1. Introduction
Soybean has the highest average area harvested among all legumes [
1]. Soybean is reported to be sensitive to several abiotic stresses as compared to other legumes and crops [
2,
3,
4]. A major abiotic stress, affecting soybean production and quality is drought stress [
5,
6], and with the fact that the intensity of drought stress conditions was increasingly noticed and recorded among the past few decades [
7], understanding the influence of drought stress on crops became vital, as such understanding can be exploited in irrigation-scheduling which, in part reduces drought-related fluctuations in food production [
8].
Crop-drought susceptibility differs from a stage to another during the crop’s life cycle [
9,
10]. This conclusion was reported on soybean [
11,
12]. Moreover, it was reported that soybean plants have low water demands at vegetative stages, whereas these demands increase during reproductive stages [
13]. As such, early drought, during vegetative stages, might not affect soybean final seed yield [
14,
15,
16]; moreover, it was reported that even at early reproductive stages (particularly flowering stages) it did not measurably affect yield, whereas drought during pod filling stages significantly decreased the yield [
17]. However, Lozovaya et al. [
18] reported that relative drought stress during seed formation (R6) stage enhanced the seeds’ quality by increasing many isoflavones; however, seed formation was affected. Yield reduction is correlated with both flower and pod abortion as a consequence of drought conditions [
6]. Moreover, soybean plants subjected to drought at vegetative stages had less leaf area, less photosynthesis rate, and less biomass; however, it was possible for the stressed plants to partially recover during post-stress period. On the other hand, drought occurred later during reproductive stages resulted in higher flower-abortion rate and reduced developed pods and seeds, with less opportunity to recover, resulting in noticeable yield loss [
19,
20].
Desclaux and Roumet [
21] reported that soybean morphology was significantly influenced by drought, especially during reproductive stages. For example, Freitas et al. [
22] reported that plant height of soybean plants was affected by drought. Both yield and seed quality of soybean are significantly influenced by drought stress conditions [
23]. In addition, chlorophyll production was reported to be influenced by drought stress [
24].
Not only the stage, but also both the duration of the drought occurrence and the genotype have a role in the damage level [
25,
26]. For example, it was reported that drought stress has different effects on different soybean cultivars in the study area, as yield, protein, and oil concentrations of certain cultivars were measurably reduced by drought but were increased in other cultivars [
27]. The authors attributed these different results to the different potentials of each cultivar for coping with drought stress, in addition to the timing when plants suffered from stress.
Nitrogen (N) is one of the most important macronutrients for plant growth and yield; it is essential for total chlorophyll content and protein synthesis. Because of its high protein concentration in the seeds (40%, on average), soybean plants have high N requirements [
28]. In addition, N is essentially needed for soybean’s vegetative growth in order to produce optimum biomass [
29]. N deficiency causes N from leaves to be remobilized to the seeds, which in part will lead to decreased photosynthesis and eventually reduced yield [
30]. Wani et al. [
31] reported that relatively small N rates resulted in significant responses from grain legumes. N fertilizer was reported to especially be important under abiotic stresses [
32]. However, the prices of N-fertilizers were increased as a result of the increased cost of the energy required for manufacturing. In addition, nitrous oxide, a greenhouse gas that contaminates water with nitrate, is formed from N-fertilizers [
33]. Therefore, minimizing chemical N-fertilizers seems to be crucial, especially with the fact that N
2-fixed nitrogen, in the case of legumes, is a cheaper source with no similar environmental effects.
As a newly entered crop in the study area, no published work on the sole response of soybean plants to different levels of drought stress and to different rates of N-fertilizer on the physio-morphological level, nor on the quantitative and/or qualitative traits was found. This research aims at evaluating the sole and combined influence of drought stress and nitrogen-fertilizer application on soybean (Glycine max (L.) Merrill. cv. Pannonia Kincse).
2. Materials and Methods
A field experiment was carried out in the experimental station of the University of Debrecen (Látókép) (N. latitude 47°33′, E. longitude 21°27′) on soybean (cv. Pannonia Kincse, Bonefarm, Hungary) during 2017, 2018, and 2019 growing seasons. Soil type of the site is calcareous chernozem. Before sowing in 2017, soil samples were collected from two depths; 0–25 cm and 25–50 cm, and the following data of soil components were provided by the chemical analysis (averaged over the two depths); pH: 6.41, OM: 2.46%, N: 3.97 ppm, P2O5: 90.7 ppm, K2O: 206.7 ppm. Sowing dates were 23 April in 2017 and 26 April in 2018 and 2019, and harvest dates were 15 September in 2017 and 16 September in 2018 and 2019.
The experimental design was split-plot design, with three irrigation regimes; non-irrigated, half-irrigated, and fully irrigated (NI, HI, and FI, respectively) being the main plots, and three N-fertilizer (NH
4NO
3) rates (applied with sowing as a single application); 0, 35, and 105 kg ha
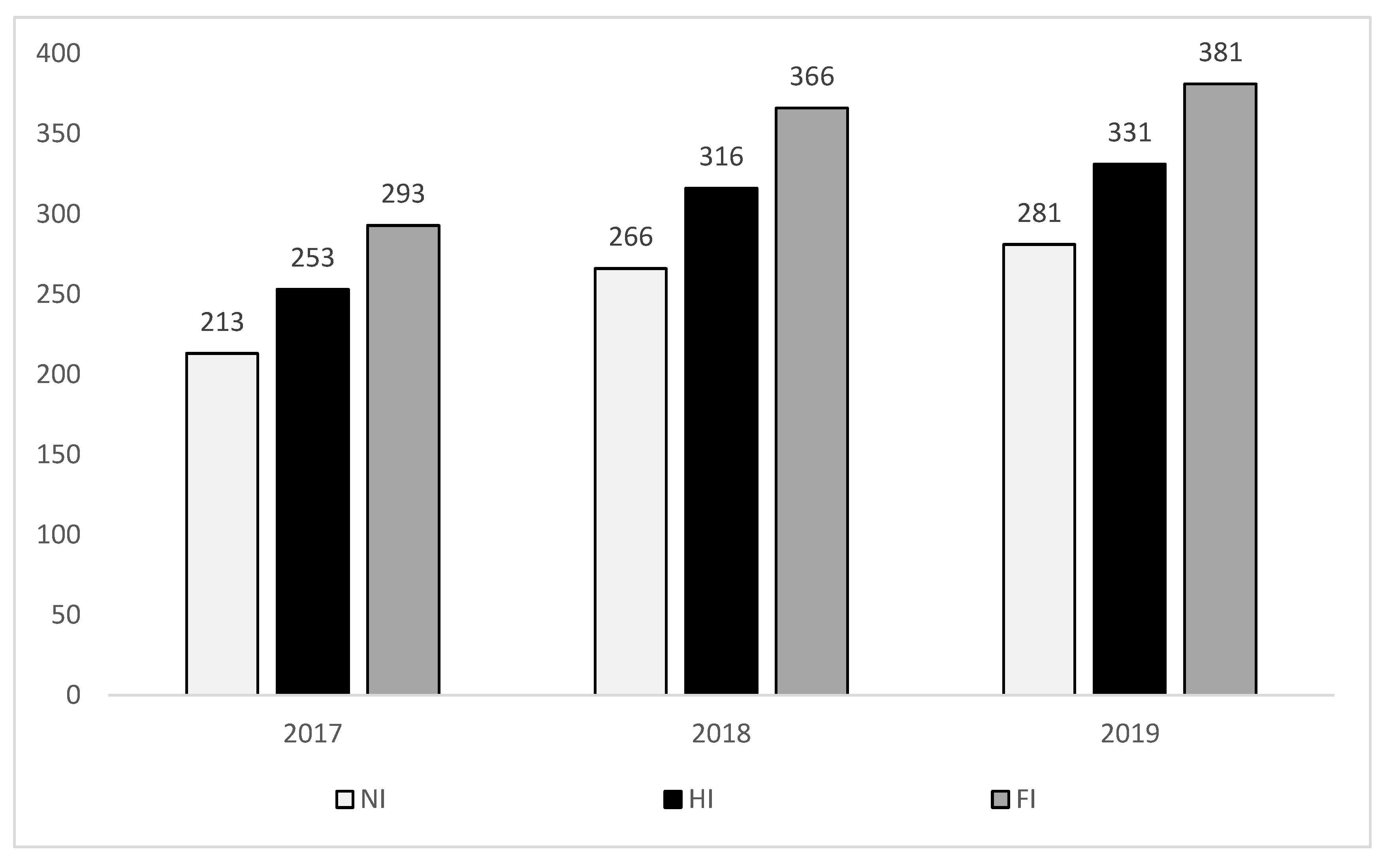
−1 N (0N, 35N and 105N, respectively) being the sub-plots with four replicates each. NI treatment received only precipitation as water irrigation amount, whereas HI treatment received, in addition to precipitation, a total of 40 mm of irrigation water in 2017 and 50 mm in 2018 and 2019. FI treatment, on the other hand, received, in addition to precipitation, a total of 80 mm of irrigation water in 2017 and 100 mm in 2018 and 2019 (
Figure 1). Based on the daily monitoring of precipitation and temperature, field capacity (FC) was regularly tested in all plots, and irrigation was applied in HI and FI treatments whenever soil water content reached 65–70% FC. FC of NI treatment reached a minimum value of 30% in 2018 and 35% in 2017 and 2019.
Final plot number was 36 (3 irrigation regime × 3 fertilization rates × 4 replications). The plot area was 49.68 m² with 12 rows in each plot.
LAI (leaf area index) values were recorded using SS1–SunScan canopy analysis system (Delta-T Devices, Cambridge, UK). Relative chlorophyll content (in the form of SPAD) (soil plant analysis development) was measured using SPAD-502Plus (Konica Minolta, Tokyo, Japan). NDVI (Normalized Difference Vegetation Index) values were recorded using Trimble Greenseeker Handheld (AS Communications Ltd., Cambridgeshire, UK). Ten randomly selected plants from the middle rows of each plot were used for the mentioned traits, and three measurements from the second most developed trifoliate (one measurement for each leaflet) were taken and then averaged. All traits were measured at four different stages of soybean’s life cycle [
34]; fourth node (V4), full bloom (R2), full pod (R4), and full seed (R6).
Flower number per plant was counted at R2 stage. Pod number per plant was counted at R4 stage. Plant height was measured at R6 stage using a standard ruler. Ten randomly selected plants from the middle rows of each plot were used for the mentioned traits.
Seed yield was calculated by harvesting the middle 4 m of each plot and adjusting the yield to 13% moisture content. Hundred-seed weight was calculated after oven-drying the seeds at 65 °C until constant weight. Both protein and oil concentrations were determined using NIR analyser Granolyser (Pfeuffer, Kitzingen, Germany).
SPSS software was run to analyze and compare the means (ANOVA) and to indicate the effect size (by means of Partial Eta Squared), followed by Tukey post-hoc test to indicate the statistically different means, and Pearson’s correlation to indicate correlation coefficient (IBM SPSS ver.26, USA software). All data presented and analyzed are means of the three years of experiment.
4. Discussion
Our results showed that higher SPAD values at both vegetative (V4) and early reproductive (R2 and R4) stages were recorded under drought stress conditions. Gavili et al. [
35] reported that both moderate and severe drought conditions significantly increased soybean SPAD values by 11% and 20%, respectively. The authors justified this increase by the increased N concentrations caused by the decreased fresh or dry matter, and the enhanced N concentration will, in turn, enhance the chlorophyll content. Bredemeier [
36] reported similar conclusion on maize. In their experiment, Fenta et al. [
37] subjected three soybean cultivars to drought stress by withholding irrigation at V3 stage for 28 days (the plants, however, received precipitation amount of 34 mm during this period). The authors reported that SPAD values of the three cultivars were reduced by drought. Drought reduced chlorophyll concentration by 7% in soybean plants [
38]. SPAD values enhanced with applied fertilization. [
39] concluded that the application of 200 kg ha
−1 of N insignificantly enhanced chlorophyll content (by 1.8% under non-stressed conditions and by 3% when drought stress occurred at R2 stage). Increasing N rate resulted in better SPAD values at different stages in soybean [
40]. Similar conclusion was reported by Shafagh-Kolvanagh et al. [
41].
We found out that drought decreased LAI values at all studied reproductive (R2, R4, and R6) stages, especially at R4 stage where the reduction was significant. Similarly, Gavili et al. [
35] reported that moderate and severe drought (corresponding to 70% and 55% FC, respectively) significantly decreased plant leaf area by 29% and 35% at V10 stage, 23% and 31% at V3 stage, and 26% and 36% at R6 stage. Karam et al. [
42] concluded that LAI decreased by 52% under drought stress conditions imposed at R2 stage. Pagter et al. [
43] explained the decreased LAI under drought stress conditions to be the result of less newly produced leaves with a smaller size and a higher falling rate. Severe drought stress imposed at R4 stage resulted in 61.4% less leaf area in soybean [
8]. Moosavi et al. [
44] reported decreased leaf area in canola plants as a result of drought stress application. Çakir [
45] also reported a 23.5% decrease in leaf area when maize plants were subjected to drought stress conditions during the tasseling period. High rate of N significantly increased LAI value at all stages except the late reproductive stage R6 where the LAI values were very close in all fertilization treatments. Caliskan et al. [
32] concluded that soybean LAI linearly increased with increased N rates. DeMooy et al. [
46] and Watanabe et al. [
47] reported that adding N fertilizer before reproductive stages enhances growth and LAI, consequently flowering and yield.
Our results showed that drought enhanced NDVI values at early stages, but reduced them at later stages. Crusiol et al. [
48] concluded that NDVI was higher in two soybean cultivars under early drought conditions, but not under late drought. Drought significantly reduced NDVI as reported by Camoglu et al. [
49]. Fertilization insignificantly enhanced NDVI values. Similar conclusion was reported on wheat [
50] and maize [
51].
Plant height was, on average, reduced by 4.0% and 5.3% under drought stress conditions compared to half- and fully irrigated regimes, respectively. Both Newark [
52] and El Kheir et al. [
53] reported decreased plant height under drought stress conditions. Gavili et al. [
35] reported a 33% and 60% plant height reduction in their experiment under 70% and 55% FC conditions, respectively. Soybean plants had 22.4% shorter plants when severe drought stress occurred at R4 stage, whereas only 9% reduction was reported when same severe drought occurred at R6 stage [
8]. Drought reduced soybean plant height by 31.1% [
22]. Neilson and Nelson [
54] explained this reduction in plant height under drought by the delayed stem elongation caused by shortened distance among nodes. An indeterminate soybean cultivar (OAC Bayfield) was put under two drought stress severities; W1 and W2 (corresponding to 25% and 50% of crop evapotranspiration (ETc), respectively as compared to control, 100% ETc) at R1 stage [
55]. Plant height decreased by 33% and 28% in W1 and W2 treatments, respectively after 9 days of stress application. Furthermore, 16 days of drought imposition resulted in 56% and 47% reduction in plant height in W1 and W2 treatments, respectively. Plant height was relatively enhanced by fertilization. Similar conclusion was reported earlier by Hanway and Weber [
56] and Dadson and Acquaah [
57].
In our experiment, drought (non-irrigated regime) significantly decreased flower number per plant in all fertilization treatments compared to half-irrigated counterparts. He et al. [
58] reported that cyclic drought (where water was withheld from V5–V6 stage until reaching 30% field capacity (FC), and then plants were re-watered to 100% FC, and again water was withheld in order to reach 30% FC) resulted in 53.8% decreased flower number per plant, whereas terminal drought (where no irrigation was applied after V5–V6 stage) further increased that ratio to 72.5%. In their experiment, Atti et al. [
55] reported that flower number per plant decreased by 79.4% and 58.8% in W1 and W2 treatments, respectively. The authors explained this decrease by both reduced node number and increased flower abortion as a result of drought stress application. Flower number in chickpea was reported to have an important role on the final yield [
59]. Flower number per plant was enhanced by fertilization. Similarly, Purcell and King [
60] reported that applying N fertilizer increased flower number in plants by reducing flower abortion rate. Brevedan et al. [
61] reported similar conclusion under both greenhouse and field conditions.
Both irrigation regimes resulted in enhanced pod number per plant compared to drought-stressed counterpart, which was similarly reported by Pookpakdi et al. [
62] and Pawar et al. [
63] and later by He et al. [
58]; the authors concluded that cyclic and terminal drought stress resulted in 42.3% and 90.4% less pods per plant. Westgate and Peterson [
64] concluded that drought stress during flowering caused a 70% reduction in pod number per plant. Exposing soybean plants to drought at pod filling stages decreased pod number per plant by 36.6%, whereas a 42.6% reduction was recorded when drought was imposed at flowering stage [
65]. Pod number decreased from 25 to 15 when available water decreased from 100 to 70% FC, and further reduction to 55% FC further decreased pod number to 14 [
35]. In their experiment, Iqbal et al. [
66] decreased FC from 100 to 50% at R4 stage to study the effect of drought at this stage on soybean; they reported that pod number per plant significantly decreased by 21.4% as a consequence of drought imposition, and when FC was further reduced to 20%, another significant reduction (by 34.7% compared to 100% FC treatment) was recorded in this trait. Leport et al. [
67] concluded that decreased pod number majorly affects the seed yield of chickpea.
Drought-stressed treatment reduced final seed yield relative to both irrigated treatments. Previously, Foroud et al. [
14] and Fenta et al. [
37] reported that drought decreased the seed yield of soybean plants. Reductions in seed yield under drought stress conditions were also reported by Liu et al. [
6] and Masoumi et al. [
68]. Moreover, seed yield was reduced by 57.4% and 95.3% as a result of cyclic and terminal drought stress, respectively [
58]. Moderate drought at R4 stage reduced soybean seed yield by 31.2% (averaged on both years of experiment), whereas severe drought at the same stage resulted in 77.7% less seed yield [
8]. The same researchers also reported that subjecting soybean plants to moderate and severe drought at R6 stage decreased the final seed yield by 33.4% and 62.4%, respectively. When drought was imposed at R4 stage, soybean plants had 32.0% and 48.7% less seed yield under 50% and 20% FC, respectively compared to 100 FC control [
66]. The authors concluded that the decrease in seed yield was mainly caused by increased number of empty pods, decreased number of seeds per plant, decreased 100-seed weight, and decreased number of pods per plant. Jumrani and Bhatia [
69] subjected soybean plants to drought stress at two different stages; V4 and R5. They reported that the seed yield was decreased by 28% and 74%, respectively compared to control treatment where no drought stress was imposed, concluding that drought had much higher effect when it was imposed at reproductive stage R5 as compared to vegetative stage V4. Fertilization increased the yield under all irrigation regimes except when high rate was applied with the absence of drought, so a conclusion that low-rate fertilization is recommended under all irrigation regimes, whereas high rates of N are only recommended under relative drought conditions can be drawn. A 10% increase in seed yield was recorded with the application of 200 kg ha
−1 N under drought stress imposed at R2 stage, whereas the same application decreased the yield by 1.5% under drought-free conditions [
39]. Hungria et al. [
70] also reported that the application of 200 kg ha
−1 of N-fertilizer did not increase the yield. Other reports also concluded that N-fertilizer application resulted in better seed yield under drought stress conditions [
30,
71]. The application of N fertilizer increased soybean drought tolerance as it enhanced the accumulation of both shoot nitrogen and shoot biomass under drought stress conditions, whereas under well-watered conditions, N decreased yield to 2597 kg ha
−1 relative to 2728 kg ha
−1 [
60]. Under severe drought stress, every 1 kg ha
−1 of N fertilizer resulted in extra 1.2 kg ha
−1 seeds [
72]. Seneviratne et al. [
73] reported that a relatively small amount of N fertilizer (46 kg/ha) significantly increased the seed yield by 84.7%. Similar conclusion was reported by Purcell and King [
60] that N fertilizer significantly increased the yield to 2798 kg ha
−1 compared to 2373 kg ha
−1 without N fertilizer; they associated this increase to increased seed number because of decreased flower and pod abortion.
Our results showed that drought-stressed treatment had, on average, less protein concentration compared to both irrigated counterparts. Similarly, Sepanlo et al. [
65] concluded that drought stress imposed on soybean plants at pod filling stage resulted in 15.5% reduction in protein concentration in the seeds. Reduced protein concentration under drought stress conditions was also reported by Turner et al. [
74] and Carrera et al. [
75]. Fertilization enhanced protein concentration under all irrigation regimes when applied in a high rate. Previously, Bloom [
76] reported that increasing applied-N rate was accompanied by enhanced protein concentration. Rotundo and Westgate [
77] reported that the addition of N fertilizer during the vegetative stages led to about 2% increase in protein concentration; they also concluded from their meta-analysis study that adding N fertilizer increased the protein content about 27% in all study environments; particularly, the increase was about 8% in field studies. N fertilizer dose also has a significant effect on the seed protein content; the dose of 100 kg/ha increased seed protein by just 2%, whereas the dose of 200 kg/ha resulted in 14% increase in seed protein [
78].
We found that drought stress resulted in better oil concentration compared to both irrigated treatments. Sepanlo et al. [
65] reported that drought at flowering stage increased oil concentration in soybean seeds by 5.7%, and further increased it (by 19.7%) when drought was imposed at pod filling stage. Boydak et al. [
79] also concluded that drought stress enhanced oil concentration.
Based on the current results, it can be concluded that the current climatic conditions in the study area have different effects on different traits of the selected soybean cultivar; however, the final seed yield is relatively stable under different irrigation regimes, most probably because this cultivar belongs to the middle maturity group (group I) which allows it to recover the possible effects of the short drought periods affecting the study area, especially with the fact that other certain traits (particularly seed oil concentration) were relatively enhanced by these periods of drought. Moreover, it could be confirmed that high N-fertilizer doses (105 kg ha−1 N in our case) did not result in measurably better yield except under non-irrigated regime, whereas a starter, small dose (35 kg ha−1 N) could result in better seed yield under all irrigation regimes. However, the high N-fertilizer rate resulted in the best seed protein concentration, regardless of irrigation regime, which might be of certain importance depending on the desired quality components in the produced seeds.
Based on the correlation analysis of the final seed yield with the studied traits (
Table 16), it can be noticed that periodical NDVI, compared to both SPAD and LAI, measurements can give an initial estimation of the expected yield and can be, consequently, an advisable, easy-to-apply practice. Plant height, in addition to the studied yield components (flower and pod number per plant and 100-seed weight) had also a strong correlation with the final seed yield.





{kind=link}
{kind=link}