Genetic Diversity among Cowpea (Vigna unguiculata (L.) Walp.) Landraces Suggests Central Mozambique as an Important Hotspot of Variation


 ,
,  ,
,  , ,
, ,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. DNA Extraction and nSSR Amplification
2.3. Genetic Diversity and Population Structure
2.4. Population Structure and Differentiation
2.5. Spatial Analysis and Genetic Diversity Rarefaction
3. Results
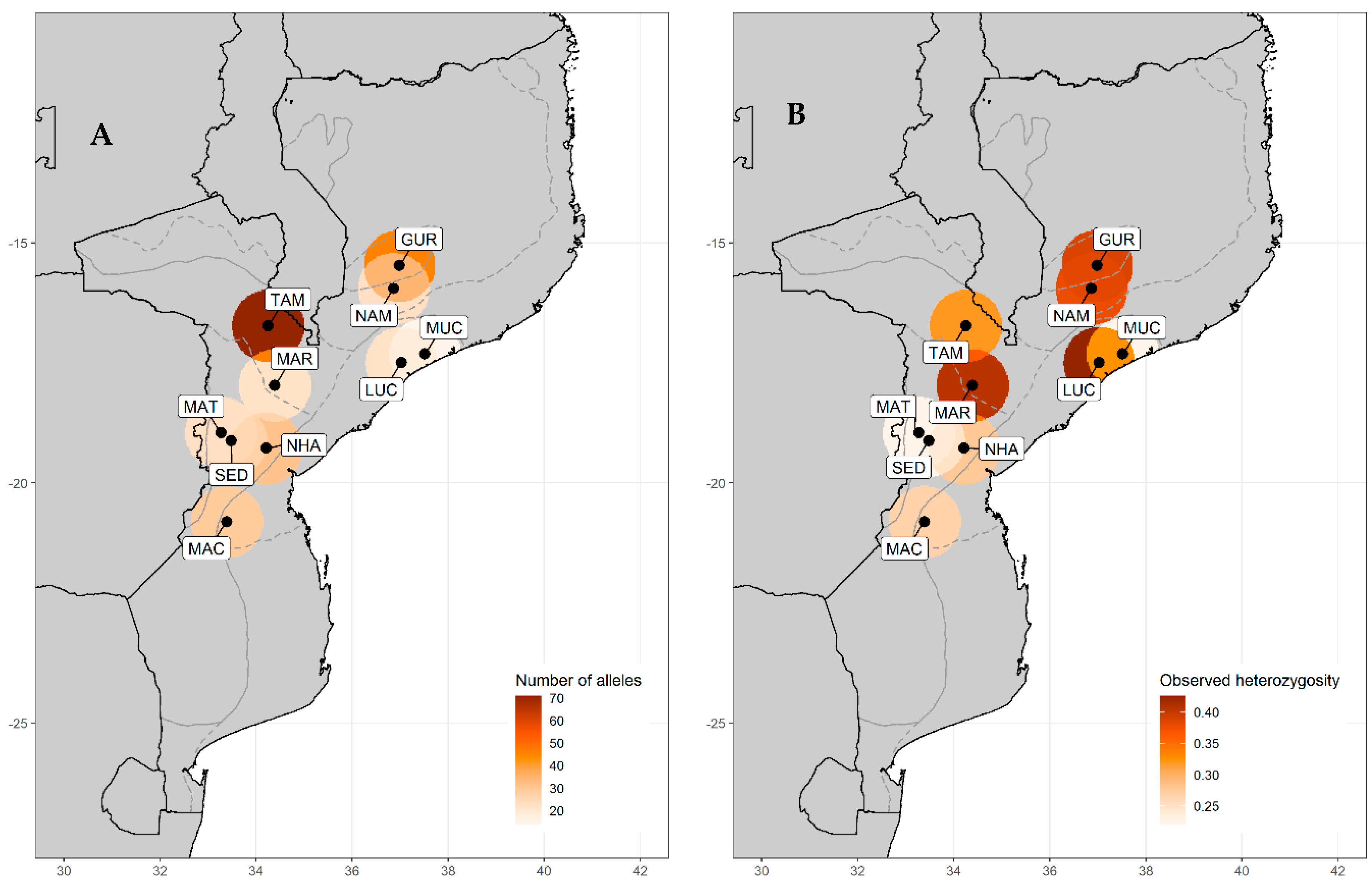
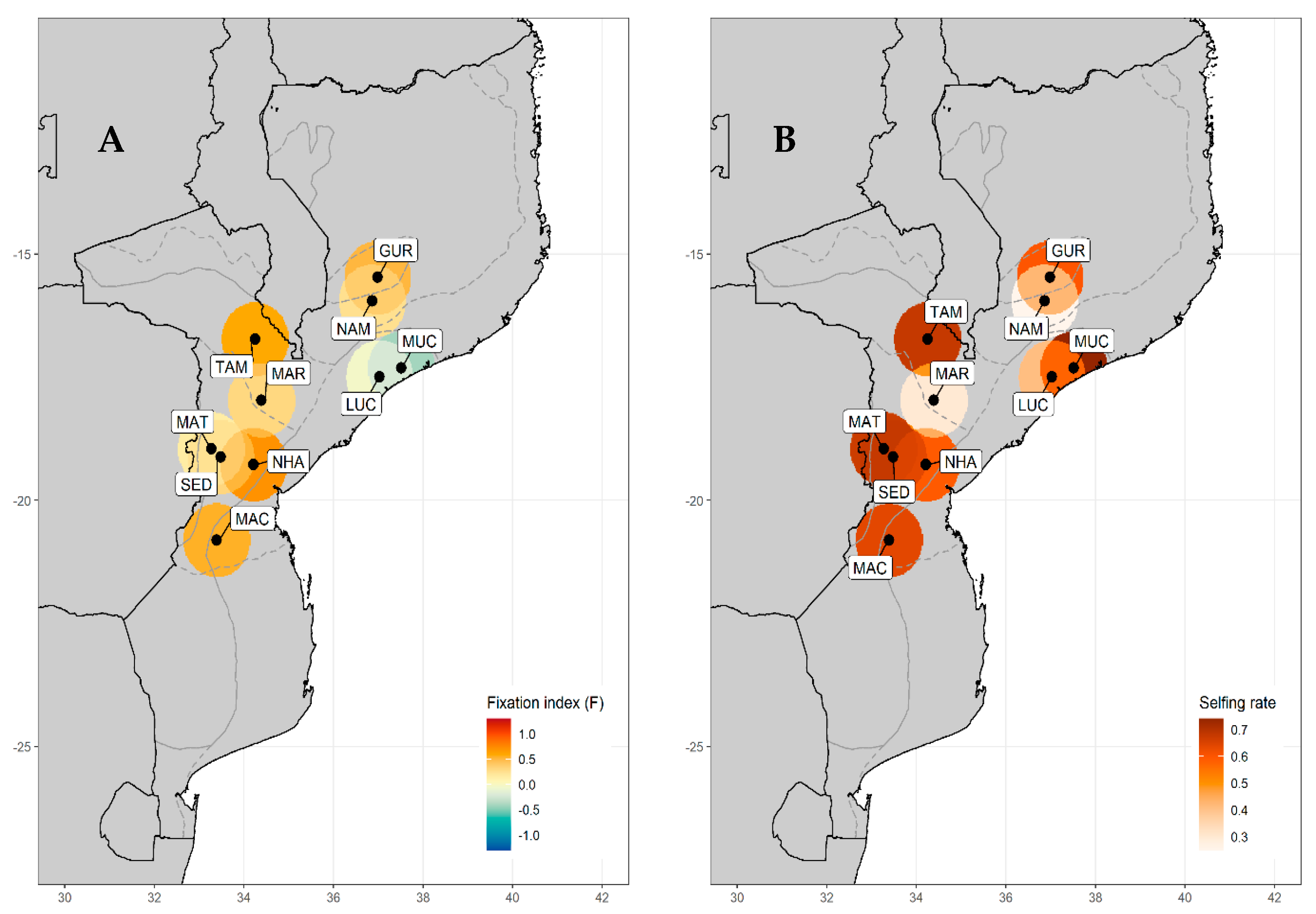
3.1. Genetic Diversity
3.2. Genetic Structure of V. unguiculata
3.3. Genetic Differentiation between Landraces
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Boukar, O.; Belko, N.; Chamarthi, S.; Togola, A.; Batieno, J.; Owusu, E.; Haruna, M.; Diallo, S.; Umar, M.L.; Olufajo, O.; et al. Cowpea (Vigna unguiculata): Genetics, genomics and breeding. Plant Breed. 2019, 138, 415–424. [Google Scholar] [CrossRef]
- Goncalves, A.; Goufo, P.; Barros, A.; Dominguez-Perles, R.; Trindade, H.; Rosa, E.A.; Ferreira, L.; Rodrigues, M. Cowpea (Vigna unguiculata L. Walp), a renewed multipurpose crop for a more sustainable agri-food system: Nutritional advantages and constraints. J. Sci. Food Agric. 2016, 96, 2941–2951. [Google Scholar] [CrossRef] [PubMed]
- Jayathilake, C.; Visvanathan, R.; Deen, A.; Bangamuwage, R.; Jayawardana, B.C.; Nammi, S.; Liyanage, R. Cowpea: An overview on its nutritional facts and health benefits. J. Sci. Food Agric. 2018, 98, 4793–4806. [Google Scholar] [CrossRef] [PubMed]
- Bado, B.; Bationo, A.; Cescas, M. Assessment of cowpea and groundnut contributions to soil fertility and succeeding sorghum yields in the Guinean savannah zone of Burkina Faso (West Africa). Biol. Fertil. Soils 2006, 43, 171–176. [Google Scholar] [CrossRef]
- Gomes, A.M.F.; Nhantumbo, N.; Ferreira-Pinto, M.; Massinga, R.; Ramalho, J.C.; Ribeiro-Barros, A. Breeding Elite Cowpea [Vigna unguiculata (L.) Walp] Varieties for Improved Food Security and Income in Africa: Opportunities and Challenges. In Legume Crops; El-Esawi, M.A., Ed.; IntechOpen: London, UK, 2019; p. 636. [Google Scholar]
- FAO. Agro-Ecological Zoning. Guidelines. FAO Soils Bulletin Soil Resources, Management and Conservation Service, FAO Land and Water Development Division; Food and Agriculture Organization of the United Nations: Rome, Italy, 1996; pp. 73–78. ISBN 92-5-103890-2. [Google Scholar]
- Goufo, P.; Moutinho-Pereira, J.M.; Jorge, T.F.; Correia, C.M.; Oliveira, M.R.; Rosa, E.A.S.; António, C.; Trindade, H. Cowpea (Vigna unguiculata L. Walp.) metabolomics: Osmoprotection as a physiological strategy for drought stress resistance and improved yield. Front. Plant Sci. 2017, 8, 586. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.M.F.; Rodrigues, A.P.; António, C.; Rodrigues, A.M.; Leitão, A.E.; Batista-Santos, P.; Nhantumbo, N.; Massinga, R.; Ribeiro-Barros, A.I.; Ramalho, J.C. Drought response of cowpea (Vigna unguiculata (L.) Walp.) landraces at leaf physiological and metabolite profile levels. Environ. Exp. Bot. 2020, 175, 104060. [Google Scholar] [CrossRef]
- Carvalho, M.; Lino-Neto, T.; Rosa, E.; Carnide, V. Cowpea: A legume crop for a challenging environment. J. Sci. Food Agric. 2017, 97, 4273–4284. [Google Scholar] [CrossRef]
- Xiong, H.; Shi, A.; Mou, B.; Qin, J.; Motes, D.; Lu, W.; Ma, J.; Weng, Y.; Yang, W.; Wu, D. Genetic diversity and population structure of cowpea (Vigna unguiculata L. Walp). PLoS ONE 2016, 11, e0160941. [Google Scholar] [CrossRef]
- Fatokun, C.; Girma, G.; Abberton, M.; Gedil, M.; Unachukwu, N.; Oyatomi, O.; Yusuf, M.; Rabbi, I.; Boukar, O. Genetic diversity and population structure of a mini-core subset from the world cowpea (Vigna unguiculata (L.) Walp.) germplasm collection. Sci. Rep. 2018, 8, 16035. [Google Scholar] [CrossRef]
- Huynh, B.; Close, T.J.; Roberts, P.A.; Hu, Z.; Wanamaker, S.; Lucas, M.R.; Chiulele, R.; Cissé, N.; David, A.; Hearne, S.; et al. Gene pools and the genetic architecture of domesticated cowpea. Plant Genome 2013, 6, 1–8. [Google Scholar] [CrossRef]
- Fang, J.; Chao, C.T.; Roberts, P.A.; Ehlers, J.D. Genetic diversity of cowpea [Vigna unguiculata (L.) Walp.] in four West African and USA breeding programs as determined by AFLP analysis. Genet. Resour. Crop Evol. 2007, 54, 1197–1209. [Google Scholar] [CrossRef]
- Mahalakshmi, V.; Ng, Q.; Lawson, M.; Ortiz, R. Cowpea [Vigna unguiculata (L.) Walp.] core collection defined by geographical, agronomical and botanical descriptors. Plant Genet. Resour. 2007, 5, 113–119. [Google Scholar] [CrossRef]
- Ehlers, J.D.; Hall, A.E. Cowpea (Vigna unguiculata (L.) Walp.). Field Crops Res. 1997, 53, 187–204. [Google Scholar] [CrossRef]
- Ficiciyan, A.; Loos, J.; Sievers-Glotzbach, S.; Tscharntke, T. More than yield: Ecosystem services of traditional versus modern crop varieties revisited. Sustainability 2018, 10, 2834. [Google Scholar] [CrossRef]
- De Luca, D.; Cennamo, P.; Del Guacchio, E.; Di Novella, R.; Caputo, P. Conservation and genetic characterisation of common bean landraces from Cilento region (southern Italy): High differentiation in spite of low genetic diversity. Genetica 2018, 146, 29–44. [Google Scholar] [CrossRef]
- Alipour, H.; Bihamta, M.R.; Mohammadi, V.; Peyghambari, S.A.; Bai, G.; Zhang, G. Genotyping-by-sequencing (GBS) revealed molecular genetic diversity of Iranian wheat landraces and cultivars. Front. Plant Sci. 2017, 8, 1293. [Google Scholar] [CrossRef]
- Ram, S.G.; Thiruvengadam, V.; Vinod, K.K. Genetic diversity among cultivars, landraces and wild relatives of rice as revealed by microsatellite markers. J. Appl. Genet. 2007, 48, 337–345. [Google Scholar] [CrossRef]
- Hagenblad, J.; Zie, J.; Leino, M.W. Exploring the population genetics of genebank and historical landrace varieties. Genet. Resour. Crop Evol. 2012, 59, 1185–1199. [Google Scholar] [CrossRef]
- Ministry of Agriculture and Fisheries (MAF). Agroecological zones and production systems. In Working Document 2/B, Program of Investment in Agricultural Extension, Process of the Formulation of Proagri; Ministry of Agriculture and Fisheries: Maputo, Mozambique, 1996; p. 72. [Google Scholar]
- Gupta, S.K.; Gopalakrishna, T. Development of unigene-derived SSR markers in cowpea (Vigna unguiculata) and their transferability to other Vigna species. Genome 2010, 53, 508–523. [Google Scholar] [CrossRef]
- Goudet, J. FSTAT, a Program to Estimate and Test Gene Diversities and Fixation Indices (Version 2.9.3.2). 2002. Available online: http://www.unil.ch/izea/softwares/fstat (accessed on 3 November 2019).
- Peakall, R.; Smouse, P.E. GenAlEx 6 genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Ritland, K. Inferences about inbreeding depression based on changes of the inbreeding coefficient. Evolution 1990, 44, 1230–1241. [Google Scholar] [CrossRef] [PubMed]
- Raymond, M.; Rousset, F. GENEPOP (version 1.2): Population genetics software for exact tests and ecumenicism. J. Hered. 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Rousset, F. Genepop’007: A complete reimplementation of the Genepop software for Windows and Linux. Mol. Ecol. Resour. 2008, 8, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Rice, W.R. Analyzing tables of statistical tests. Evolution 1989, 43, 223–225. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Rosenberg, N.A.; Donnelly, P. Association mapping in structured populations. Am. J. Hum. Genet. 2000, 67, 170–181. [Google Scholar] [CrossRef] [PubMed]
- Earl, D.A.; von Holdt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 8, 2611–2620. [Google Scholar] [CrossRef]
- Jakobsson, M.; Rosenberg, N.A. CLUMPP: A cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef]
- Langella, O. Populations 1.2.30. Laboratoire Populations. Genetique et Evolution. Centre National de la Recherche Scientifique; CNRS UPR9034: Gif Sur Yvette, France, 2007; Available online: http://bioinformatics.org/populations/ (accessed on 3 November 2019).
- Nei, M.; Tajima, F.; Tateno, Y. Accuracy of estimated phylogenetic trees from molecular data. II. Gene frequency data. J. Mol. Evol. 1983, 19, 153–170. [Google Scholar] [CrossRef]
- Excoffier, L.; Laval, G.; Schneider, S. ARLEQUIN, Ver 3.0. An integrated software package for population genetic data analysis. Evol. Bionf. Online 1983, 1, 47–50. [Google Scholar]
- Lopes, M.S.; El-Basyoni, I.; Baenziger, P.S.; Singh, S.; Royo, C.; Ozbek, K.; Aktas, H.; Ozer, E.; Ozdemir, F.; Manickavelu, A.; et al. Exploiting genetic diversity from landraces in wheat breeding for adaptation to climate change. J. Exp. Bot. 2015, 66, 3477–3486. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, S.L.; Ceccarelli, S.; Blair, M.W.; Upadhyaya, H.D.; Are, A.K.; Ortiz, R. Landrace germplasm for improving yield and abiotic stress adaptation. Trends Plant Sci. 2016, 21, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Monteagudo, A.; Casas, A.M.; Cantalapiedra, C.P.; Contreras-Moreira, B.; Gracia, M.P.; Igartua, E. Harnessing novel diversity from landraces to improve an elite barley variety. Front. Plant Sci. 2019, 10, 434. [Google Scholar] [CrossRef] [PubMed]
- Hour, A.; Hsieh, W.; Chang, S.; Wu, Y.; Chin, H.; Lin, Y. Genetic diversity of landraces and improved varieties of rice (Oryza sativa L.) in Taiwan. Rice 2020. [Google Scholar] [CrossRef]
- Ganeva, G.M.; Korzun, V.; Landjeva, S.; Popova, Z.; Christov, N.K. Genetic diversity assessment of Bulgarian durum wheat (Triticum durum Desf.) landraces and modern cultivars using microsatellite markers. Genet. Resour. Crop Evol. 2010, 57, 273–285. [Google Scholar] [CrossRef]
- Kumbhar, S.D.; Kulwal, P.L.; Patil, J.V.; Sarawate, C.D.; Gaikwad, A.P.; Jadhav, A.S. Genetic diversity and population structure in landraces and improved rice varieties from India. Rice Sci. 2015, 22, 99–107. [Google Scholar] [CrossRef]
- Soriano, J.M.; Villegas, D.; Aranzana, M.J.; García del Moral, L.F.; Royo, C. Genetic structure of modern durum wheat cultivars and Mediterranean landraces matches with their agronomic performance. PLoS ONE 2016, 11, e0160983. [Google Scholar] [CrossRef] [PubMed]
- Pascual, L.; Ruiz, M.; López-Fernández, M.; Pérez-Peña, H.; Benavente, H.; Vázquez, J.F.; Sansaloni, C.; Giraldo, P. Genomic analysis of Spanish wheat landraces reveals their variability and potential for breeding. BMC Genom. 2020, 21, 122. [Google Scholar] [CrossRef]
- FAO/IPGRI. Review and Development of Indicators for Genetic Diversity, Genetic Erosion and Genetic Vulnerability (GDEV): Summary Report of a Joint FAO/IPGRI Workshop; FAO/IPGRI: Rome, Italy, 2002. [Google Scholar]
- Carvalho, M.; Muñoz-Amatriaín, M.; Castro, I.; Lino-Neto, T.; Matos, M.; Egea-Cortines, M.; Rosa, E.; Close, T.; Carnide, V. Genetic diversity and structure of Iberian Peninsula cowpeas compared to world-wide cowpea accessions using high density SNP markers. BMC Genom. 2017, 18, 891. [Google Scholar] [CrossRef]
- Chen, H.; Chen, H.; Hu, L.; Wang, L.; Wang, S.; Wang, M.L.; Cheng, X. Genetic diversity and a population structure analysis of accessions in the Chinese cowpea [Vigna unguiculata (L.) Walp.] germplasm collection. Crop J. 2017, 5, 363–372. [Google Scholar] [CrossRef]
- Zannouou, A.; Kossou, D.K.; Ahanchede, A.; Zoundjihékpon, J.; Agbicodo, E.; Struik, P.C.; Sanni, A. Genetic variability of cultivated cowpea in Benin assessed by random amplified polymorphic DNA. Afr. J. Biotechnol. 2008, 7, 24. [Google Scholar]
- Egbadzor, K.F.; Danquah, E.Y.; Ofori, K.; Yeboah, M.; Offei, S.K. Diversity in 118 cowpea (Vigna unguiculata (L.) Walp.) accessions assessed with 16 morphological traits. Int. J. Plant Breed. Genet. 2014, 8, 35–43. [Google Scholar] [CrossRef]
- Ali, Z.B.; Yao, K.N.; Odeny, D.A.; Kyalo, M.; Skilton, R.; Eltahir, I.M. Assessing the genetic diversity of cowpea (Vigna unguiculata (L.) Walp.) accessions from Sudan using simple sequence repeat (SSR)markers. Afr. J. Plant Sci. 2015, 9, 293–304. [Google Scholar] [CrossRef]
- Njonjo, M.W.; Muthomi, J.W.; Mwang’ombe, A.W. Production practices, postharvest handling, and quality of cowpea seed used by farmers in Makueni and Taita Taveta counties in Kenya. Int. J. Agron. 2019. [Google Scholar] [CrossRef]
- Kimani, P.; Kamundia, D.K.; Narla, R.D.; Mwang’ombe, A.W. An integrated seed delivery system and seed research in Kenya: Case studies of pigeon pea, onion and dry bean. In Seed Systems, Science, and Policy in East and Central Africa; Francis, J.A., Ed.; CTA: The Hague, The Netherlands, 2014; pp. 73–88. [Google Scholar]
- Hall, A. Breeding for adaptation to drought and heat in cowpea. Eur. J. Agron. 2004, 21, 447–454. [Google Scholar] [CrossRef]
- Muchero, W.; Ehlers, J.D.; Roberts, P.A. Seedling stage drought-induced phenotypes and drought-responsive genes in diverse cowpea genotypes. Crop Sci. 2008, 48, 541–552. [Google Scholar] [CrossRef]
- Mbeyagala, E.K.; Mukasa, B.S.; Tukamuhabwa, P.; Bisikwa, J. Evaluation of cowpea genotypes for virus resistance under natural conditions in Uganda. J. Agric. Sci. 2014, 6, 176–187. [Google Scholar] [CrossRef]
- Bellon, M.R.; van Etten, J. Climate change and on-farm conservation of crop landraces in centres of diversity In Plant Genetic Resources and Climate Change; CABI Publishing: Egham, UK, 2014; pp. 137–150. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence 5′-3′ | Na | Ho | He | FIS |
|---|---|---|---|---|---|
| VuUGM33 | F: AAAGGTGGGGGATTATGAGG | 83 | 0.907 | 0.806 | −0.091 |
| R: TGTCCAATCCTGATGGATGA | |||||
| VuUGM71 | F: TTCACAACCTGTCCACCTCA | 125 | 0.143 | 0.548 | 0.783 |
| R: GGCGTCCCAACAGATAAGAA | |||||
| VuUGM05 | F: GCGGGATTCTATTCCAGTGA | 82 | 0.174 | 0.617 | 0.767 |
| R: TCCATTGGGTTTCTCAACCT | |||||
| VuUGM39 | F: CGAAAAAGCATGATCAACCA | 97 | 0.149 | 0.749 | 0.851 |
| R: CCCCTTTCGCTAAAATTTCC | |||||
| VuUGM22 | F: CAATCACCATTCACCAAACA | 112 | 0.181 | 0.629 | 0.749 |
| R: TATTGGGACTCAGGTCTTGG | |||||
| VuUGM31 | F: TGGTTCACTTCCCATATTGTC | 122 | 0.136 | 0.711 | 0.857 |
| R: AGGCAGAGACGAAGGAGTGA | |||||
| VuUGM40 | F: TTCTACATGGTTTTGGGGTCA | 145 | 1.003 | 0.671 | −0.426 |
| R: GAGCTTGCCCTCAAGAATTG | |||||
| VuUGM68 | F: TGATTGATGGTGGTGTAGCC | 59 | 0.415 | 0.397 | −0.008 |
| R: GCACTTCACTCATCGTTGCT | |||||
| VuUGM74 | F: GCCTCCTCTCACAAACTTGC | 49 | 0.014 | 0.016 | 0.018 |
| Landrace Identity | Province | AEZ | N | NA | AR | Ho | He | FIS | s |
|---|---|---|---|---|---|---|---|---|---|
| Gurué (GUR) | North Zambezia | R10 | 6 | 46 | 1.751 | 0.389 | 0.688 | 0.506 | 60% |
| Namarroi (NAM) | North Zambezia | R7 | 4 | 23 | 1.534 | 0.379 | 0.454 | 0.250 | 25% |
| Muchela (MUC) | Central Zambezia | R7 | 4 | 14 | 1.250 | 0.222 | 0.535 | −0.412 | 74% |
| Lucas Branco (LUC) | South Zambezia | R7 | 4 | 22 | 1.432 | 0.426 | 0.577 | −0.032 | 41% |
| Nhamatanda (NHA) | Central Sofala | R4 | 4 | 31 | 1.682 | 0.278 | 0.479 | 0.707 | 59% |
| Maringué (MAR) | Central Sofala | R5 | 3 | 22 | 1.503 | 0.407 | 0.494 | 0.310 | 30% |
| Tambara (TAM) | North Manica | R6 | 23 | 71 | 1.612 | 0.320 | 0.654 | 0.592 | 68% |
| Sede nova (SED) | North Manica | R6 | 3 | 23 | 1.562 | 0.222 | 0.451 | 0.323 | 69% |
| Matsinho (MAT) | Central Manica | R4 | 3 | 23 | 1.577 | 0.222 | 0.451 | 0.156 | 67% |
| Machaze (MAC) | South Manica | R3 | 5 | 29 | 1.555 | 0.267 | 0.500 | 0.549 | 64% |
| IT-16 | Commercial cultivar | R4 | 1 | 12 | 1.333 | 0.333 | 0.167 | - | |
| IT-18 | Commercial cultivar | R6 | 1 | 11 | 1.222 | 0.222 | 0.111 | - |
| Source of Variance | Degrees of Freedom | Sum of Squares | % of Variance | |
|---|---|---|---|---|
| Among landraces | ||||
| Among AEZs | 6 | 77.612 | 19.92 | |
| Among genotypes within landraces | 54 | 207.109 | 47.39 | |
| Within genotypes | 61 | 60.001 | 32.69 | |
| Among cowpea accessions vs. cultivars | ||||
| Among accessions | 1 | 4.772 | 0.12 | |
| Among accessions | 58 | 279.949 | 65.58 | |
| Within accessions | 61 | 60.000 | 34.30 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomes, A.M.F.; Draper, D.; Talhinhas, P.; Santos, P.B.; Simões, F.; Nhantumbo, N.; Massinga, R.; Ramalho, J.C.; Marques, I.; Ribeiro-Barros, A.I. Genetic Diversity among Cowpea (Vigna unguiculata (L.) Walp.) Landraces Suggests Central Mozambique as an Important Hotspot of Variation. Agronomy 2020, 10, 1893. https://doi.org/10.3390/agronomy10121893
Gomes AMF, Draper D, Talhinhas P, Santos PB, Simões F, Nhantumbo N, Massinga R, Ramalho JC, Marques I, Ribeiro-Barros AI. Genetic Diversity among Cowpea (Vigna unguiculata (L.) Walp.) Landraces Suggests Central Mozambique as an Important Hotspot of Variation. Agronomy. 2020; 10(12):1893. https://doi.org/10.3390/agronomy10121893
Chicago/Turabian StyleGomes, Ana Maria Figueira, David Draper, Pedro Talhinhas, Paula Batista Santos, Fernanda Simões, Nascimento Nhantumbo, Rafael Massinga, José C. Ramalho, Isabel Marques, and Ana I. Ribeiro-Barros. 2020. "Genetic Diversity among Cowpea (Vigna unguiculata (L.) Walp.) Landraces Suggests Central Mozambique as an Important Hotspot of Variation" Agronomy 10, no. 12: 1893. https://doi.org/10.3390/agronomy10121893
APA StyleGomes, A. M. F., Draper, D., Talhinhas, P., Santos, P. B., Simões, F., Nhantumbo, N., Massinga, R., Ramalho, J. C., Marques, I., & Ribeiro-Barros, A. I. (2020). Genetic Diversity among Cowpea (Vigna unguiculata (L.) Walp.) Landraces Suggests Central Mozambique as an Important Hotspot of Variation. Agronomy, 10(12), 1893. https://doi.org/10.3390/agronomy10121893