b | 60.53b | 45.53b | 71.12a | 43.49b | 48.05A |
| full budding | 15.86b | 68.36a | 90.08a | 108.31a | 122.21a | 80.96A | |
| onset of anthesis | 7.58b | 38.91b | 41.73b | 57.27b | 156.59a | 60.42A | |
| Specification | Chlorophyll a (μg g−1 Fresh Weight) | ||||||
|---|---|---|---|---|---|---|---|
| C | L1 | L5 | F1 | F5 | Mean | ||
| Year | Developmental Stage | ||||||
| 2014 | onset of budding | 1849a | 1688a | 1349b | 2053a | 2399b | 1866A |
| full budding | 1915a | 2053b | 1936a | 2273b | 2131b | 2061A | |
| onset of anthesis | 1878b | 2090a | 2118a | 1697c | 1766c | 1910A | |
| 2015 | onset of budding | 2052a | 2244a | 1749b | 2094a | 1419b | 1912B |
| full budding | 964b | 1654a | 1606a | 1670a | 1439a | 1467B | |
| onset of anthesis | 2886a | 2753a | 3154a | 2205b | 2252b | 2650A | |
| 2016 | onset of budding | 2077b | 2482a | 1484c | 1927b | 1713c | 1937A |
| full budding | 2266a | 1688a | 2139a | 2225a | 2570a | 2178A | |
| onset of anthesis | 3584a | 2110b | 2078b | 1831b | 2786a | 2478A | |
| Specification | Chlorophyll b (μg g−1 Fresh Weight) | ||||||
|---|---|---|---|---|---|---|---|
| C | L1 | L5 | F1 | F5 | Mean | ||
| Year | Developmental Stage | ||||||
| 2014 | onset of budding | 566a | 504a | 414b | 632a | 723b | 568A |
| full budding | 598b | 644a | 593b | 704a | 687a | 645A | |
| onset of anthesis | 589b | 671a | 659a | 566b | 554b | 608A | |
| 2015 | onset of budding | 621a | 649a | 513b | 628a | 415c | 565B |
| full budding | 285b | 460a | 498a | 540a | 427a | 442B | |
| onset of anthesis | 962a | 895a | 1012a | 667b | 740b | 855A | |
| 2016 | onset of budding | 774b | 918a | 598c | 691b | 522c | 701A |
| full budding | 690a | 528a | 668a | 702a | 792a | 676A | |
| onset of anthesis | 1025a | 590b | 597b | 552b | 819a | 716A | |
| Specification | Carotenoids (μg g−1 Fresh Weight) | ||||||
|---|---|---|---|---|---|---|---|
| C | L1 | L5 | F1 | F5 | Mean | ||
| Year | Developmental Stage | ||||||
| 2014 | onset of budding | 187b | 194b | 158b | 235a | 323a | 220A |
| full budding | 172b | 258a | 206b | 244a | 204b | 217A | |
| onset of anthesis | 263a | 238a | 248a | 160c | 244a | 231A | |
| 2015 | onset of budding | 435a | 464a | 413a | 402a | 350c | 413A |
| full budding | 173b | 296a | 257a | 152b | 273a | 230B | |
| onset of anthesis | 387b | 470a | 444b | 329b | 348b | 395A | |
| 2016 | onset of budding | 333a | 305a | 154c | 221c | 208c | 244B |
| full budding | 267a | 171b | 218a | 278a | 345a | 256B | |
| onset of anthesis | 712a | 543a | 343b | 275c | 522c | 479A | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dziwulska-Hunek, A.; Kornarzyńska-Gregorowicz, A.; Niemczynowicz, A.; Matwijczuk, A. Influence of Electromagnetic Stimulation of Seeds on the Photosynthetic Indicators in Medicago sativa L. Leaves at Various Stages of Development. Agronomy 2020, 10, 594. https://doi.org/10.3390/agronomy10040594
Dziwulska-Hunek A, Kornarzyńska-Gregorowicz A, Niemczynowicz A, Matwijczuk A. Influence of Electromagnetic Stimulation of Seeds on the Photosynthetic Indicators in Medicago sativa L. Leaves at Various Stages of Development. Agronomy. 2020; 10(4):594. https://doi.org/10.3390/agronomy10040594
Chicago/Turabian StyleDziwulska-Hunek, Agata, Agnieszka Kornarzyńska-Gregorowicz, Agnieszka Niemczynowicz, and Arkadiusz Matwijczuk. 2020. "Influence of Electromagnetic Stimulation of Seeds on the Photosynthetic Indicators in Medicago sativa L. Leaves at Various Stages of Development" Agronomy 10, no. 4: 594. https://doi.org/10.3390/agronomy10040594
APA StyleDziwulska-Hunek, A., Kornarzyńska-Gregorowicz, A., Niemczynowicz, A., & Matwijczuk, A. (2020). Influence of Electromagnetic Stimulation of Seeds on the Photosynthetic Indicators in Medicago sativa L. Leaves at Various Stages of Development. Agronomy, 10(4), 594. https://doi.org/10.3390/agronomy10040594