Aerobic Rice with or without Strategic Irrigation in the Subtropics




Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Weather
2.2. Experimental Design and Treatments
2.3. Crop Management
2.4. Rice Varieties
2.5. Crop and Yield Parameters
2.6. Above-Ground Development, Growth, and Gas Exchange Parameters
- L = length of flag leaf in centimetres;
- B = breadth of flag leaf from widest portion in centimetres;
- C = constant factor, which is 0.74 for the flag leaf of rice.
2.7. Root Parameters
2.8. Water Productivity
2.9. Data Analysis
3. Results
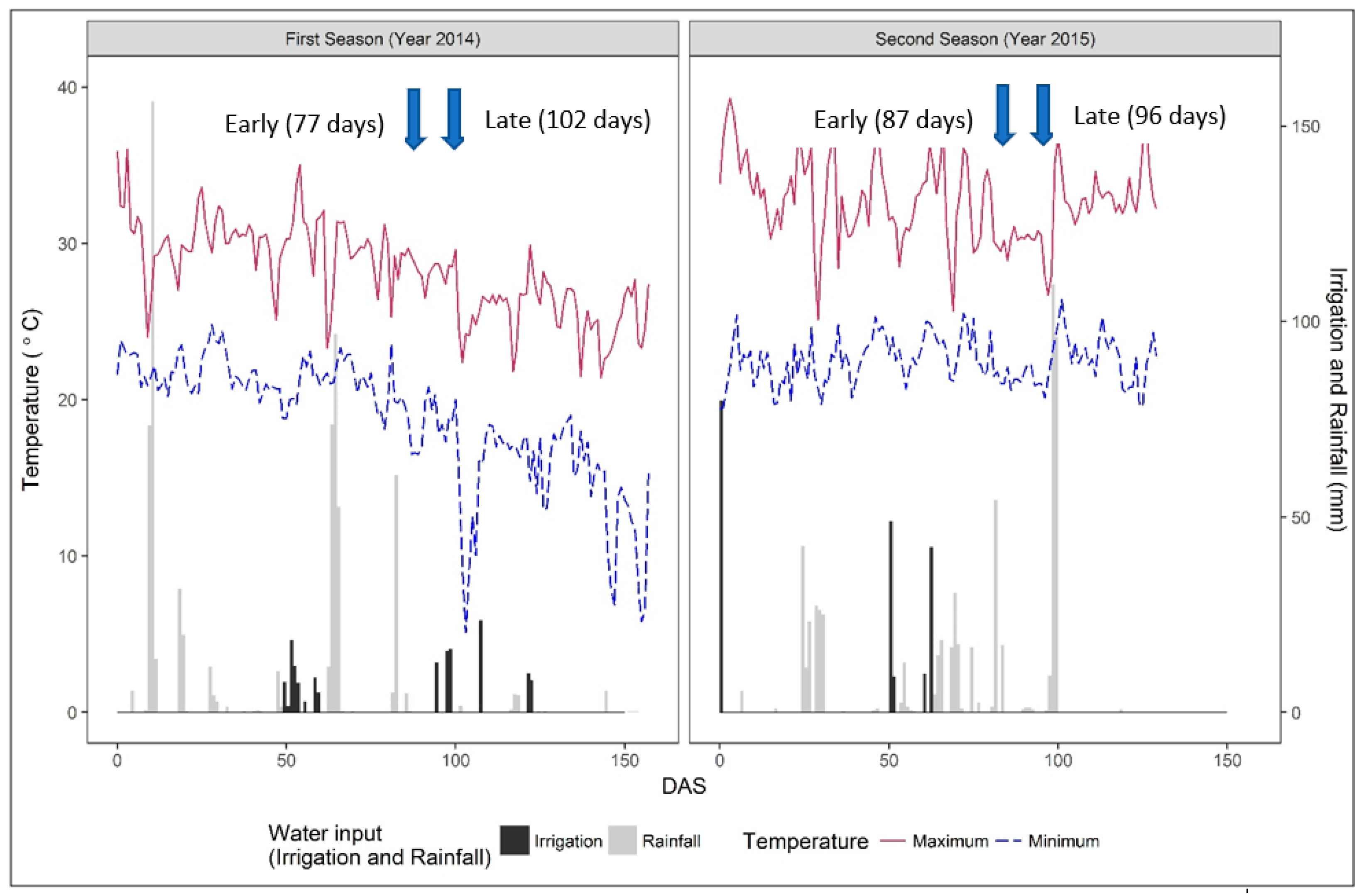
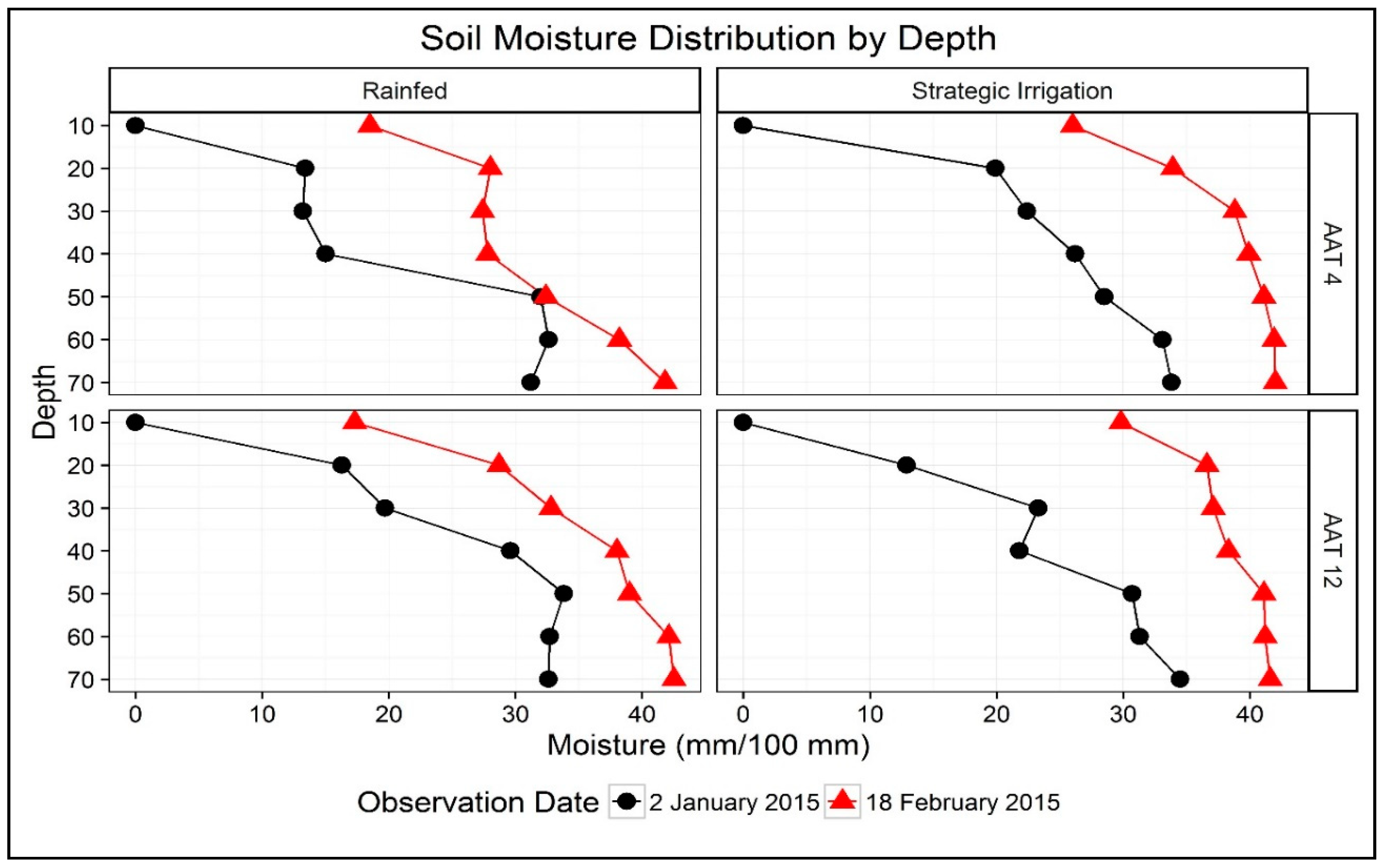
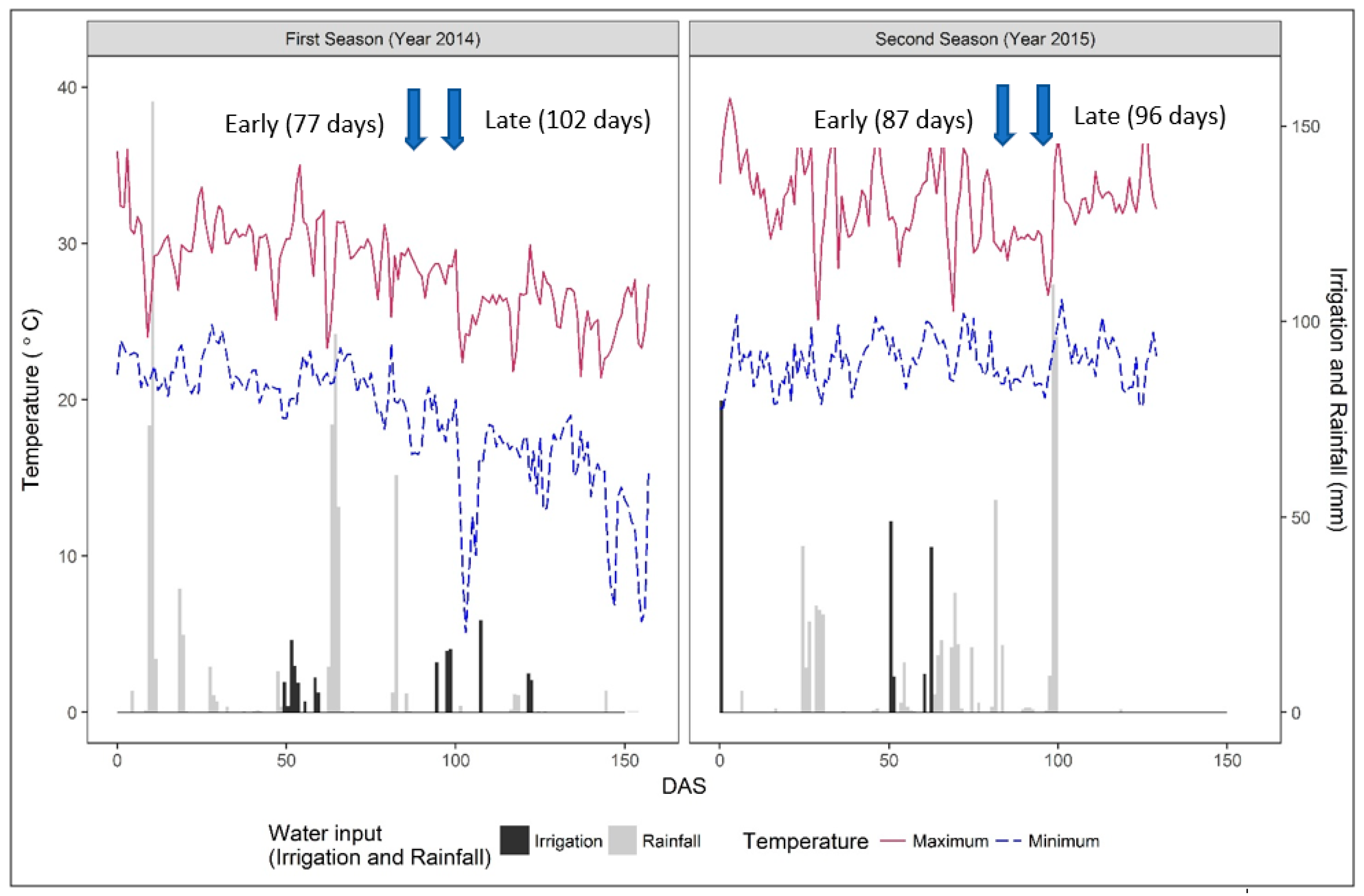
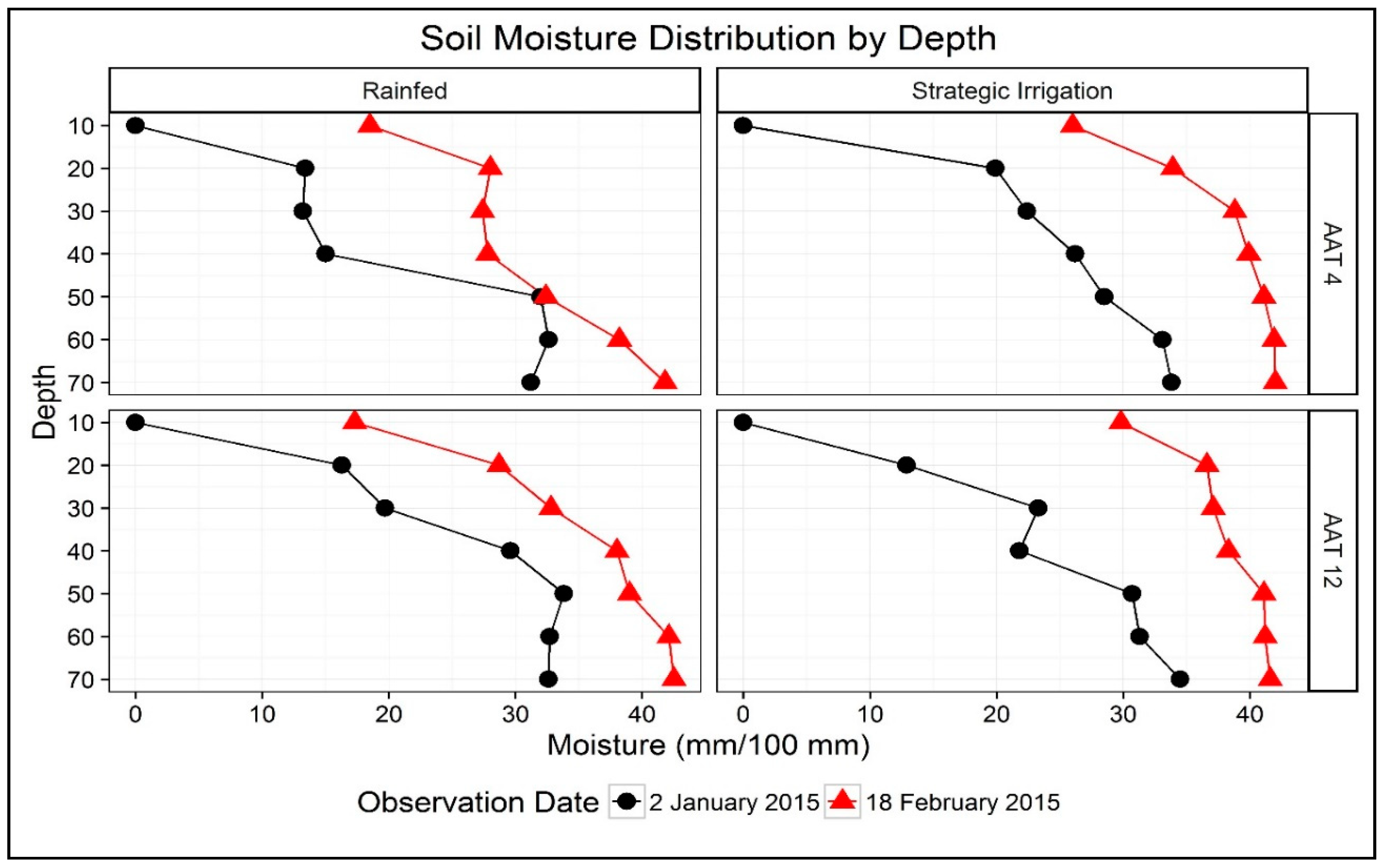
3.1. Weather Parameters, Irrigation Inputs, and Soil Moisture Dynamics
3.2. Above-Ground Growth and Development and Gas Exchange Parameters
3.2.1. Days to Flowering
3.2.2. Leaf Area Index
3.2.3. Leaf Chlorophyll Concentration
3.2.4. Leaf Gas Exchange Parameters and Instantaneous Water Use Efficiency (WUE)
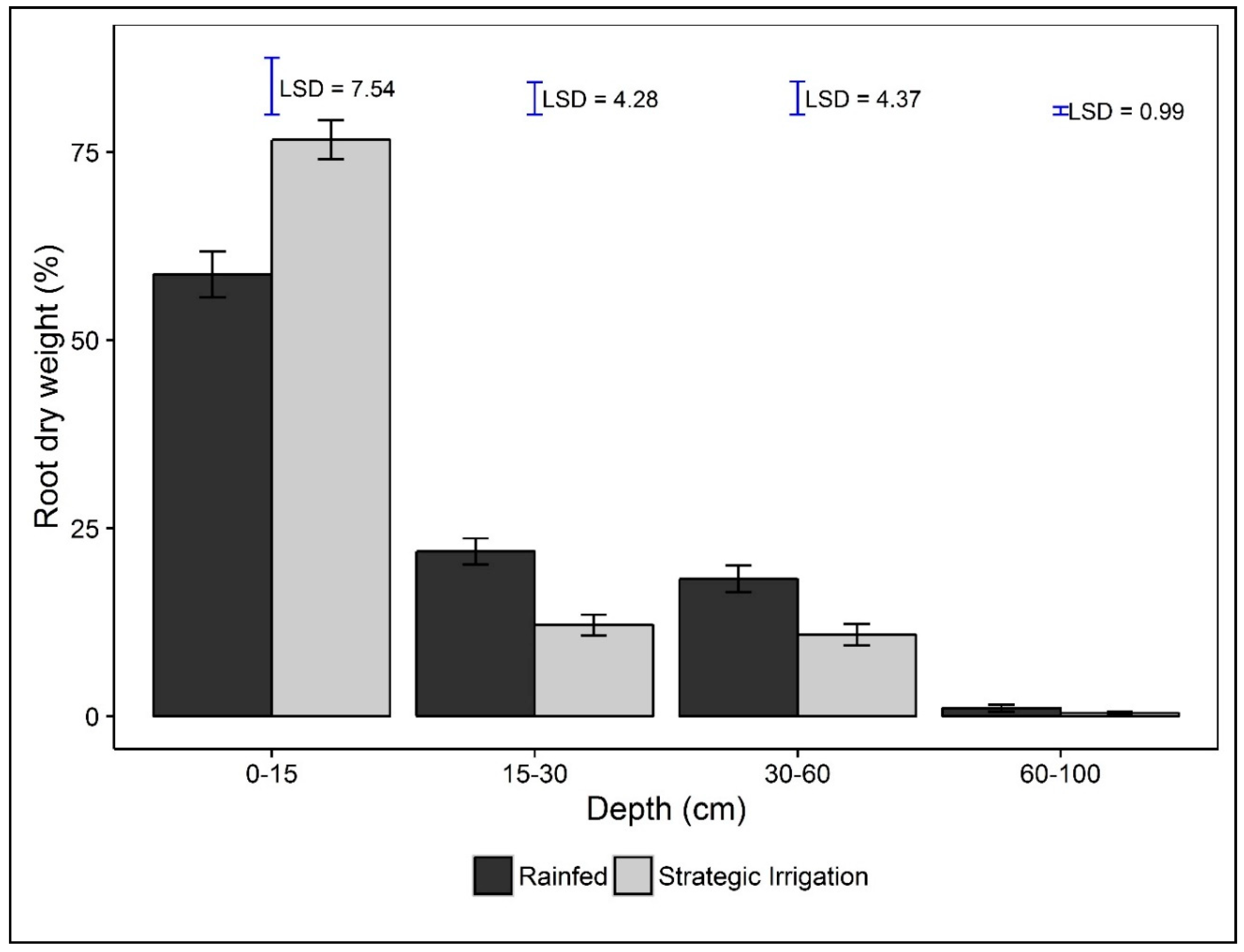
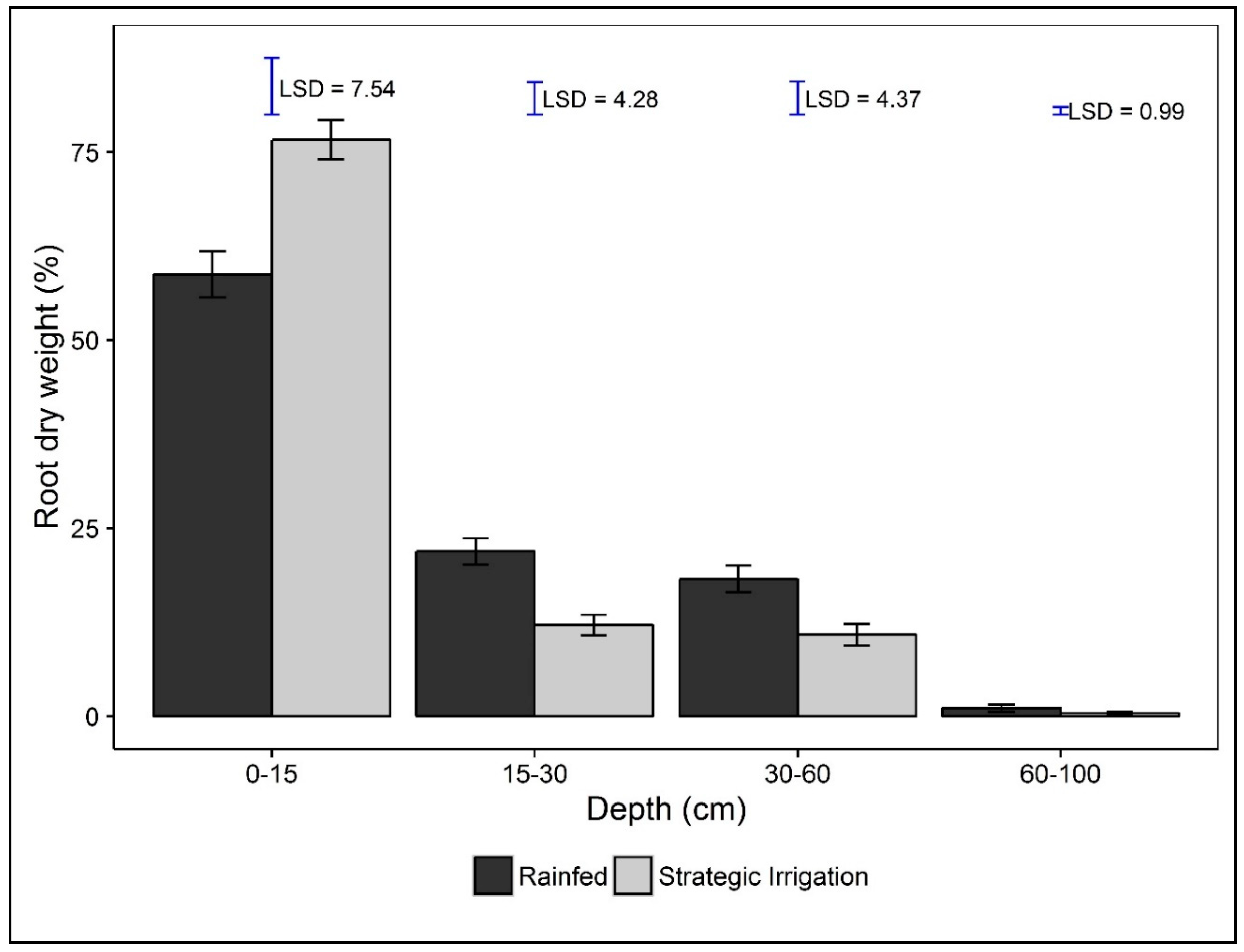
3.3. Root Parameters
3.4. Yield and Its Components
3.4.1. Straw and Aboveground Biomass Yields
3.4.2. Sample Plot Grain Yields
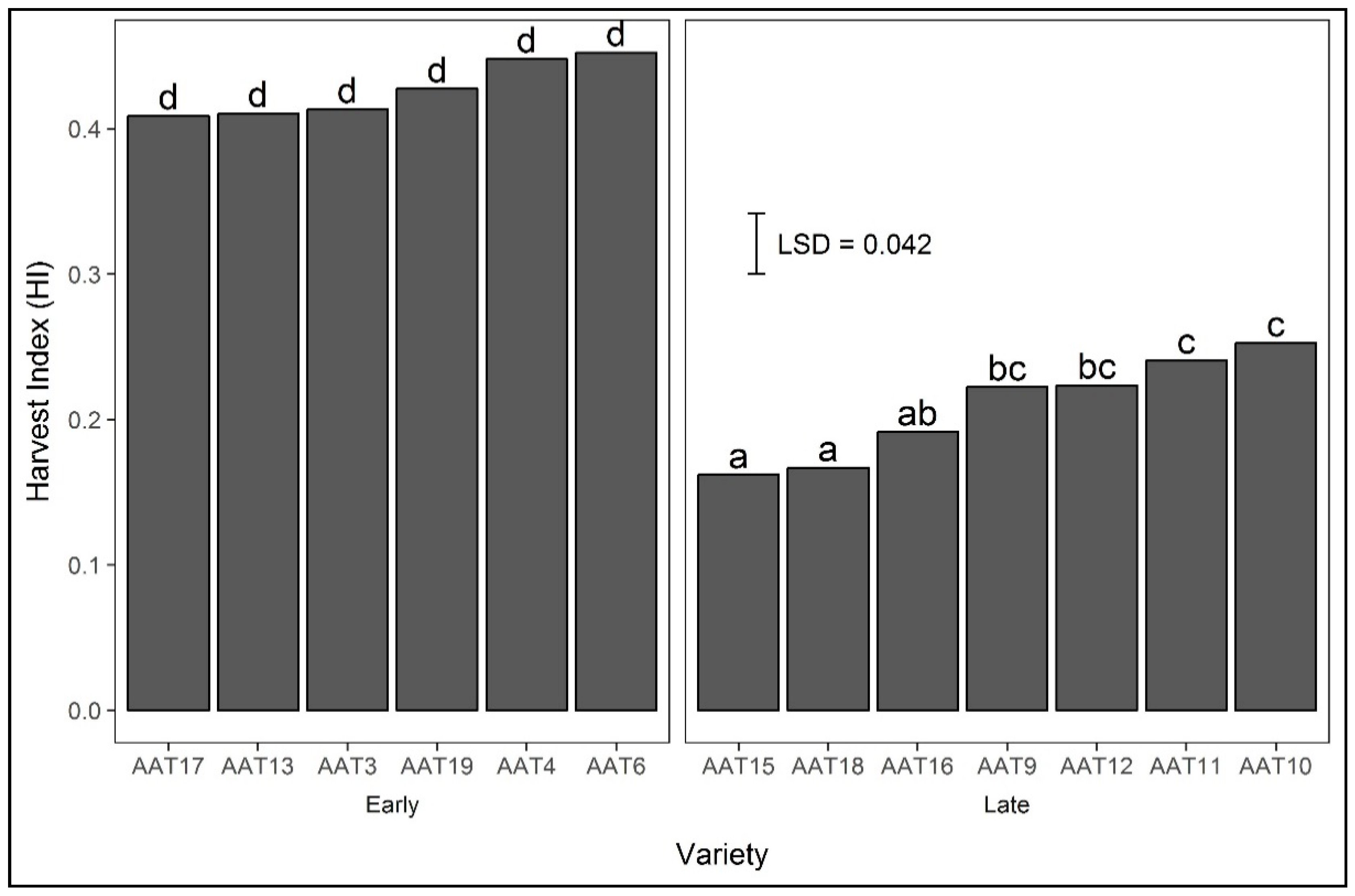
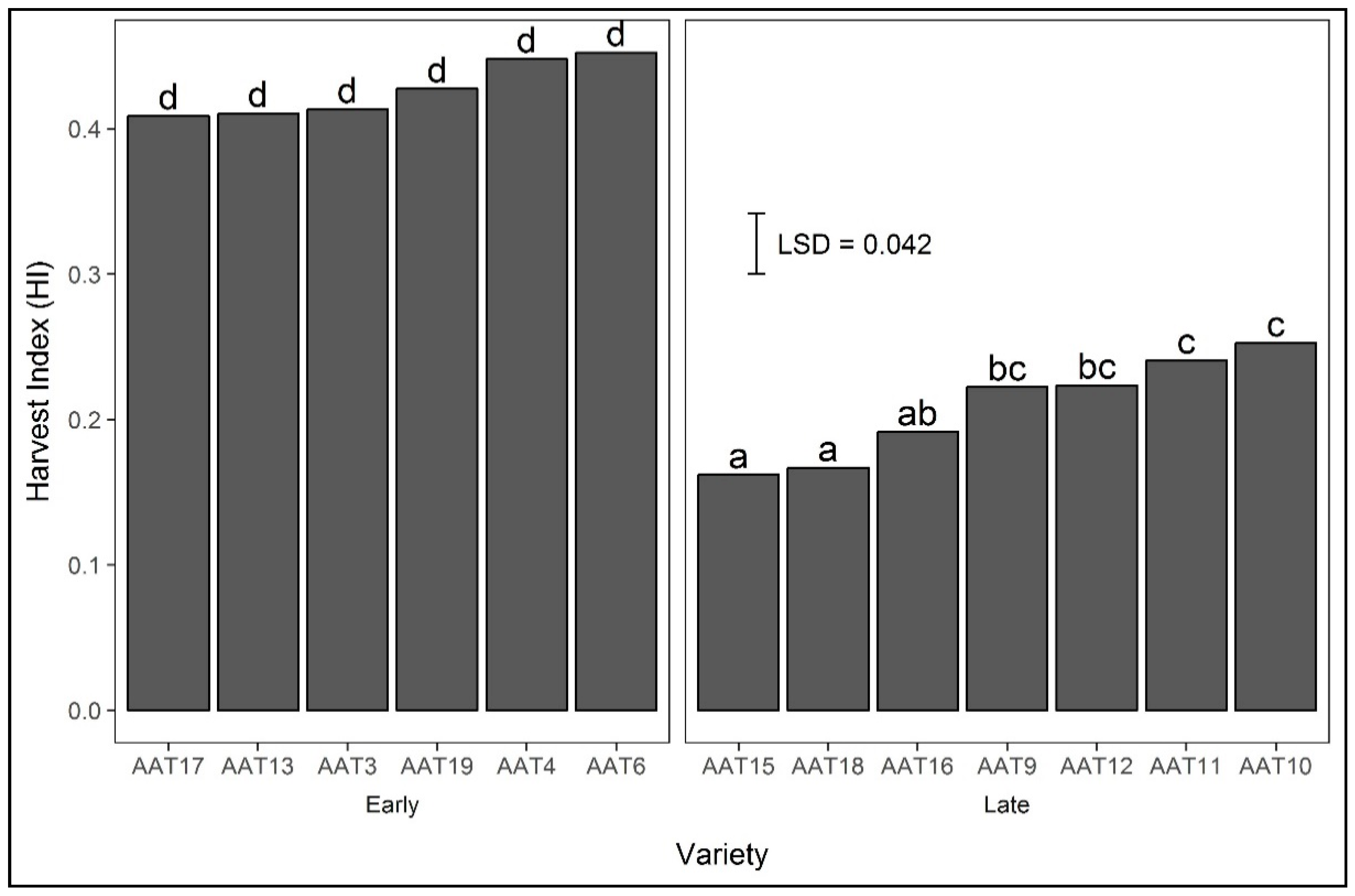
3.4.3. Harvest Index
3.4.4. Yield Attributing Parameters
3.5. Water Productivity
4. Discussion
4.1. Influence of Planting Time, Maturity Class, and Strategic Irrigation and Phenology on Yields
4.2. Effects of Planting Time, Maturity Class, and Strategic Irrigation on Light Capture and Use and the Influence on Yields
4.3. Effects of Planting Time, Maturity Class, and Strategic Irrigation on Root Growth and the Influence on Yields
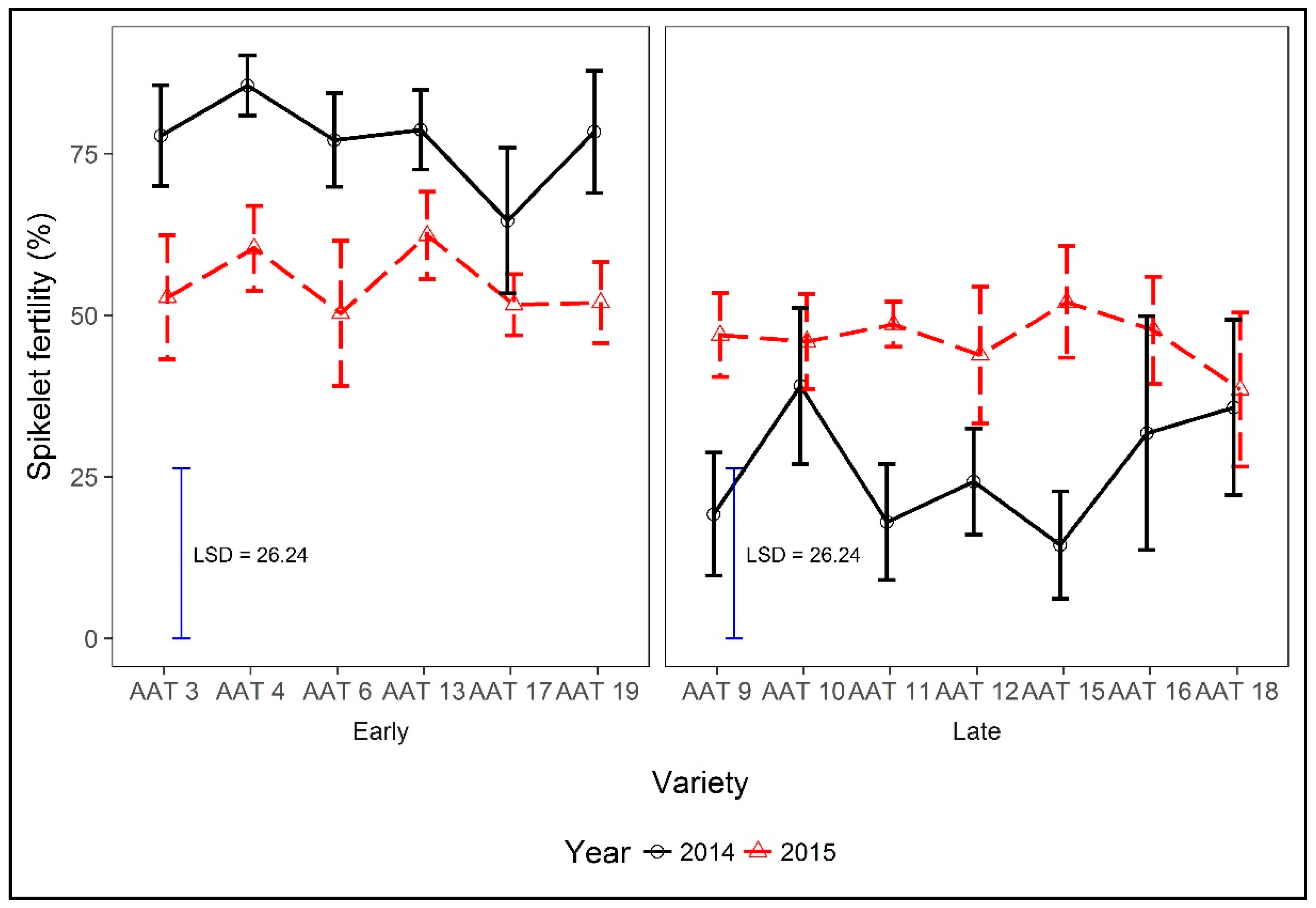
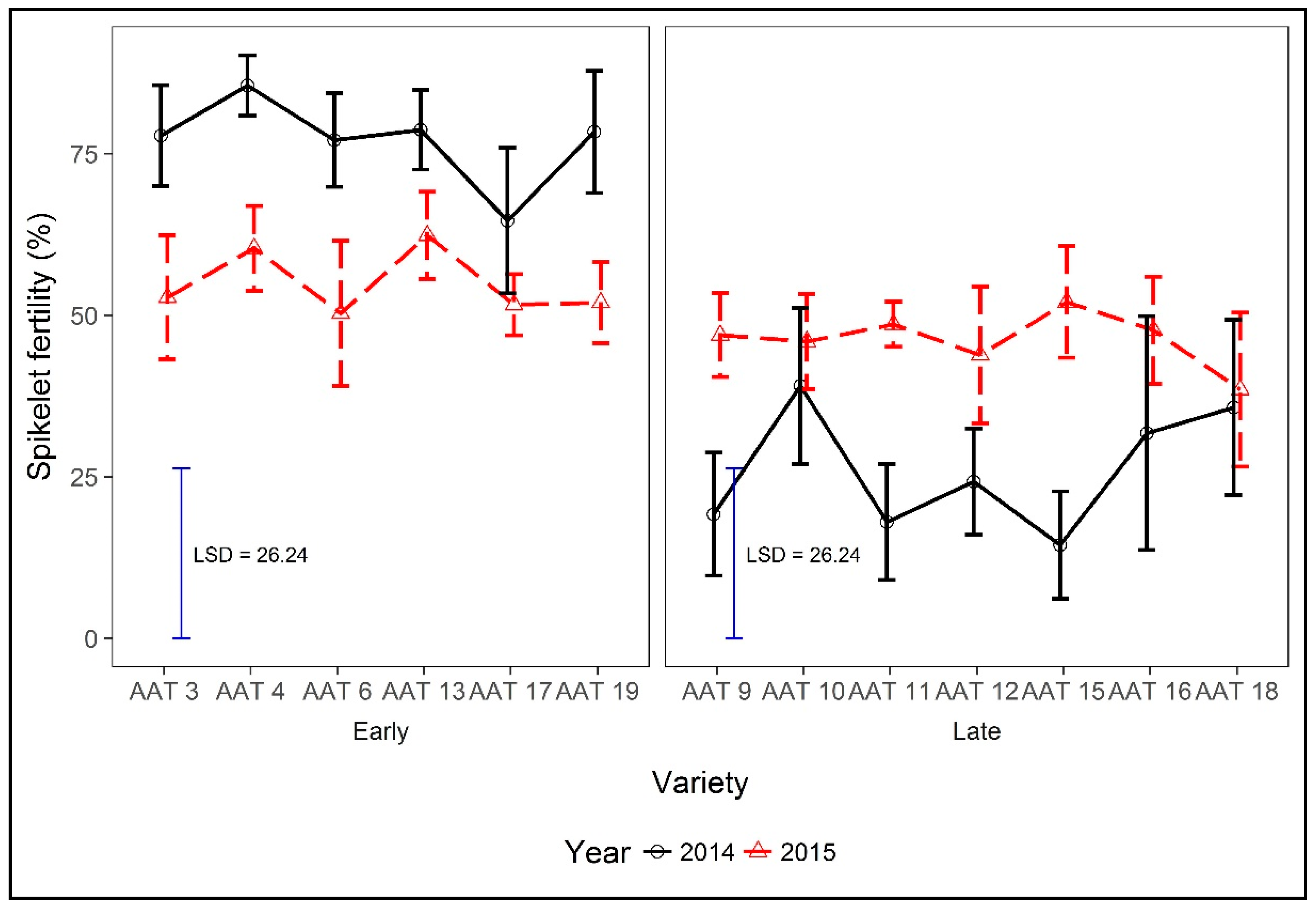
4.4. Effects of Planting Time, Maturity Class, and Strategic Irrigation on Spikelet Fertility, and Other Yield Determining Attributes and the Influence on Yields
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- George, T.; Magbanua, R.; Order, W.; Van Keer, K.; Trébuil, G.; Reoma, V. Upland rice response to phosphorus fertilization in Asia. Agron. J. 2001, 93, 1362–1370. [Google Scholar] [CrossRef]
- Huaqi, W.; Bouman, B.; Zhao, D.; Changgui, W.; Moya, P. Aerobic rice in northern China: Opportunities and challenges. In Water-Wise Rice Production; Bouman, B.A.M., Hengsdijk, H., Hardy, B., Bindraban, P.S., Tuong, T.P., Ladha, J.K., Eds.; International Rice Research Institute: Los Baños, Philippines, 2002; pp. 143–154. [Google Scholar]
- George, T.; Magbanua, R.; Garrity, D.P.; Tubana, B.S.; Quiton, J. Rapid yield loss of rice cropped successively in aerobic soil. Agron. J. 2002, 94, 981–989. [Google Scholar] [CrossRef]
- Datta, S.K.; Krupp, H.K.; Alvarez, E.I.; Modgal, S.C. Water management in flooded tropical rice. In Water Management in Philippine Irrigation Systems: Research and Operations; International Rice Research Institute: Los Baños, Philippines, 1973; pp. 1–18. [Google Scholar]
- Wang, H.; Siopongco, J.; Wade, L.J.; Yamauchi, A. Fractal analysis on root systems of rice plants in response to drought stress. Env. Exp. Bot. 2009, 65, 338–344. [Google Scholar] [CrossRef]
- Long, S.P.; Zhu, X.G.; Naidu, S.L.; Ort, D.R. Can improvement in photosynthesis increase crop yields? Plant Cell Environ. 2006, 29, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Centritto, M.; Lauteri, M.; Monteverdi, M.C.; Serraj, R. Leaf gas exchange, carbon isotope discrimination, and grain yield in contrasting rice genotypes subjected to water deficits during the reproductive stage. J. Exp. Bot. 2009, 60, 2325–2339. [Google Scholar] [CrossRef] [PubMed]
- Dingkuhn, M.; Farquhar, G.; De, D.S.; O’Toole, J.; Datta, S. Discrimination of 13C among upland rices having different water use efficiencies. Aust. J. Agric. Res. 1991, 42, 1123–1131. [Google Scholar] [CrossRef]
- Xu, Y.; This, D.; Pausch, R.C.; Vonhof, W.M.; Coburn, J.R.; Comstock, J.P.; McCouch, S.R. Leaf-level water use efficiency determined by carbon isotope discrimination in rice seedlings: Genetic variation associated with population structure and QTL mapping. Theor. Appl. Genet. 2009, 118, 1065–1081. [Google Scholar] [CrossRef]
- Sinclair, T.R. Effective water use required for improving crop growth rather than transpiration efficiency. Front. Plant Sci. 2018, 9, 1442. [Google Scholar] [CrossRef]
- Serraj, R.; Dimayuga, G.; Gowda, V.; Guan, Y.; He, H.; Impa, S.; Liu, D.; Mabesa, R.; Sellamuthu, R.; Torres, R. Drought-resistant rice: Physiological framework for an integrated research strategy. In Drought Frontiers in Rice: Crop Improvement for Increased Rainfed Production; Serraj, R., Bennett, J., Hardy, B., Eds.; World Scientific Publishing: Los Banos, Philippines, 2008; pp. 139–170. [Google Scholar]
- Chaves, M. Effects of water deficits on carbon assimilation. J. Exp. Bot. 1991, 42, 1–16. [Google Scholar] [CrossRef]
- Lawlor, D.W.; Cornic, G. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant Cell Env. 2002, 25, 275–294. [Google Scholar] [CrossRef] [Green Version]
- Ekanayake, I.J.; De Datta, S.K.; Steponkus, P.L. Spikelet sterility and flowering response of rice to water stress at anthesis. Ann. Bot. 1989, 63, 257–264. [Google Scholar] [CrossRef]
- Murchie, E.H.; Chen, Y.; Hubbart, S.; Peng, S.; Horton, P. Interactions between senescence and leaf orientation determine in situ patterns of photosynthesis and photoinhibition in field-grown rice. Plant Physiol. 1999, 119, 553–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, S. Fundamentals of Rice Crop Science; International Rice Research Institute: Los Baños, Philippines, 1981. [Google Scholar]
- Dingkuhn, M.; De, D.S.; Dorffling, K.; Javellana, C.; Datta, S. Varietal differences in leaf water potential, leaf net CO2 assimilation, conductivity and water use efficiency in upland rice. Aust. J. Agric. Res. 1989, 40, 1183–1192. [Google Scholar] [CrossRef]
- Kashiwagi, J.; Krishnamurthy, L.; Gaur, P.M.; Upadhyaya, H.D.; Varshney, R.K.; Tobita, S. Traits of relevance to improve yield under terminal drought stress in chickpea (Cicer arietinum L.). Field Crops Res. 2013, 145, 88–95. [Google Scholar] [CrossRef] [Green Version]
- Reddy, S.H.; Kambalimath, S.K.; Singhal, R.K.; Chikkakariyappa, M.K.; Muthurajan, R.; Rajanna, M.P.; Sreevathsa, R.; Sevanthi, A.M.; Mohapatra, T.; Sarla, N.; et al. Allele-specific analysis of single parent backcross populationidentifies HOX10 transcription factor as a candidate gene regulating rice root growth. Physiol. Plant 2019, 166, 596–611. [Google Scholar] [CrossRef] [Green Version]
- Ekanayake, I.J.; O’Toole, J.C.; Garrity, D.P.; Masajo, T.M. Inheritance of root characters and their relations to drought resistance in rice. Crop Sci. 1985, 25, 927–933. [Google Scholar] [CrossRef]
- Kumar, N.; Nandwal, A.S.; Waldia, R.S.; Singh, S.; Devi, S.; Sharma, K.D.; Kumar, A. Drought tolerance in chickpea as evaluated by root characteristics, plant water status, membrane integrity and chlorophyll fluorescence techniques. Exp. Agric. 2012, 48, 378–387. [Google Scholar] [CrossRef]
- Hund, A.; Ruta, N.; Liedgens, M. Rooting depth and water use efficiency of tropical maize inbred lines, differing in drought tolerance. Plant Soil 2009, 318, 311–325. [Google Scholar] [CrossRef] [Green Version]
- Hirayama, M.; Wada, Y.; Nemoto, H. Estimation of drought tolerance based on leaf temperature in upland rice breeding. Breed. Sci. 2006, 56, 47–54. [Google Scholar] [CrossRef] [Green Version]
- White, J.W.; Castillo, J.A.; Ehleringer, J. Associations between productivity, root growth and carbon isotope discrimination in Phaseolus vulgaris under water deficit. Funct. Plant Biol. 1990, 17, 189–198. [Google Scholar] [CrossRef]
- Impa, S.; Nadaradjan, S.; Boominathan, P.; Shashidhar, G.; Bindumadhava, H.; Sheshshayee, M. Carbon isotope discrimination accurately reflects variability in WUE measured at a whole plant level in rice. Crop Sci. 2005, 45, 2517–2522. [Google Scholar] [CrossRef]
- Fukai, S.; Inthapan, P. Growth and yield of rice cultivars under sprinkler irrigation in south-eastern Queensland. 1. Effects of sowing time. Aust. J. Exp. Agric. 1988, 28, 237–242. [Google Scholar] [CrossRef]
- Oliver, S.N.; Van Dongen, J.T.; Alfred, S.C.; Mamun, E.A.; Zhao, X.; Saini, H.S.; Fernandes, S.F.; Blanchard, C.L.; Sutton, B.G.; Geigenberger, P.; et al. Cold-induced repression of the rice anther-specific cell wall invertase gene OSINV4 is correlated with sucrose accumulation and pollen sterility. Plant Cell Env. 2005, 28, 1534–1551. [Google Scholar] [CrossRef]
- Challagulla, V.; Bhattarai, S.; Midmore, D. In-vitro vs in-vivo inoculation: Screening for resistance of Australian rice genotypes against blast fungus. Rice Sci. 2015, 22, 132–137. [Google Scholar] [CrossRef] [Green Version]
- Bureau of Meteorology. Climate data online. Bureau of Meteorology, Australian Government, 2016. Available online: https://www.ncdc.noaa.gov/cdo-web/ (accessed on 23 September 2018).
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research, 2nd ed.; John Wiley & Sons: New York, NY, USA, 1984. [Google Scholar]
- Gardner, E.A.; Coughlan, K.J.; Silburn, D.M. Soil water measurement and management on Vertisols in Queensland, Australia. In Management of Vertisols in Sub-Saharan Africa; Jutzi, S.C., Haque, I., McIntire, J., Stares, J.E.S., Eds.; International Livestock Centre for Africa: Nairobi, Kenya, 1988. [Google Scholar]
- Bioversity International, IRRI, WARDA. Descriptors for Wild and Cultivated Rice (Oryza spp.); Bioversity International: Rome, Italy; International Rice Research Institute: Los Baños, Philippines; WARDA, Africa Rice Center: Cotonou, Benin, 2007. [Google Scholar]
- Yoshida, S.; Forno, D.A.; Cock, J.; Gomez, K.A. Laboratory Manual for Physiological Studies of Rice; International Rice Research Institute: Los Baños, Philippines, 1976. [Google Scholar]
- Palaniswamy, K.M.; Gomez, K.A. Length-width method for estimating leaf area of rice. Agron. J. 1974, 66, 430–433. [Google Scholar] [CrossRef]
- Kumar, A.; Guha, A.; Bimolata, W.; Reddy, A.R.; Laha, G.S.; Sundaram, R.; Pandey, M.K.; Ghazi, I.A. Leaf gas exchange physiology in rice genotypes infected with bacterial blight: An attempt to link photosynthesis with disease severity and rice yield. Aust. J. Crop Sci. 2013, 7, 32–39. [Google Scholar]
- Yang, C.; Yang, L.; Yang, Y.; Ouyang, Z. Rice root growth and nutrient uptake as influenced by organic manure in continuously and alternately flooded paddy soils. Agric. Water Manag. 2004, 70, 67–81. [Google Scholar] [CrossRef]
- Fukai, S. Phenology in rainfed lowland rice. Field Crops Res. 1999, 64, 51–60. [Google Scholar] [CrossRef]
- Gunawardena, T.A.; Fukai, S.; Blamey, F.P.C. Low temperature induced spikelet sterility in rice. I. Nitrogen fertilisation and sensitive reproductive period. Aust. J. Agric. Res. 2003, 54, 937–946. [Google Scholar] [CrossRef]
- Farrell, T.; Fukai, S.; Williams, R. Minimising cold damage during reproductive development among temperate rice genotypes. I. Avoiding low temperature with the use of appropriate sowing time and photoperiod-sensitive varieties. Aust. J. Agric. Res. 2006, 57, 75–88. [Google Scholar] [CrossRef]
- Martinez-Eixarch, M.; Ellis, R.H. Temporal sensitivities of rice seed development from spikelet fertility to viable mature seed to extreme-temperature. Crop Sci. 2015, 55, 354–364. [Google Scholar] [CrossRef] [Green Version]
- Devkota, K.; Manschadi, A.; Lamers, J.; Humphreys, E.; Devkota, M.; Egamberdiev, O.; Gupta, R.; Sayre, K.; Vlek, P. Growth and yield of rice (Oryza sativa L.) under resource conservation technologies in the irrigated drylands of Central Asia. Field Crops Res. 2013, 149, 115–126. [Google Scholar] [CrossRef]
- Morris, R.A.; Rumbaoa, F.M., Jr. Rainfall recurrence analysis for extrapolating rice-based cropping patterns. In Proceedings of the A Symposium on the Agrometeorology of the Rice Crop; WMO-IRRI Symposium: Los Baños, Philippines, 1979; pp. 223–233. [Google Scholar]
- Zhao, D.L.; Atlin, G.N.; Amante, M.; Cruz, M.T.S.; Kumar, A. Developing aerobic rice cultivars for water-short irrigated and drought-prone rainfed areas in the tropics. Crop Sci. 2010, 50, 2268–2276. [Google Scholar] [CrossRef]
- Pantuwan, G.; Fukai, S.; Cooper, M.; Rajatasereekul, S.; O’Toole, J.C. Yield response of rice (Oryza sativa L.) genotypes to drought under rainfed lowlands: 2. Selection of drought resistant genotypes. Field Crops Res. 2002, 73, 169–180. [Google Scholar] [CrossRef]
- Song, Y.; Gao, Z.; Luan, W. Interaction between temperature and photoperiod in regulation of flowering time in rice. Sci. China Life Sci. 2012, 55, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Mackill, D.J.; Coffman, W.R.; Garrity, D.P. Rainfed Lowland Rice Improvement; International Rice Research Institute: Los Baños, Philippines, 1996; p. 242. [Google Scholar]
- Tang, L.; Zhu, Y.; Hannaway, D.; Meng, Y.; Liu, L.; Chen, L.; Cao, W. RiceGrow: A rice growth and productivity model. NJAS Wagening. J. Life Sci. 2009, 57, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Stansel, J.W.; Fries, R.E. A conceptual agromet rice model. In Agrometeorology of the Rice Crop; Cowell, R.L., Ed.; WMO/IRRI: Los Baños, Philippines, 1980; pp. 201–212. [Google Scholar]
- Yin, X.; Kropff, M.J.; Goudriaan, J. Changes in temperature sensitivity of development from sowing to flowering in rice. Crop Sci. 1997, 37, 1787–1794. [Google Scholar] [CrossRef]
- Raboin, L.M.; Randriambololona, T.; Radanielina, T.; Ramanantsoanirina, A.; Ahmadi, N.; Dusserre, J. Upland rice varieties for smallholder farming in the cold conditions in Madagascar’s tropical highlands. Field Crops Res. 2014, 169, 11–20. [Google Scholar] [CrossRef]
- Tao, H.; Brueck, H.; Dittert, K.; Kreye, C.; Lin, S.; Sattelmacher, B. Growth and yield formation of rice (Oryza sativa L.) in the water-saving ground cover rice production system (GCRPS). Field Crops Res. 2006, 95, 1–12. [Google Scholar] [CrossRef]
- Yue, B.; Xue, W.Y.; Luo, L.J.; Xing, Y.Z. QTL analysis for flag leaf characteristics and their relationships with yield and yield traits in rice. Acta Genet. Sin. 2006, 33, 824–832. [Google Scholar] [CrossRef]
- Cao, S.Q.; Zhai, H.Q.; Yang, T.N.; Zhang, R.X.; Kuang, T.Y. Studies on photosynthetic rate and function duration of rice germplasm. Chin. J. Rice Sci. 2001, 15, 29–34. [Google Scholar]
- Chen, W.F.; Xu, Z.J.; Zhang, B.L. Physiological Bases of Super High Yield Breeding in Rice; Liao Ning Science and Technology Publishing Company: Shenyang, China, 1995. [Google Scholar]
- Lauteri, M.; Haworth, M.; Serraj, R.; Monteverde, M.C.; Centritto, M. Photosynthetic diffusional constraints affect yield in drought stressed rice cultivars during flowering. PLoS ONE 2015, 10, e0117631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teng, S.; Qian, Q.; Zeng, D.; Kunihiro, Y.; Fujimoto, K.; Huang, D.; Zhu, L. QTL analysis of leaf photosynthetic rate and related physiological traits in rice (Oryza sativa L.). Euphytica 2004, 135, 1–7. [Google Scholar] [CrossRef]
- Gu, J.; Yin, X.; Stomph, T.-J.; Wang, H.; Struik, P. Physiological basis of genetic variation in leaf photosynthesis among rice (Oryza sativa L.) introgression lines under drought and well-watered conditions. J. Exp. Bot. 2012, 63, 695–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henry, A. IRRI’s drought stress research in rice with emphasis on roots: Accomplishments over the last 50 years. Plant Root 2013, 7, 92–106. [Google Scholar] [CrossRef] [Green Version]
- Chang, T.T.; Vergara, B.S. Varietal diversity and morpho-agronomic characteristics of upland rice. In Major Research in Upland Rice; International Rice Research Institute: Los Baños, Philippines, 1975; pp. 72–90. [Google Scholar]
- Araki, H.; Iijima, M. Stable isotope analysis of water extraction from subsoil in upland rice (Oryza sativa L.) as affected by drought and soil compaction. Plant Soil 2005, 270, 147–157. [Google Scholar] [CrossRef]
- Niones, J.M.; Suralta, R.R.; Inuka, Y.; Yamauchi, A. Field evaluation on functional roles of root plastic responses on dry matter production and grain yield of rice under cycles of transient soil moisture stresses using chromosome segment substitution lines. Plant Soil 2012, 359, 107–120. [Google Scholar] [CrossRef]
- Lilley, J.M.; Fukai, S. Effect of timing and severity of water deficit on four diverse rice cultivars I. Rooting pattern and soil water extraction. Field Crops Res. 1994, 37, 205–213. [Google Scholar] [CrossRef]
- Zheng, C.; Zhang, Z.; Hao, S.; Chen, W.; Pan, Y.; Wang, Z. Agronomic growth performance of super rice under water-saving irrigation methods with different water-controlled thresholds in different growth stages. Agronomy 2020, 10, 239. [Google Scholar] [CrossRef] [Green Version]
- Gunawardena, T.A.; Fukai, S. The interaction of nitrogen application and temperature during reproductive stage on spikelet sterility in field-grown rice. Aust. J. Agric. Res. 2005, 56, 625–636. [Google Scholar] [CrossRef]
- Cruz, R.; O’Toole, J. Dryland rice response to an irrigation gradient at flowering stage. Agron. J. 1984, 76, 178–183. [Google Scholar] [CrossRef]
- Jongdee, B.; Fukai, S.; Cooper, M. Leaf water potential and osmotic adjustment as physiological traits to improve drought tolerance in rice. Field Crops Res. 2002, 76, 153–163. [Google Scholar] [CrossRef]
- Saini, H.S.; Lalonde, S. Injuries to reproductive development under water stress, and their consequences for crop productivity. J. Crop. Prod. 1997, 1, 223–248. [Google Scholar] [CrossRef]
- Lanceras, J.C.; Pantuwan, G.; Jongdee, B.; Toojinda, T. Quantitative trait loci associated with drought tolerance at reproductive stage in rice. Plant Physiol. 2004, 135, 384–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujita, K.; Coronel, V.P.; Yoshida, S. Grain filling characteristics of rice varieties (Oryza sativa L.) differing in grain size under controlled environmental conditions. Soil Sci. Plant Nutr. 1984, 30, 445–454. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.R.; Wolfrum, E.J.; Vick, P.; Ismail, A.; Greenberg, A.J.; McCrouch, S.R. Robust phenotyping strategies for evaluation of stem non-structural carbohydrates (NSC) in rice. J. Exp. Bot. 2016, 67, 6125–6138. [Google Scholar] [CrossRef] [Green Version]
- Bouman, B.A.M.; Peng, S.; Castañeda, A.R.; Visperas, R.M. Yield and water use of irrigated tropical aerobic rice systems. Agric. Water Manag. 2005, 74, 87–105. [Google Scholar] [CrossRef]
- Kato, Y.; Okami, M.; Katsura, K. Yield potential and water use efficiency of aerobic rice (Oryza sativa L.) in Japan. Field Crops Res. 2009, 113, 328–334. [Google Scholar] [CrossRef]
- Stevens, G.; Vories, E.; Heiser, J.; Rhine, M.; Dunn, D. Experimentation on cultivation of rice irrigated with a center pivot system. In Irrigation Systems and Practices in Challenging Environments; Lee, T.S., Ed.; INTECH Open Access Publisher: Rijeka, Croatia, 2012; pp. 134–254. [Google Scholar]
- Shi, Y.; Shen, Q.; Mao, Z.; Li, W. Biological response of rice crop cultivated on upland soil condition and the effect of mulching on it. Plant Nutr. Fertil. Sci. 2001, 7, 271–277. [Google Scholar]
- Sudhir, Y.; Gill, G.; Humphreys, E.; Kukal, S.S.; Walia, U.S. Effect of water management on dry seeded and puddled transplanted rice. Part 1: Crop performance. Field Crops Res. 2011, 120, 112–122. [Google Scholar] [CrossRef]
- Matsunami, M.; Matsunami, T.; Kokubun, M. Growth and yield of new rice for Africa (NERICAs) under different ecosystems and nitrogen levels. Plant Prod. Sci. 2009, 12, 381–389. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Varieties | Maturity | 2014 | 2015 | Mean Yield (t/ha) over Two Seasons * | |||||
|---|---|---|---|---|---|---|---|---|---|
| Rainfed | Strategic Irrigation | Mean | Rainfed | Strategic Irrigation | Mean | ||||
| AAT 9 | Late | 0.10 | 3.03 | 1.57 | 0.82 | 3.81 | 2.31 | 1.94 abc | |
| AAT 10 | Late | 0.84 | 2.38 | 1.61 | 1.14 | 4.38 | 2.76 | 2.19 c | |
| AAT 11 | Late | 0.49 | 3.08 | 1.79 | 0.61 | 3.90 | 2.25 | 2.02 bc | |
| AAT 12 | Late | 0.26 | 2.81 | 1.53 | 0.79 | 3.66 | 2.23 | 1.88 abc | |
| AAT 15 | Late | 0.11 | 1.34 | 0.72 | 0.24 | 4.36 | 2.30 | 1.51 ab | |
| AAT 16 | Late | 0.12 | 2.24 | 1.18 | 0.59 | 4.06 | 2.33 | 1.75 abc | |
| AAT 18 | Late | 0.10 | 1.81 | 0.96 | 0.42 | 3.32 | 1.87 | 1.41 a | |
| AAT 3 | Early | 2.12 | 4.20 | 3.16 | 1.94 | 3.90 | 2.92 | 3.04 d | |
| AAT 4 | Early | 3.85 | 3.48 | 3.67 | 2.10 | 5.23 | 3.67 | 3.67 e | |
| AAT 6 | Early | 2.71 | 4.48 | 3.60 | 2.72 | 4.62 | 3.67 | 3.63 e | |
| AAT 13 | Early | 2.19 | 4.68 | 3.44 | 2.02 | 4.58 | 3.30 | 3.37 de | |
| AAT 17 | Early | 2.99 | 4.57 | 3.78 | 1.94 | 4.06 | 3.00 | 3.39 de | |
| AAT 19 | Early | 2.71 | 3.64 | 3.18 | 2.64 | 4.95 | 3.80 | 3.49 de | |
| Average | 1.43 | 3.21 | 2.32 | 1.38 | 4.22 | 2.80 | 2.56 | ||
| p-value and LSD0.05 | |||||||||
| Year (Y) | 0.107 (0.735) | Y×V | 0.002 (0.756) | Y×I×V | 0.109 (1.147) | ||||
| Variety (V) | <0.001 (0.485) | V×I | 0.178 (0.815) | ||||||
| Irrigation (I) | 0.006 (0.747) | Y×I | 0.093 (0.676) | ||||||
| Varieties | Maturity | Year 2014 | Year 2015 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Rainfed | Strategic Irrigation | Mean | Rainfed | Strategic Irrigation | Mean | ||||
| AAT 9 | Late | 2.88 | 3.24 | 3.06 | 1.37 | 2.60 | 1.98 | ||
| AAT 10 | Late | 3.79 | 3.57 | 3.68 | 1.24 | 3.05 | 2.14 | ||
| AAT 11 | Late | 3.07 | 3.89 | 3.48 | 1.30 | 2.82 | 2.06 | ||
| AAT 12 | Late | 3.34 | 4.45 | 3.89 | 1.85 | 3.07 | 2.46 | ||
| AAT 15 | Late | 2.58 | 2.67 | 2.63 | 1.37 | 2.51 | 1.94 | ||
| AAT 16 | Late | 2.84 | 3.64 | 3.24 | 1.08 | 2.22 | 1.65 | ||
| AAT 18 | Late | 2.68 | 3.30 | 2.99 | 1.23 | 2.71 | 1.97 | ||
| AAT 3 | Early | 1.63 | 2.43 | 2.03 | 1.73 | 3.99 | 2.86 | ||
| AAT 4 | Early | 1.97 | 1.79 | 1.88 | 1.84 | 3.94 | 2.89 | ||
| AAT 6 | Early | 1.86 | 2.74 | 2.30 | 1.47 | 4.89 | 3.18 | ||
| AAT 13 | Early | 2.35 | 3.15 | 2.75 | 1.90 | 4.30 | 3.10 | ||
| AAT 17 | Early | 2.89 | 3.23 | 3.06 | 1.91 | 4.19 | 3.05 | ||
| AAT 19 | Early | 2.62 | 2.45 | 2.54 | 1.79 | 4.69 | 3.24 | ||
| Average | 2.65 | 3.12 | 2.89 | 1.54 | 3.46 | 2.50 | |||
| p-value (LSD0.05) | |||||||||
| Year (Y) | 0.269 (1.089) | Y × V | <0.001 (0.942) | Y × I × V | 0.201 (1.196) | ||||
| Variety (V) | 0.036 (0.542) | V × I | 0.302 (0.801) | ||||||
| Irrigation (I) | 0.020 (0.731) | Y × I | 0.051 (0.897) | ||||||
| Year | DAS | Rainfed | Strategic Irrigation | LSD (p = 0.05) |
|---|---|---|---|---|
| 2014 | 83 | 13.2 | 13.2 | ns |
| 103 | 7.7 | 8.2 | ns | |
| 133 | 3.3 | 6.0 | 0.94 | |
| 2015 | 51 | 7.9 | 7.9 | ns |
| 82 | 14.7 | 14.2 | ns | |
| 95 | 3.8 | 11.7 | 0.55 | |
| 110 | 6.7 | 6.1 | ns |
| Soil Depth (cm) | ||||||
|---|---|---|---|---|---|---|
| Root Parameters | Source of Variation | Total | 0–15 | 15–30 | 30–60 | 60–100 |
| Root Length Density (RLD) (mm/cm3) | Irrigation (I) | 0.341 | 0.042 | 0.713 | 0.419 | 0.720 |
| Variety (V) | 0.190 | 0.109 | 0.598 | <0.001 | 0.497 | |
| Root Diameter (RD) (mm) | Irrigation (I) | 0.872 | 0.200 | 0.750 | 0.698 | 0.394 |
| Variety (V) | 0.847 | 0.301 | 0.245 | 0.036 | 0.483 | |
| Root Weight Density (RWD) (mg/cm3) | Irrigation (I) | 0.210 | 0.176 | 0.740 | 0.825 | 0.401 |
| Variety (V) | 0.195 | 0.270 | 0.173 | 0.002 | 0.840 | |
| Varieties | Maturity | Year 2014 | Year 2015 | Two-Season Mean * | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Rainfed | Strategic Irrigation | Mean | Rainfed | Strategic Irrigation | Mean | |||||
| AAT 9 | Late | 220 | 150 | 185 | 101 | 187 | 144 | 165 a | ||
| AAT 10 | Late | 145 | 179 | 162 | 88 | 172 | 130 | 146 a | ||
| AAT 11 | Late | 169 | 189 | 179 | 118 | 193 | 156 | 168 a | ||
| AAT 12 | Late | 142 | 189 | 165 | 102 | 170 | 136 | 151 a | ||
| AAT 15 | Late | 153 | 169 | 161 | 108 | 154 | 131 | 146 a | ||
| AAT 16 | Late | 170 | 175 | 172 | 115 | 137 | 126 | 149 a | ||
| AAT 18 | Late | 204 | 222 | 213 | 74 | 153 | 114 | 164 a | ||
| AAT 3 | Early | 100 | 129 | 114 | 106 | 123 | 115 | 115 b | ||
| AAT 4 | Early | 100 | 128 | 114 | 101 | 115 | 108 | 111 b | ||
| AAT 6 | Early | 91 | 109 | 100 | 103 | 117 | 110 | 105 b | ||
| AAT 13 | Early | 107 | 125 | 116 | 106 | 109 | 107 | 112 b | ||
| AAT 17 | Early | 90 | 110 | 100 | 102 | 123 | 113 | 107 b | ||
| AAT 19 | Early | 81 | 116 | 98 | 105 | 124 | 114 | 106 b | ||
| Average | 136 | 153 | 145 | 102 | 144 | 123 | 134 | |||
| p-value (LSD 0.05) | ||||||||||
| Year (Y) | 0.045 (20.03) | Y × V | 0.004 (33.85) | Y × I × V | 0.368 (55.61) | |||||
| Variety (V) | <0.001 (23.99) | V × I | 0.796 (39.11) | |||||||
| Irrigation (I) | 0.033 (23.48) | Y × I | 0.145 (20.11) | |||||||
| Varieties | Maturity | Year 2014 | Year 2015 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Rainfed | Strategic Irrigation | Mean | Rainfed | Strategic Irrigation | Mean | ||||
| AAT 9 | Late | 18.7 | 18.1 | 18.4 | 19.5 | 21.4 | 20.4 | ||
| AAT 10 | Late | 21.3 | 23.2 | 22.3 | 18.8 | 21.7 | 20.3 | ||
| AAT 11 | Late | 18.2 | 22.5 | 20.3 | 18.8 | 22.3 | 20.6 | ||
| AAT 12 | Late | 20.4 | 21.3 | 20.8 | 21.2 | 21.7 | 21.5 | ||
| AAT 15 | Late | 26.2 | 26.3 | 26.3 | 24.9 | 29.0 | 27.0 | ||
| AAT 16 | Late | 15.8 | 19.9 | 17.8 | 18.3 | 20.2 | 19.2 | ||
| AAT 18 | Late | 19.4 | 20.7 | 20.0 | 17.7 | 21.0 | 19.3 | ||
| AAT 3 | Early | 26.7 | 26.5 | 26.6 | 26.3 | 29.5 | 27.9 | ||
| AAT 4 | Early | 29.4 | 27.0 | 28.2 | 25.9 | 28.1 | 27.0 | ||
| AAT 6 | Early | 24.7 | 27.9 | 26.3 | 26.0 | 28.7 | 27.3 | ||
| AAT 13 | Early | 26.2 | 26.3 | 26.3 | 24.9 | 29.0 | 27.0 | ||
| AAT 17 | Early | 27.3 | 24.6 | 25.9 | 25.4 | 28.2 | 26.8 | ||
| AAT 19 | Early | 29.0 | 26.0 | 27.5 | 25.7 | 29.0 | 27.4 | ||
| Average | 22.6 | 23.5 | 23.1 | 22.1 | 24.6 | 23.4 | |||
| p-value (LSD0.05) | |||||||||
| Year (Y) | 0.428 (1.426) | Y × V | 7.66 (2.803) | Y × I × V | 0.006 (3.232) | ||||
| Variety (V) | <0.001 (2.009) | V × I | 0.033 (2.294) | ||||||
| Irrigation (I) | 0.009 (0.723) | Y × I | 0.04 (1.191) | ||||||
| Flowering | 2014 (January–May 2014) | 2015 (November 2014–March 2015) | ||||
|---|---|---|---|---|---|---|
| Sowing to Flowering GDD | Ave pp | No. of days for Flowering | Sowing to Flowering GDD | Ave pp | No. of Days for Flowering | |
| Early (japonica) | 1225 | 12.55 | 76 | 1554 | 13.40 | 87 |
| Late (indica) | 1565 | 12.26 | 102 | 1695 | 13.35 | 96 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silwal, S.; Bhattarai, S.P.; Midmore, D.J. Aerobic Rice with or without Strategic Irrigation in the Subtropics. Agronomy 2020, 10, 1831. https://doi.org/10.3390/agronomy10111831
Silwal S, Bhattarai SP, Midmore DJ. Aerobic Rice with or without Strategic Irrigation in the Subtropics. Agronomy. 2020; 10(11):1831. https://doi.org/10.3390/agronomy10111831
Chicago/Turabian StyleSilwal, Sachesh, Surya P. Bhattarai, and David J. Midmore. 2020. "Aerobic Rice with or without Strategic Irrigation in the Subtropics" Agronomy 10, no. 11: 1831. https://doi.org/10.3390/agronomy10111831
APA StyleSilwal, S., Bhattarai, S. P., & Midmore, D. J. (2020). Aerobic Rice with or without Strategic Irrigation in the Subtropics. Agronomy, 10(11), 1831. https://doi.org/10.3390/agronomy10111831