Optimizing Nitrogen Fertigation Rates for Young Southern Highbush Blueberry


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Site
2.2. Experimental Design and Treatments
2.3. Plant Growth Measurement
2.4. Soil Sample Collection and Analysis
2.5. Fruit Yield and Quality Measurements
2.6. Data Analysis
3. Results and Discussion
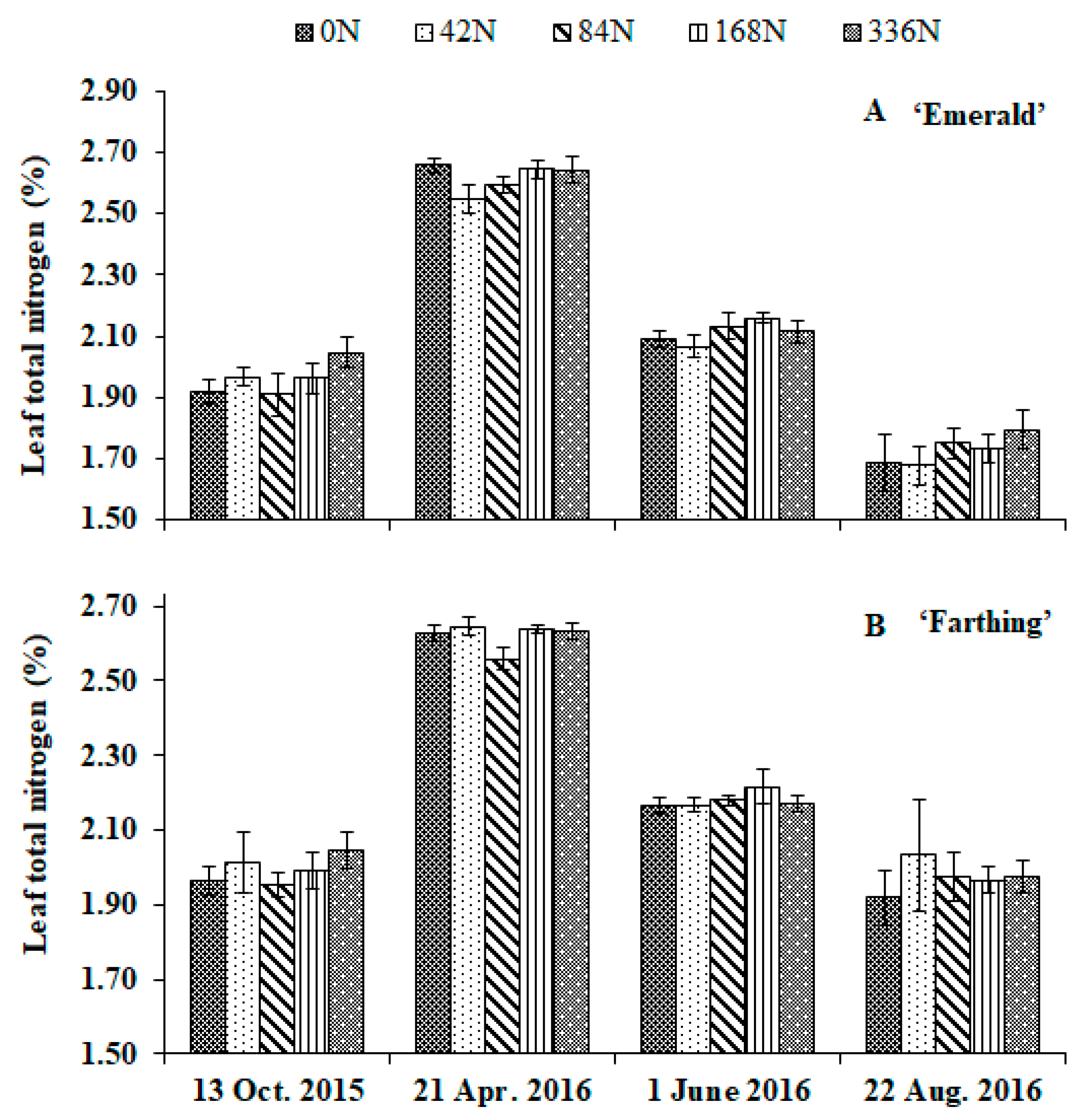
3.1. Leaf Nutrient Concentration
3.2. Soil Nitrate Concentration
3.3. Plant Growth
3.4. Bloom Time
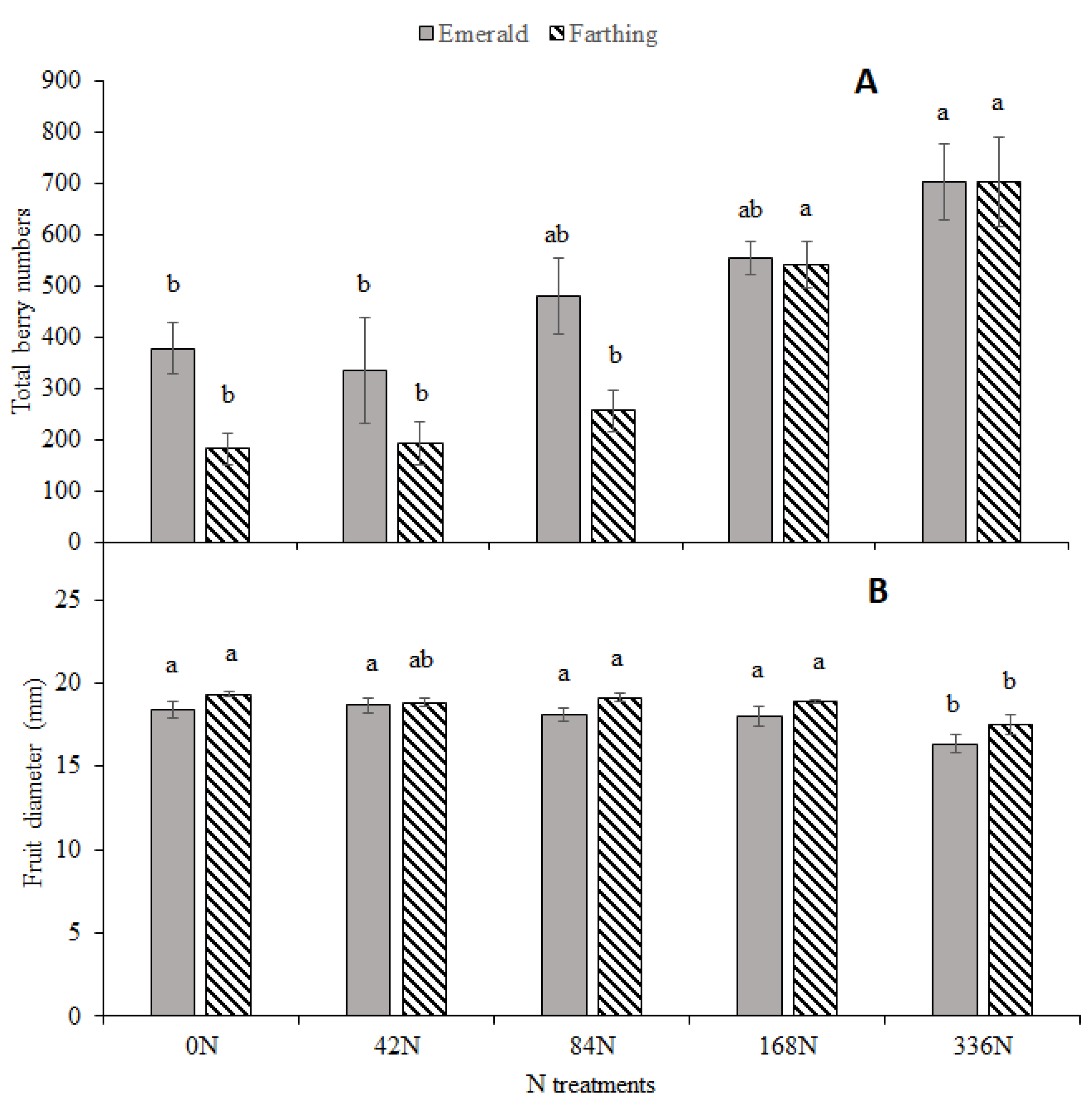
3.5. Fruit Yield, Berry Number, Berry Size
3.6. Fruit Quality
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- National Agriculture Statistics and Services (NASS); United Stated Department of Agriculture (USDA). Noncitrus Fruits and Nuts 2018 Summary. Agricultural Statistics Board. 2019. Available online: https://www.nass.usda.gov/Publications/Todays_Reports/reports/ncit0619.pdf (accessed on 18 January 2020).
- Williamson, J.G.; Miller, E.P. Effects of fertilizer rate and form on vegetative growth and yield of southern highbush blueberry in pine bark culture. HortTechnology 2009, 19, 152–157. [Google Scholar] [CrossRef]
- Banados, M.P. Dry Weight and 15N-Nitrogen and Partitioning, Growth, and Development of Young and Mature Blueberry Plants. Ph.D. Thesis, Oregon State University, Corvallis, OR, USA, 2006. Available online: http://ir.library.oregonstate.edu/xmlui/bitstream/handle/1957/9649/Banados_Maria_P_2006.pdf?sequence=1&origin=publication_detail (accessed on 19 December 2019).
- Fang, Y.; Williamson, J.; Darnell, R.; Li, Y.; Liu, G. Nitrogen Uptake and Allocation at Different Growth Stages of Young Southern Highbush Blueberry Plants. HortScience 2017, 52, 905–909. [Google Scholar] [CrossRef]
- Banados, M.P.; Strik, B.C.; Bryla, D.R.; Righetti, T.L. Response of highbush blueberry to nitrogen fertilizer during field establishment, I: Accumulation and allocation of fertilizer nitrogen and biomass. HortScience 2012, 47, 648–655. [Google Scholar] [CrossRef]
- White, L.D. The Effect of Pre-Plant Incorporation with Sawdust, Sawdust Mulch, and Nitrogen Fertilizer Rate on Soil Properties and Nitrogen Uptake and Growth of ‘Elliott’ Highbush Blueberry. Master’s Thesis, Oregon State University, Corvallis, OR, USA, 2006. Available online: http://ir.library.oregonstate.edu/xmlui/bitstream/handle/1957/1363/WholeThesis.pdf?sequence=1 (accessed on 19 December 2019).
- Bryla, D.R.; Machado, R.M. Comparative effects of nitrogen fertigation and granular fertilizer application on growth and availability of soil nitrogen during establishment of highbush blueberry. Front. Plant Sci. 2011, 2, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bryla, D.R.; Strik, B.C. Nutrient requirements, leaf tissue standards, and new options for fertigation of northern highbush blueberry. HortTechnology 2015, 25, 464–470. [Google Scholar] [CrossRef]
- Hanson, E.J. Nitrogen fertilization of highbush blueberry. In Proceedings of the VIII International Symposium on Vaccinium Culture, ISHS Acta Horticulturae 715, 347–352, Sevilla, Spain, 31 August 2006. [Google Scholar]
- Thomas, B.P.; Law, L.; Stankey, D.L. Soil survey of Marion County Area, Florida. USDA/Natural Resources Conservation Service. 1979. Available online: https://www.nrcs.usda.gov/Internet/FSE_MANUSCRIPTS/florida/FL608/0/Marion.pdf (accessed on 10 March 2020).
- Krewer, G.W.; Ruter, J.M. Fertilizing Highbush Blueberries in Pine Bark Beds. 2009. Available online: http://athenaeum.libs.uga.edu/bitstream/handle/10724/12290/B1291.pdf?sequence=1 (accessed on 19 December 2019).
- Campillo, C.; Prieto, M.H.; Daza, C.; Monino, M.J.; Garcia, M.I. Using digital images to characterize canopy coverage and light interception in a processing tomato crop. Hortscience 2008, 43, 1780–1786. [Google Scholar] [CrossRef]
- RStudio Team. RStudio: Integrated Development for R; RStudio, Inc.: Boston, MA, USA, 2016; Available online: http://www.rstudio.com/ (accessed on 19 December 2019).
- Hochmuth, G.; Hanlon, E.; Overman, A. Fertilizer Experimentation, Data Analyses, and Interpretation for Developing Fertilization Recommendations-Examples with Vegetable Crop Research; University of Florida Institution of Food and Agriculture Science: Gainesville, FL, USA, 2011; Available online: https://edis.ifas.ufl.edu/ss548 (accessed on 19 December 2019).
- Mills, A.H.; Jones, J.B. Plant Analysis Handbook II: A Practical Sampling, Preparation, Analysis and Interpretation Guide; Micro-Macro: Athens, GA, USA, 1996. [Google Scholar]
- Ballinger, W.E.; Kenworthy, A.L.; Bell, H.K.; Benne, E.J.; Bass, S.T. Relationship between nutrient element content of blueberry foliage and fruit. Mich. State Univ. Q. Bull. 1958, 40, 906–911. [Google Scholar]
- Bryla, D.R.; Strik, B.C.; Banados, M.P.; Righetti, T.L. Response of highbush blueberry to nitrogen fertilizer during field establishment—II. Plant nutrient requirements in relation to nitrogen fertilizer supply. HortScience 2012, 47, 917–926. [Google Scholar] [CrossRef]
- Cummings, G.A. Plant and soil effects of fertilizer and lime applied to highbush blueberries. J. Am. Soc. Hortic. Sci. 1987, 103, 302–305. [Google Scholar]
- Cummings, G.; Bickford, C.; Nelson, L. Fertilizer and lime rates influence highbush blueberry growth and foliar elemental content during establishment. J. Am. Soc. Hortic. Sci. 1971, 96, 184–186. [Google Scholar]
- Nario, A.; Pino, I.; Zapata, F.; Albornoz, M.P.; Baherle, P. Nitrogen (15N) fertilizer use efficiency in peach (Prunus persica L.) cv. Goldencrest trees in Chile. Sci. Hortic. 2003, 97, 279–287. [Google Scholar] [CrossRef]
- Tagliavini, M.; Quartieri, M.; Millard, P. Remobilised nitrogen and root uptake of nitrate for spring leaf growth, flowers and developing fruits of pear (Pyrus communis L.) trees. Plant Soil 1997, 195, 137–142. [Google Scholar] [CrossRef]
- Tagliavini, M.; Millard, P.; Quartieri, M. Storage of foliar-absorbed nitrogen and remobilization for spring growth in young nectarine (Prunus persica var. nectarina) trees. Tree Physiol. 1998, 18, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Birkhold, K.T.; Darnell, R.L. Contribution of storage and currently assimilated nitrogen to vegetative and reproductive growth of rabbiteye blueberry. J. Am. Soc. Hortic. Sci. 1993, 118, 101–108. [Google Scholar] [CrossRef]
- George, A.P.; Nissen, R.J. Effects of water stress, nitrogen and paclobutrazol on flowering, yield and fruit quality of the low-chill peach cultivar, ‘Flordaprince’. Sci. Hortic. 1992, 49, 197–209. [Google Scholar] [CrossRef]
- Wilber, W.L.; Williamson, J.G. Effects of fertilizer rate on growth and fruiting of containerized southern highbush blueberry. HortScience 2008, 43, 143–145. [Google Scholar] [CrossRef]
- Ehret, D.L.; Frey, B.; Forge, T.; Helmer, T.; Bryla, D.R.; Zebarth, B.J. Effects of nitrogen rate and application method on early production and fruit quality in highbush blueberry. Can. J. Plant Sci. 2014, 94, 1165–1179. [Google Scholar] [CrossRef]
- Kozinski, B. Influence of mulching and nitrogen fertilization rate on growth and yield of highbush blueberry. In Proceedings of the VIII International Symposium on Vaccinium Culture; Acta Horticulturae; ISHS: Leuven, Belgium, 2006; Volume 715, pp. 231–236. [Google Scholar]
- Maust, B.E.; Williamson, J.G.; Darnell, R.L. Flower bud density affects vegetative and fruit development in field-grown southern highbush blueberry. HortScience 1999, 34, 607–610. [Google Scholar] [CrossRef]
- Casamali, B.; Darnell, R.L.; Kovaleski, A.P.; Olmstead, J.W.; Williamson, J.G. Vegetative and Reproductive Traits of Two Southern Highbush Blueberry Cultivars Grafted onto Vaccinium arboreum Rootstocks. HortScience 2016, 51, 880–886. [Google Scholar] [CrossRef]
- Ourecky, D.K.; Bourne, M.C. Measurement of strawberry texture with an Instron machine. Proc. Am. Soc. Hort. Sci. 1968, 93, 317–325. [Google Scholar]
- Ehlenfeldt, M.K.; Martin, R.B. A survey of fruit firmness in highbush blueberry and species-introgressed blueberry cultivars. HortScience 2002, 37, 386–389. [Google Scholar] [CrossRef]
- Skupień, K. Chemical composition of selected cultivars of highbush blueberry fruit (Vaccinium corymbosum L.). Folia Hortic. 2006, 18, 47–56. [Google Scholar]
- Beckles, D.M. Factors affecting the postharvest soluble solids and sugar content of tomato (Solanum lycopersicum L.) fruit. Postharvest Biol. Technol. 2012, 63, 129–140. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Nitrate (mg kg−1) | ||||
|---|---|---|---|---|
| N (kg/ha/year) | 23 March | 17 May | 18 July | 15 September |
| 15 cm z | ||||
| 0 | 1.19 b y | 2.19 c | 3.10 | 1.86 b |
| 42 | 1.94 b | 2.05 c | 4.58 | 0.81 b |
| 84 | 1.64 b | 1.92 c | 2.84 | 2.58 b |
| 168 | 3.30 b | 8.31 b | 7.96 | 7.99 b |
| 336 | 6.60 a | 13.62 a | 2.79 | 34.06 a |
| Significance x | L *** | L *** | NS | L *** |
| r-value w | 0.85 | 0.92 | 0.88 | |
| 30 cm | ||||
| 0 | 1.23 b | 2.30 b | 1.59 | 1.09 b |
| 42 | 1.70 b | 2.64 b | 4.01 | 0.40 b |
| 84 | 1.12 b | 2.89 b | 1.77 | 1.97 b |
| 168 | 2.17 b | 7.28 b | 6.62 | 4.49 b |
| 336 | 5.40 a | 17.51 a | 2.08 | 23.63 a |
| Significance | L *** | L *** | NS | L *** |
| r-value | 0.79 | 0.88 | 0.80 | |
| 76 cm | ||||
| 0 | 0.75 b | 1.91 b | 1.22 | 0.40 b |
| 42 | 0.73 b | 2.41 b | 1.51 | 1.55 ab |
| 84 | 0.85 b | 1.92 b | 3.87 | 1.26 ab |
| 168 | 1.43 b | 2.64 b | 5.79 | 3.86 a |
| 336 | 4.31 a | 7.49 a | 0.85 | 3.72 ab |
| Significance | L *** | L *** | NS | L ** |
| r-value | 0.81 | 0.76 | 0.59 | |
| N (kg/ha/yr) | ‘Emerald’ Ground Cover (%) | ‘Farthing’ Ground Cover (%) | ||
|---|---|---|---|---|
| 2015 | 2016 | 2015 | 2016 | |
| 0 | 14.38 b z | 57.25 b | 9.03 b | 60.39 b |
| 42 | 11.91 b | 58.88 b | 8.97 b | 60.82 b |
| 84 | 13.90 b | 70.06 b | 8.00 b | 77.21 ab |
| 168 | 19.65 ab | 70.42 b | 16.44 a | 83.36 ab |
| 336 | 23.13 a | 93.85 a | 17.36 a | 87.1 a |
| Significance y | L *** | L *** | L *** | L *** |
| R value x | 0.69 | 0.76 | 0.75 | 0.63 |
| Weeks z | ||
|---|---|---|
| N (kg/ha/yr) | Emerald | Farthing |
| 0 | 8.70 a y | 10.50 a |
| 42 | 8.50 a | 10.20 ab |
| 84 | 7.20 ab | 10.50 a |
| 168 | 5.60 bc | 9.60 bc |
| 336 | 4.20 c | 9.00 c |
| Significance x | L *** | L *** |
| R value w | −0.89 | −0.8 |
| Cumulative Percentage of Yield (%) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| N (kg/ha/yr) | Week 16 z | Week 17 | Week 18 | Week 19 | Week 20 | Week 21 | Week 22 | Week 23 | Week 24 | Week 25 |
| ‘Emerald’ | ||||||||||
| 0 | - | 0.40 b y | 5.04 c | 23.80 b | 51.15 b | 80.07 b | 93.86 | 100 | - | - |
| 42 | - | 0.62 b | 8.16 c | 30.02 b | 59.60 b | 83.99 ab | 92.91 | 100 | - | - |
| 84 | - | 1.85 b | 13.37 c | 39.21 b | 67.45 ab | 87.42 ab | 95.25 | 100 | - | - |
| 168 | - | 5.04 b | 31.43 b | 60.17 a | 82.84 a | 95.47 a | 97.62 | 100 | - | - |
| 336 | - | 21.53 a | 49.49 a | 72.09 a | 87.15 a | 95.57 a | 98.15 | 100 | - | - |
| ‘Farthing’ | ||||||||||
| 0 | 1.01 b | 1.84 b | 6.02 b | 15.95 b | 40.39 c | 68.85 c | 86.67 b | 93.37 b | 96.36 ab | 100 |
| 42 | 1.56 b | 2.21 b | 6.89 b | 17.04 b | 42.81 c | 71.75 bc | 85.67 b | 92.85 b | 95.62 b | 100 |
| 84 | 0.91 b | 1.96 b | 8.77 b | 21.25 b | 47.76 bc | 73.38 bc | 89.29 b | 94.59 ab | 97.60 ab | 100 |
| 168 | 2.79 ab | 4.44 ab | 13.72 b | 25.62 ab | 55.29 ab | 80.39 ab | 92.83 a | 96.82 ab | 98.31 ab | 100 |
| 336 | 4.98 a | 8.21 a | 24.28 a | 36.64 a | 63.80 a | 83.75 a | 94.28 a | 97.76 a | 98.84 a | 100 |
| Fruit Firmness (g/mm) | Total Soluble Solids to Titratable Acidity Ratio | |||||
|---|---|---|---|---|---|---|
| N (kg/ha/yr) | 2-May | 9-May | 16-May | 9-May | 16-May | 23-May |
| 0N | 254.0 b | 239.2 b | 229.2 b | 11.51ab z | 14.11 | 15.63 bc |
| 42N | 263.3 ab | 247.6 b | 231.5 ab | 10.33 b | 11.95 | 14.70 c |
| 84N | 251.4 b | 239.4 b | 225.0 b | 12.11 ab | 14.52 | 16.10 bc |
| 168N | 253.4 b | 239.0 b | 219.4 b | 13.97 a | 14.64 | 19.71 ab |
| 336N | 272.2 a | 271.2 a | 248.5 a | 12.00 ab | 12.84 | 21.98 a |
| Significance y | L * | L *** | L * | Q * | NS | L *** |
| R value x | 0.4 | 0.65 | 0.4 | −0.53 | 0.8 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, Y.; Williamson, J.; Darnell, R.; Li, Y.; Liu, G. Optimizing Nitrogen Fertigation Rates for Young Southern Highbush Blueberry. Agronomy 2020, 10, 389. https://doi.org/10.3390/agronomy10030389
Fang Y, Williamson J, Darnell R, Li Y, Liu G. Optimizing Nitrogen Fertigation Rates for Young Southern Highbush Blueberry. Agronomy. 2020; 10(3):389. https://doi.org/10.3390/agronomy10030389
Chicago/Turabian StyleFang, Yang, Jeffrey Williamson, Rebecca Darnell, Yuncong Li, and Guodong Liu. 2020. "Optimizing Nitrogen Fertigation Rates for Young Southern Highbush Blueberry" Agronomy 10, no. 3: 389. https://doi.org/10.3390/agronomy10030389
APA StyleFang, Y., Williamson, J., Darnell, R., Li, Y., & Liu, G. (2020). Optimizing Nitrogen Fertigation Rates for Young Southern Highbush Blueberry. Agronomy, 10(3), 389. https://doi.org/10.3390/agronomy10030389