Changes in Leaf Structure and Chemical Compositions Investigated by FTIR Are Correlated with Different Low Potassium Adaptation of Two Cotton Genotypes


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatments
2.2. Determination of SPAD Value and Photosynthetic Pigment Contents
2.3. Composition and Structure Analysis of Functional Leaves by FTIR
2.4. Statistical Analysis
3. Results
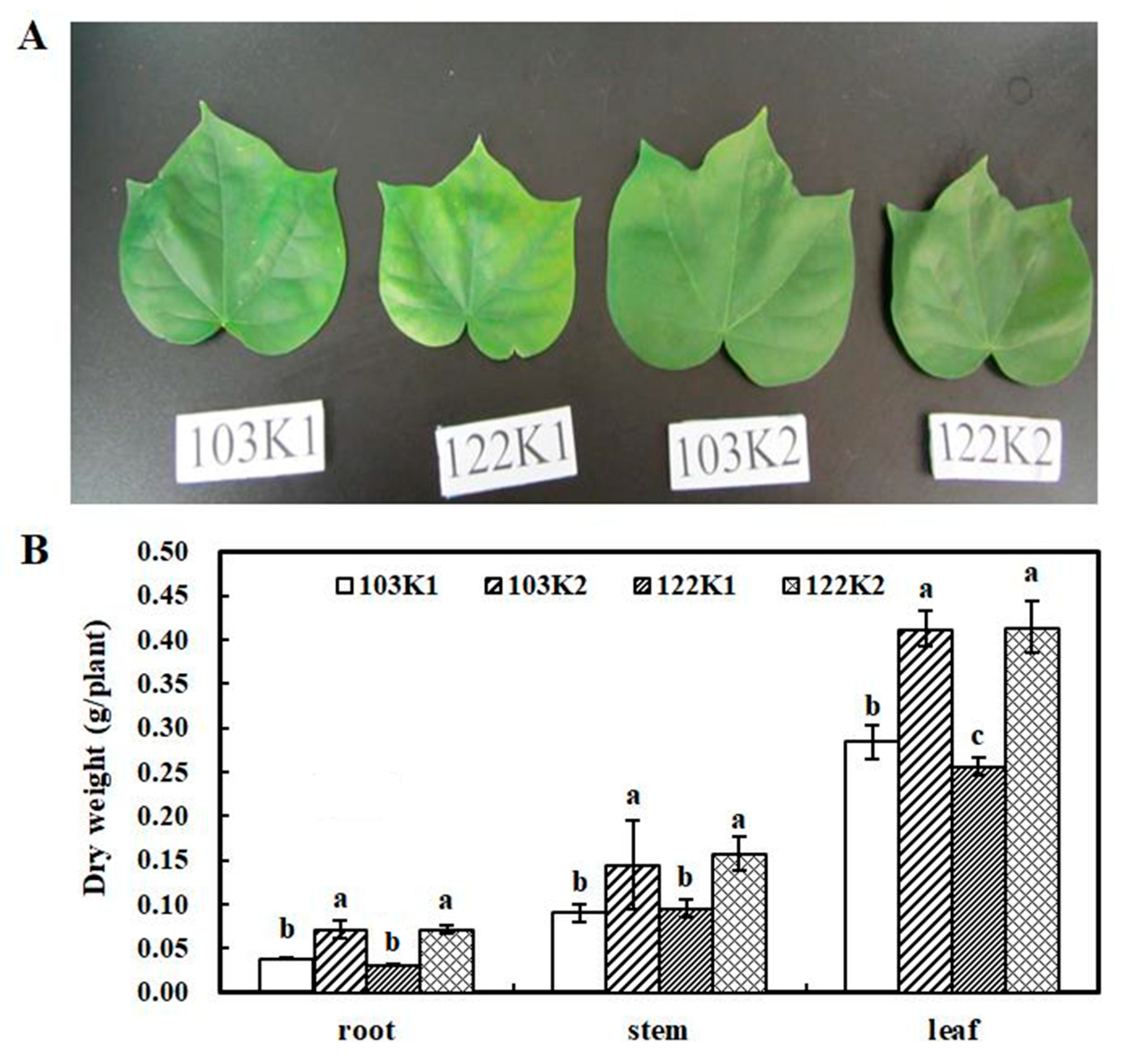
3.1. Plant Growth and Symptoms of Functional Leaves under Different K Treatments
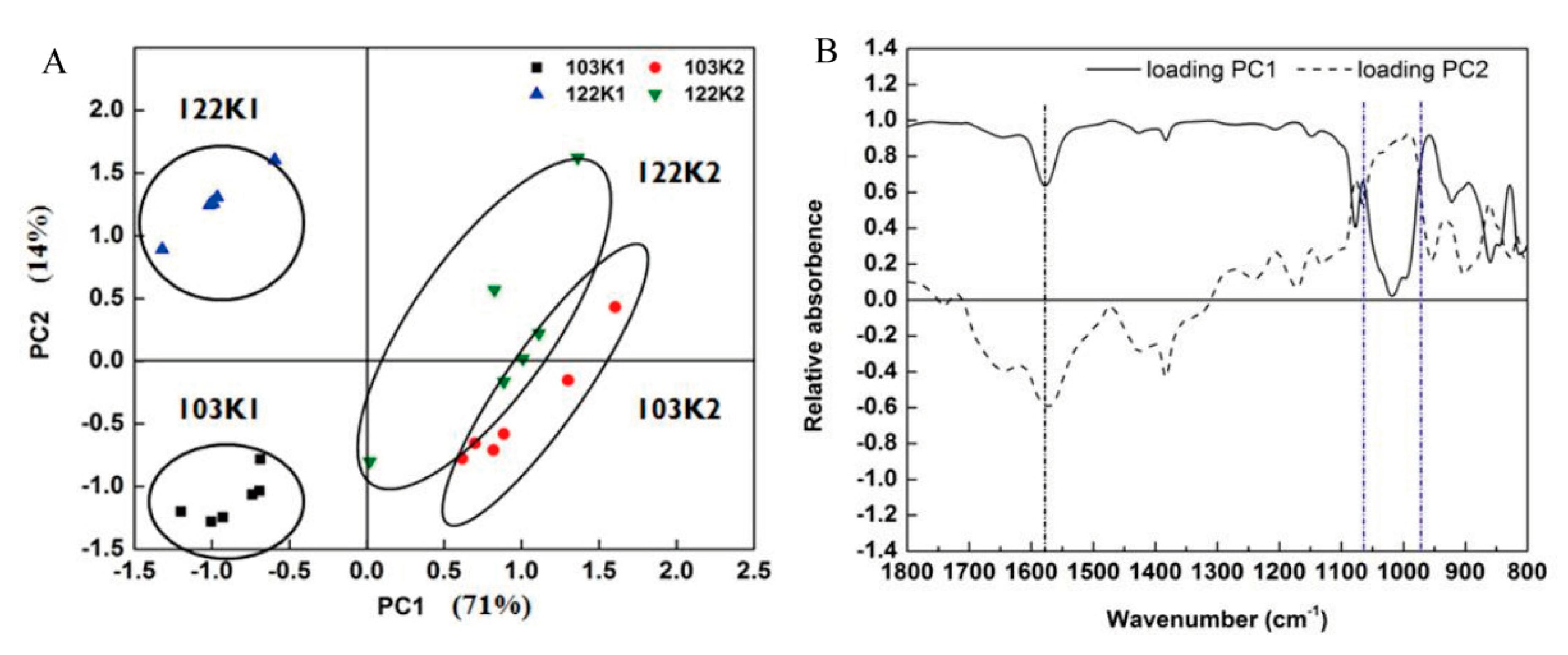
3.2. FTIR Spectral Analysis of the Functional Leaves in the Two Cotton Genotypes
3.3. Semiquantitative Analyses of the Main Absorption Bands in Functional Leaves under Low K
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sullivan, W.M.; Jiang, Z.C.; Hull, R.J. Root morphology and its relationship with nitrate uptake in Kentucky bluegrass. Crop Sci. 2000, 40, 765–772. [Google Scholar] [CrossRef]
- Marschner, P. Mineral Nutrition of Higher Plants, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2012; pp. 299–312. [Google Scholar]
- Szczerba, M.W.; Britto, D.T.; Kronzucker, H.J. K+ transport in plants: Physiology and molecular biology. J. Plant Physiol. 2009, 166, 447–466. [Google Scholar] [CrossRef] [PubMed]
- Alemán, F.; Nieves-Cordones, M.; Martínez, V.; Rubio, F. Root K+ Acquisition in Plants: The Arabidopsis thaliana Model. Plant Cell Physiol. 2011, 52, 1603–1612. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Hua, H.; Eneji, A.E.; Li, Z.; Duan, L.; Tian, X. Genotypic variations in photosynthetic and physiological adjustment to potassium deficiency in cotton (gossypium hirsutum). J. Photochem. Photobiol. B Biol. 2012, 110, 1–8. [Google Scholar] [CrossRef]
- Zörba, C.; Mehmet, S.; Edgar, P. Potassium in agriculture—Status and perspectives. J. Plant Physiol. 2014, 171, 656–669. [Google Scholar] [CrossRef] [PubMed]
- Rengel, Z.; Damon, P.M. Crops and genotypes differ in efficiency of potassium uptake and use. Physiol. Plant. 2008, 133, 624–636. [Google Scholar] [CrossRef]
- Pettersson, S.; Jensén, P. Variation among species and varieties in uptake and utilization of potassium. Plant Soil 1983, 72, 231–237. [Google Scholar] [CrossRef]
- El-Bassam, N. A concept of selection for ‘low input’ wheat varieties. Euphytica 1998, 100, 95–100. [Google Scholar] [CrossRef]
- George, M.S.; Lu, G.Q.; Zhou, W.J. Genotypic variation for potassium uptake and utilization efficiency in sweet potato (Ipomoea batatas L.). Field Crop. Res. 2002, 77, 7–15. [Google Scholar] [CrossRef]
- Yang, X.E.; Liu, J.X.; Wang, W.M.; Li, H.; Luo, A.C.; Ye, Z.Q.; Yang, Y. Genotypic differences and some associated plant traits in potassium internal use efficiency of lowland rice (oryza sativa L.). Nutr. Cycl. Agroecosyst. 2003, 67, 273–282. [Google Scholar] [CrossRef]
- Damon, P.M.; Rengel, Z. Wheat genotypes differ in potassium efficiency under glasshouse and field conditions. Aust. J. Agric. Res. 2007, 58, 816–825. [Google Scholar] [CrossRef]
- Hao, Y.S.; Jiang, C.C.; Wang, X.L.; Xia, Y.; Chen, F. Differences of potassium efficiency characteristics and root morphology between two cotton genotypes. Acta Agron. Sin. 2011, 37, 2094–2098. [Google Scholar] [CrossRef]
- Jiang, C.C.; Gao, X.Z.; Wang, Y.H.; Lu, J.W.; Xu, F.S.; Shi, L. Potassium efficiency of various cotton genotypes and its nutritional mechanisms. Plant Nutr. Fertil. Sci. 2005, 11, 781–786. [Google Scholar]
- Jiang, C.C.; Chen, F.; Gao, X.Z.; Lu, J.W.; Wan, K.N.; Nian, F.Z.; Wang, Y.H. Different responses to potassium stress between potassium high efficiency and potassium low efficiency cotton genotypes efficiency and potassium low efficiency cotton genotypes. Sci. Agric. Sin. 2008, 41, 488–493. [Google Scholar]
- Xia, Y.; Jiang, C.C.; Chen, F.; Lu, J.W.; Wang, Y.H. Differences in growth and potassium−use efficiency of two cotton genotypes. Commun. Soil Sci. Plant Anal. 2011, 42, 132–143. [Google Scholar] [CrossRef]
- Xia, Y.; Jiang, C.C.; Wang, X.; Chen, F. Effects of low potassium stress on the photosynthesis and photosynthate partitioning of cotton. Chin. J. Appl. Ecol. 2013, 32, 1476–1482. [Google Scholar]
- Hao, Y.S.; Lei, J.; Wang, Q.L.; Wu, L.S.; Jiang, C.C. Two typical K−efficiency cotton genotypes differ in potassium absorption kinetic parameters and patterns. Acta Agric. Scand. Sect. B Soil Plant Sci. 2015, 1, 45–53. [Google Scholar] [CrossRef]
- Hao, Y.S.; Lei, J.; Wu, X.W.; Wu, L.S.; Jiang, C.C. Photosynthate transport rather than photosynthesis rate is critical for low potassium adaptation of two cotton genotypes. Acta Agric. Scand. Sect. B Soil Plant Sci. 2016, 66, 170–177. [Google Scholar] [CrossRef]
- Yang, J.; Yen, H.E. Early salt stress effects on the changes in chemical composition in leaves of ice plant and arabidopsis. A fourier transform infrared spectroscopy study. Plant Physiol. 2002, 130, 1032–1042. [Google Scholar] [CrossRef]
- Dokken, K.M.; Davis, L.C.; Marinkovic, N.S. Use of infrared microspectroscopy in plant growth and development. Appl. Spectrosc. Rev. 2005, 40, 301–326. [Google Scholar] [CrossRef]
- Liu, G.D.; Dong, X.C.; Liu, L.C.; Wu, L.S.; Peng, S.A.; Jiang, C.C. Boron deficiency is correlated with changes in cell wall structure that lead to growth defects in the leaves of navel orange seedlings. Sci. Hortic. 2014, 176, 54–62. [Google Scholar] [CrossRef]
- Wu, X.W.; Muhammad, R.; Yan, L.; Du, C.Q.; Liu, L.Y.; Jiang, C.C. Boron deficiency in trifoliate orange induces changes in pectin composition and architecture of components in root cell walls. Front. Plant Sci. 2017, 8, 1882. [Google Scholar] [CrossRef] [PubMed]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Calif. Agric. Exp. Stn. 1950, 347, 357–359. [Google Scholar]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extraets in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Zhang, X.B.; Liu, P.; Li, D.T.; Xu, G.T.; Jiang, M.J. FTIR characterization of chemical composition changes of plant root cell wall induced by chromium. Spectrosc. Spectr. Anal. 2008, 28, 1067–1070. [Google Scholar]
- Kaack, K.; Pedersen, H.L. Effects of potassium, phosphorus and nitrogen fertilization on endogenous ethylene and quality characteristics of apples (Malus Domestica L.). J. Plant Nutr. 2014, 37, 1148–1155. [Google Scholar] [CrossRef]
- Evans, H.J.; Sorger, G.J. Role of mineral elements with emphasis on the univalent cations. Annu. Rev. Plant Physiol. 2003, 17, 47–76. [Google Scholar] [CrossRef]
- Lubin, M.; Ennis, H.L. On the role of intracellular potassium in protein synthesis. BBA Spec. 1964, 80, 614–631. [Google Scholar] [CrossRef]
- Helal, H.M.; Mengel, K. Nitrogen metabolism of young barley plants as affected by NaCl−salinity and potassium. Plant Soil 1979, 51, 457–462. [Google Scholar] [CrossRef]
- Wu, X.W.; Hao, Y.S.; Lei, J.; Jiang, C.C. FTIR spectroscopic characterization of material composition in functional leaf of cotton under potassium or/and boron stress. Spectrosc. Spectr. Anal. 2016, 3, 676–680. [Google Scholar]
- Xie, M.X.; Liu, Y. Determination of protein secondary structure infrared spectrum band for the amide III. Chem. J. Chin. Univ. 2003, 24, 226–231. [Google Scholar]
- Handakumbura, P.P.; Hazen, S.P. Transcriptional regulation of grass secondary cell wall biosynthesis: Playing catch-up with Arabidopsis thaliana. Front. Plant Sci. 2012, 3, 74. [Google Scholar] [CrossRef] [PubMed]
- Reddy, K.R.; Zhao, D. Interactive effects of elevated CO2 and potassium deficiency on photosynthesis, growth, and biomass partitioning of cotton. Field Crop. Res. 2005, 94, 201–213. [Google Scholar] [CrossRef]
- Chérel, I.; Lefoulon, C.; Boeglin, M.; Sentenac, H. Molecular mechanisms involved in plant adaptation to low K+ availability. J. Exp. Bot. 2014, 65, 833–848. [Google Scholar] [CrossRef]
- Xu, Y.W.; Zou, Y.T.; Husaini, A.M.; Zeng, J.W.; Guan, L.L.; Liu, Q.; Wu, W. Optimization of potassium for proper growth and physiological response of Houttuynia cordata Thunb. Environ. Exp. Bot. 2011, 71, 292–297. [Google Scholar] [CrossRef]
- Zhao, D.; Oosterhuis, D.M.; Bednarz, C.W. Influence of potassium deficiency on photosynthesis, chlorophyll content, and chloroplast ultrastructure of cotton plants. Photosynthetica 2001, 39, 103–109. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient Varieties | Contents | |
|---|---|---|
| Macronutrients | NH4NO3 | 0.24 g L−1 |
| Na2HPO4 | 0.10 g L−1 | |
| NaH2PO4 | 0.10 g L−1 | |
| CaCl2·2H2O | 0.36 g L−1 | |
| MgSO4·7H2O | 0.50 g L−1 | |
| Micronutrients | EDTA−Fe | 48.5 mg L−1 |
| H3BO3 | 2.86 mg L−1 | |
| MnCl2·4H2O | 1.81 mg L−1 | |
| ZnSO4·7H2O, | 0.22 mg L−1 | |
| CuSO4·5H2O | 0.08 mg L−1 | |
| Na2MoO4·4H2O | 0.09 mg L−1 |
| Wavenumber (cm−1) | Assignments | Possible Chemicals | Physiological Functions |
|---|---|---|---|
| 1735 | C=O stretching | Ester compound | Cell membrane permeability and cell wall structure |
| 1650 | C=O stretching in peptide bonds | Amide I | Protein structure and content |
| 1550 | N–H bending or C–N stretching | Amide II | |
| 1436 | C–N stretching in peptide bonds | Protein | |
| 1384 | CH3 stretching | Cellulose | Cell wall structure |
| 1244 | C=O stretching or NH2 deformation | Amide III | Protein structure |
| 1103 | C–C, C–O stretching, or C–H bending | Carbohydrates | Photosynthesis |
| 1076 | |||
| 1035 |
| Wavenumber (cm−1) | Treatment | Variation (%) | ||||
|---|---|---|---|---|---|---|
| 103–K1 | 103–K2 | 122–K1 | 122–K2 | 103 | 122 | |
| 1649 | 0.96 | 0.98 | 0.87 | 0.89 | −2.3 | −1.5 |
| 1433 | 0.95 | 1.05 * | 0.86 | 0.93 * | −9.5 | −7.0 |
| 1407 | 0.95 | 1.05 * | 0.86 | 0.93 * | −9.5 | −7.0 |
| 1244 | 0.56 | 0.64 * | 0.58 | 0.63 * | −12.6 | −7.8 |
| 1101 | 1.20 | 1.17 | 1.25 * | 1.10 | 2.9 | 13.1 * |
| 1076 | 1.20 * | 1.12 | 1.30 * | 1.06 | 6.5 | 22.3 * |
| 1037 | 1.02 * | 0.95 | 1.15 * | 0.91 | 7.4 | 27.5 * |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, X.; Hao, Y.; Riaz, M.; Jiang, C. Changes in Leaf Structure and Chemical Compositions Investigated by FTIR Are Correlated with Different Low Potassium Adaptation of Two Cotton Genotypes. Agronomy 2020, 10, 479. https://doi.org/10.3390/agronomy10040479
Wu X, Hao Y, Riaz M, Jiang C. Changes in Leaf Structure and Chemical Compositions Investigated by FTIR Are Correlated with Different Low Potassium Adaptation of Two Cotton Genotypes. Agronomy. 2020; 10(4):479. https://doi.org/10.3390/agronomy10040479
Chicago/Turabian StyleWu, Xiuwen, Yanshu Hao, Muhammad Riaz, and Cuncang Jiang. 2020. "Changes in Leaf Structure and Chemical Compositions Investigated by FTIR Are Correlated with Different Low Potassium Adaptation of Two Cotton Genotypes" Agronomy 10, no. 4: 479. https://doi.org/10.3390/agronomy10040479
APA StyleWu, X., Hao, Y., Riaz, M., & Jiang, C. (2020). Changes in Leaf Structure and Chemical Compositions Investigated by FTIR Are Correlated with Different Low Potassium Adaptation of Two Cotton Genotypes. Agronomy, 10(4), 479. https://doi.org/10.3390/agronomy10040479