Comparison of the Yield and Chemical Composition of Eleven Timothy (Phleum pratense L.) Genotypes under Three Locations in Poland

 ,
,  , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Field
2.2. Statistical Analysis
3. Results
3.1. Weather Conditions
3.2. Yield Assessment
3.3. Quality Analysis
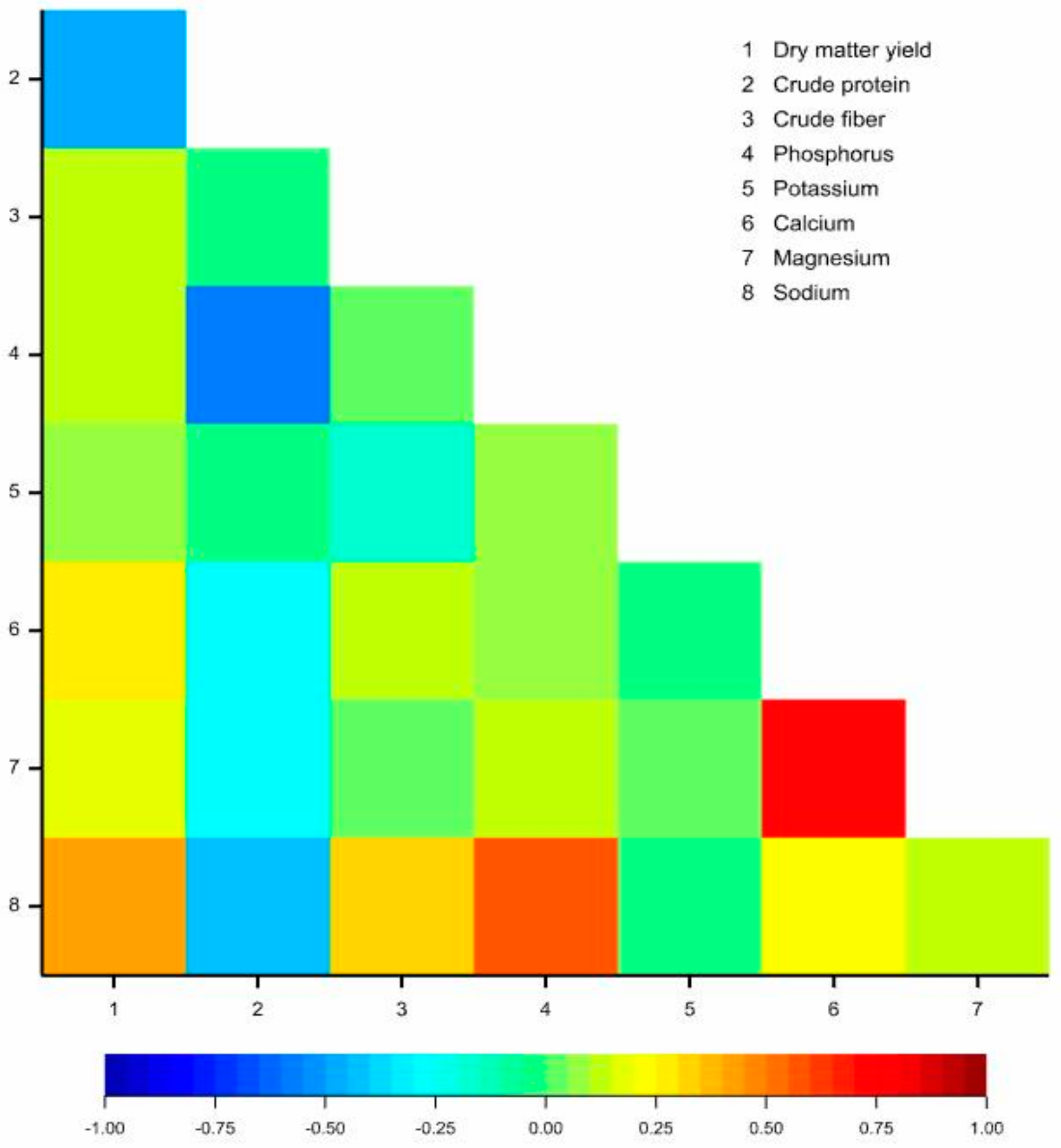
3.4. Correlations between Observed Traits
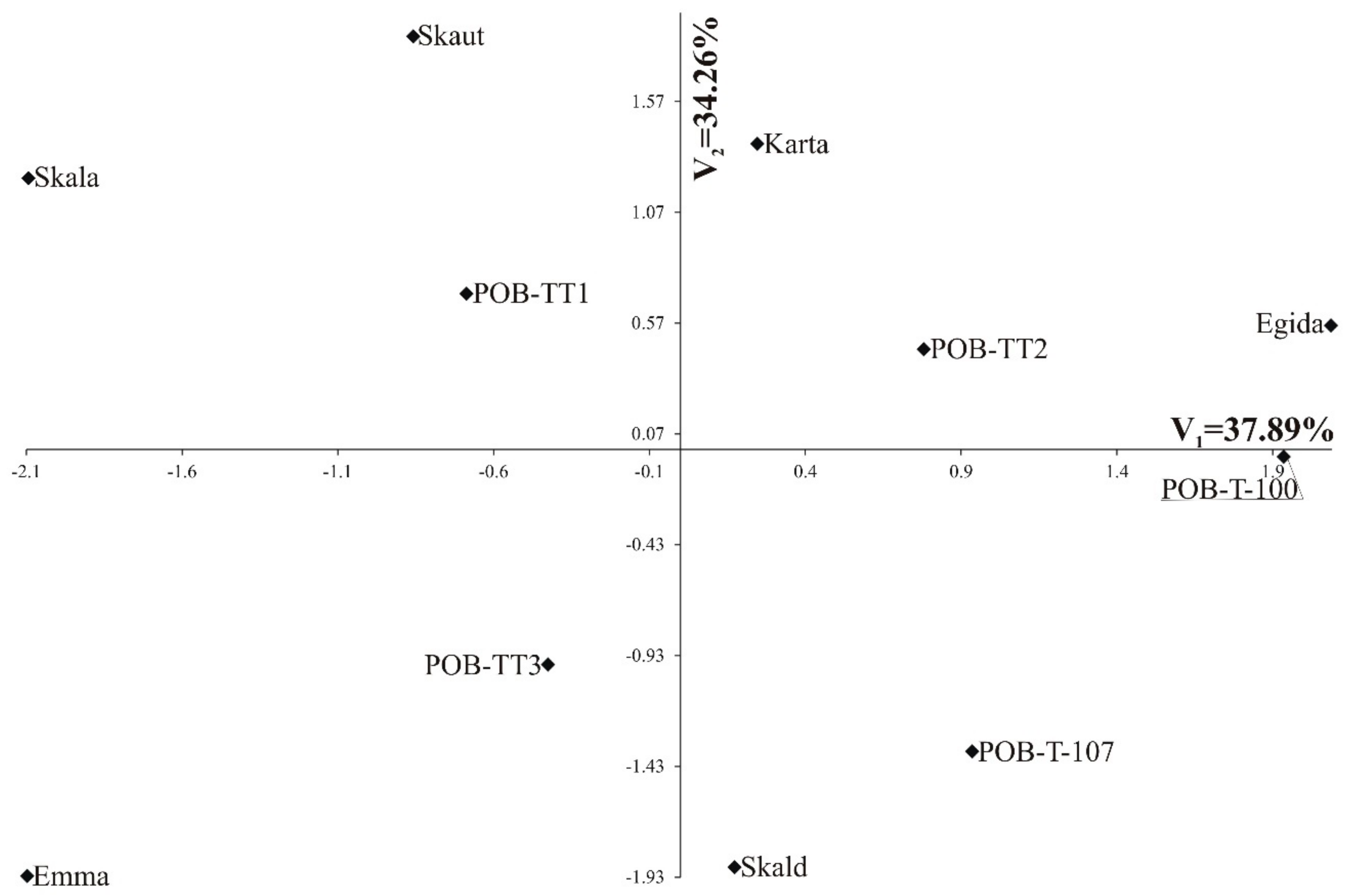
3.5. Multivariate Relationships
4. Discussion
5. Future Breeding Strategies
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Larsen, A. Breeding winter hardy grasses. Euphytica 1994, 77, 231–237. [Google Scholar] [CrossRef]
- Bélanger, G.; Michaud, R.; Jefferson, P.G.; Tremblay, G.F.; Brégard, A. Improving the nutritive value of timothy through management and breeding. Can. J. Plant Sci. 2001, 81, 577–585. [Google Scholar] [CrossRef]
- Berg, C.C.; McElroy, A.R.; Kunelius, H.T. Timothy. In Cool-Season Forage Grasses; Moser, L.E., Buxton, D.R., Casler, M.D., Eds.; ASA: Brooklyn, NY, USA; CSSA: Hong Kong; SSSA: Madison, WI, USA, 1996; pp. 643–664. [Google Scholar]
- Bélanger, G.; McQueen, R.E. Digestibility and cell wall concentration of early- and late-maturing timothy (Phleum pratense L.) cultivars. Can. J. Plant Sci. 1996, 76, 107–112. [Google Scholar] [CrossRef][Green Version]
- Bélanger, G.; McQueen, R.E. Analysis of the nutritive value of timothy grown with varying N nutrition. Grass Forage Sci. 1998, 53, 109–119. [Google Scholar] [CrossRef]
- Casler, M.D.; Vogel, K.P. Accomplishments and impact from breeding for increased forage nutritional value. Crop Sci. 1999, 39, 12–20. [Google Scholar] [CrossRef]
- Brégard, A.; Bélanger, G.; Michaud, R.; Tremblay, G.F. Biomass partitioning, forage nutritive value, and yield of contrasting genotypes of timothy. Crop Sci. 2001, 41, 1212–1219. [Google Scholar] [CrossRef]
- Claessens, A.; Michaud, R.; Bélanger, G.; Mather, D.E. Responses to divergent phenotypic selection for fiber traits in timothy. Crop Sci. 2005, 45, 1017–1022. [Google Scholar] [CrossRef]
- Claessens, A.; Michaud, R.; Bélanger, G.; Mather, D.E. Leaf and stem characteristics of timothy plants divergently selected for the ration of lignin to cellulose. Crop Sci. 2005, 45, 2425–2429. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 18th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 2006. [Google Scholar]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Rencher, A.C. Interpretation of canonical discriminant functions, canonical variates, and principal components. Am. Stat. 1992, 46, 217–225. [Google Scholar]
- Seidler-Łożykowska, K.; Bocianowski, J. Evaluation of variability of morphological traits of selected caraway (Carum carvi L.) genotypes. Ind. Crops Prod. 2012, 35, 140–145. [Google Scholar] [CrossRef]
- Mahalanobis, P.C. On the generalized distance in statistics. Proc. Natl. Inst. Sci. India 1936, 12, 49–55. [Google Scholar]
- Genstat, 18th ed.; v.18.2.0.18409; VSN International Ltd.: Hemel Hempstead, UK, 2016.
- Thorvaldsson, G.; Andersson, S. Variations in timothy dry matter yield and nutritional value as affected by harvest date, nitrogen fertilization, year and location in Northern Sweden. Acta Agric. Scand. 1986, 36, 367–385. [Google Scholar] [CrossRef]
- Thorvaldsson, G.; Björnsson, H. The effects of weather on growth, crude protein and digestibility of some grass species in Iceland. Icel. Agric. Sci. 1990, 4, 19–36. [Google Scholar]
- Michaud, R.; Tremblay, G.F.; Bélanger, G. Protein fractions and N fertilization in timothy. Czech J. Genet. Plant Breed. 2003, 39, 279–281. [Google Scholar]
- De Araüjo, M.R.A.; Coulman, B.E.; Rakow, G. Genetic variation, heritability and progeny testing in meadow bromegrass. Plant Breed. 2002, 121, 417–424. [Google Scholar] [CrossRef]
- Jankowski, K.; Jodełka, J. Analysis of the impact of climatic factors on the yield and nutritional value of four legume-grass mixtures. Sci. Noteb. Siedlce Agric. 2000, 57, 93–101. [Google Scholar]
- Falkowski, M.; Kukułka, I.; Kozłowski, S. Chemical Properties of Meadow Plants; Academy of Agriculture Poznań: Poznań, Poland, 2000; p. 132. [Google Scholar]
- Lahuta, L.B.; Ciak, M.; Rybiński, W.; Bocianowski, J.; Börner, A. Diversity of the composition and content of soluble carbohydrates in seeds of the genus Vicia (Leguminosae). Genet. Resour. Crop Eviron. 2018, 65, 541–554. [Google Scholar] [CrossRef]
- Niemann, J.; Bocianowski, J.; Wojciechowski, A. Effects of genotype and environment on seed quality traits variability in interspecific cross-derived Brassica lines. Euphytica 2018, 214, 193. [Google Scholar] [CrossRef]
- Bocianowski, J.; Majchrzak, L. Analysis of effects of cover crop and tillage method combinations on the phenotypic traits of spring wheat (Triticum aestivum L.) using multivariate methods. Appl. Ecol. Environ. Res. 2019, 17, 15267–15276. [Google Scholar] [CrossRef]
- Shamsuddin, A.K.M. Genetic diversity in relation to heterosis and combining ability in spring wheat. Theor. Appl. Genet. 1985, 70, 306–308. [Google Scholar] [CrossRef] [PubMed]
- Seidler-Łożykowska, K.; Bocianowski, J.; Król, D. The evaluation of the variability of morphological and chemical traits of the selected lemon balm (Melissa officinalis L.) genotypes. Ind. Crops Prod. 2013, 49, 515–520. [Google Scholar] [CrossRef]
- Nowosad, K.; Liersch, A.; Popławska, W.; Bocianowski, J. Genotype by environment interaction for seed yield in rapeseed (Brassica napus L.) using additive main effects and multiplicative interaction model. Euphytica 2016, 208, 187–194. [Google Scholar] [CrossRef]
- Bocianowski, J.; Nowosad, K.; Szulc, P. Soil tillage methods by years interaction for harvest index of maize (Zea mays L.) using additive main effects and multiplicative interaction model. Acta Agric. Scand. Sect. B 2019, 69, 75–81. [Google Scholar] [CrossRef]
- Bocianowski, J.; Szulc, P.; Nowosad, K. Soil tillage methods by years interaction for dry matter of plant yield of maize (Zea mays L.) using additive main effects and multiplicative interaction model. J. Integr. Agric. 2018, 17, 2836–2839. [Google Scholar] [CrossRef]
- Wrońska-Pilarek, D.; Szkudlarz, P.; Bocianowski, J. Systematic importance of morphological features of pollen grains of species from Erica (Ericaceae) genus. PLoS ONE 2018, 13, e0204557. [Google Scholar] [CrossRef]
- Bocianowski, J.; Nowosad, K.; Bujak, H.; Łuczkiewicz, T.; Piesik, D. Evaluation of the breeding value of the spring oilseed rape (Brassica napus L.) inbred lines based on a multi-trait analysis. Indian J. Genet. Plant Breed. 2016, 76, 284–289. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | Nieznanice | Polanowice | Palikije |
|---|---|---|---|
| Soil type | brown | degraded chernozem derived from loess | brown |
| Complex agricultural soil suitability | rye, good | wheat, very good | wheat, good |
| Class of soil valuation | IIIb | I | IIIa |
| soil pH in KCl | 6.0 | 7.2 | 6.5 |
| Trait | Cut | Dry Matter Yield | Crude Protein | Crude Fiber | Phosphorus | Potassium | Calcium | Magnesium |
|---|---|---|---|---|---|---|---|---|
| Crude protein | I | −0.29 * | ||||||
| II | 0.28 * | |||||||
| III | −0.51 *** | |||||||
| Crude fiber | I | 0.02 | −0.44 *** | |||||
| II | 0.62 *** | 0.40 *** | ||||||
| III | 0.14 | −0.06 | ||||||
| Phosphorus | I | 0.27 * | −0.44 *** | 0.16 | ||||
| II | −0.39 ** | −0.61 *** | −0.33 ** | |||||
| III | 0.15 | −0.57 *** | 0.01 | |||||
| Potassium | I | −0.09 | −0.22 | 0.07 | −0.05 | |||
| II | −0.18 | −0.22 | −0.10 | 0.14 | ||||
| III | 0.13 | −0.05 | −0.41 *** | 0.13 | ||||
| Calcium | I | 0.28 * | −0.19 | 0.03 | 0.34 ** | −0.34 ** | ||
| II | 0.15 | −0.19 | 0.18 | −0.04 | −0.22 | |||
| III | 0.33 ** | −0.34 ** | 0.06 | 0.07 | −0.31 * | |||
| Magnesium | I | 0.41 *** | −0.14 | −0.01 | 0.16 | −0.16 | 0.71 *** | |
| II | −0.05 | −0.23 | −0.04 | 0.20 | −0.09 | 0.68 *** | ||
| III | 0.20 | −0.36 ** | −0.03 | 0.17 | −0.15 | 0.77 *** | ||
| Sodium | I | 0.23 | −0.75 *** | 0.48 *** | 0.53 *** | 0.26 * | 0.29 * | 0.29 * |
| II | 0.21 | −0.41 *** | 0.38 ** | 0.48 *** | 0.22 | 0.24 | 0.10 | |
| III | 0.48 *** | −0.46 *** | 0.34 ** | 0.62 *** | 0.00 | 0.24 | 0.12 |
| Cut | I | II | III | |||
|---|---|---|---|---|---|---|
| Trait | V1 | V2 | V1 | V2 | V1 | V2 |
| Dry matter yield | 0.014 | 0.781 ** | 0.224 | 0.079 | −0.08 | −0.359 |
| Crude protein | −0.576 | −0.028 | 0.434 | 0.055 | −0.524 | 0.202 |
| Crude fiber | 0.091 | 0.146 | −0.006 | −0.302 | 0.923 *** | 0.222 |
| Phosphorus | 0.472 | 0.694 * | 0.112 | 0.484 | 0.043 | 0.075 |
| Potassium | 0.102 | −0.837 ** | −0.319 | −0.765 ** | −0.491 | 0.294 |
| Calcium | −0.685 * | 0.623 * | 0.685 * | 0.336 | −0.029 | −0.816 ** |
| Magnesium | −0.901 *** | 0.337 | 0.980 *** | −0.067 | 0.134 | −0.974 *** |
| Sodium | −0.712 * | −0.48 | −0.249 | −0.468 | −0.363 | 0.169 |
| Percentage of elucidated multivariate variability | 46.63 | 31.23 | 53.44 | 21.89 | 37.89 | 34.26 |
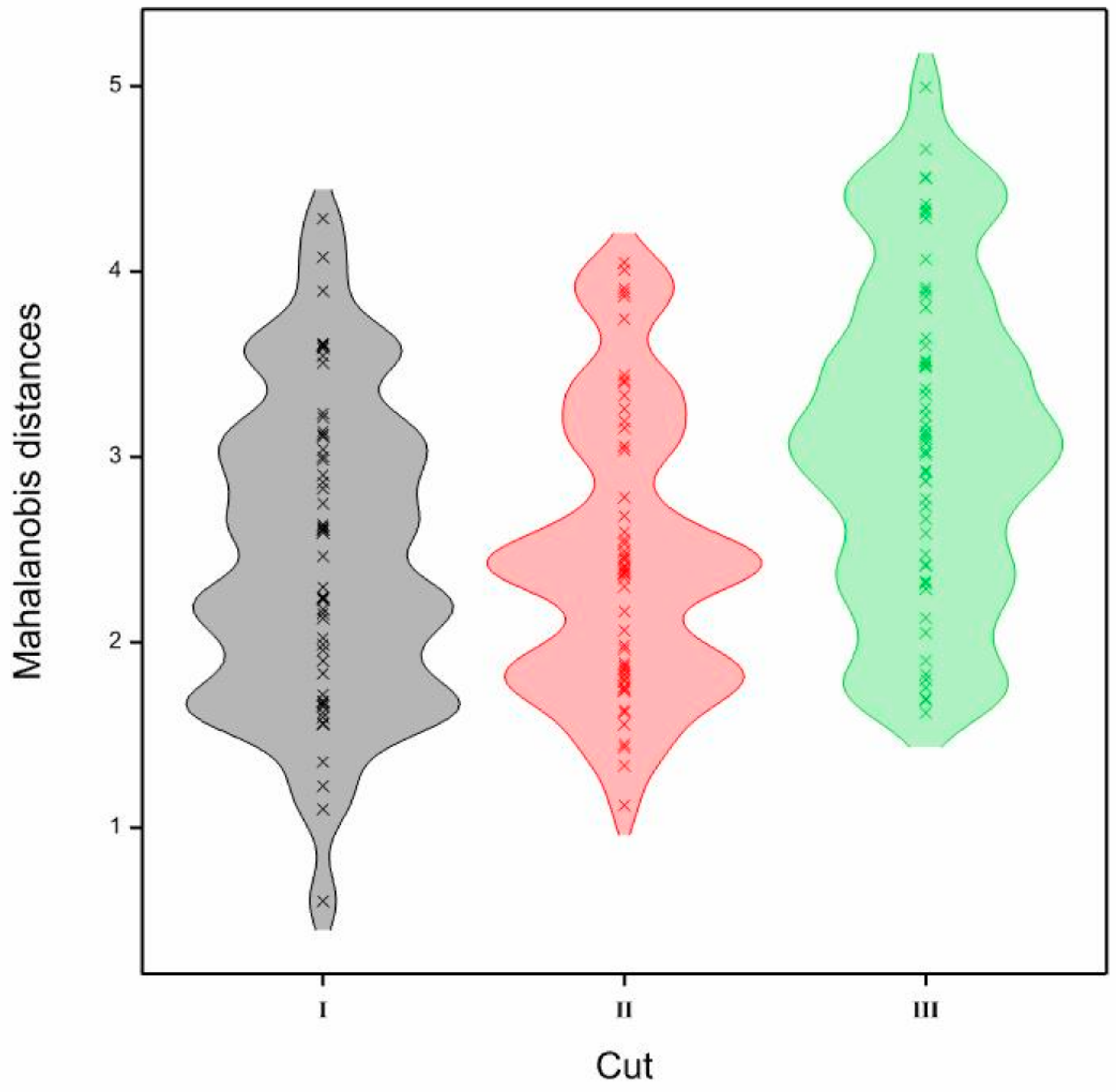
| Genotype | Cut | POB-TT1 | POB-TT2 | POB-TT3 | POB-T-100 | POB-T-107 | Skald | Egida | Emma | Skala | Skaut |
|---|---|---|---|---|---|---|---|---|---|---|---|
| POB-TT2 | I | 0.604 | |||||||||
| II | 1.121 | ||||||||||
| III | 1.902 | ||||||||||
| POB-TT3 | I | 1.558 | 1.683 | ||||||||
| II | 1.749 | 1.451 | |||||||||
| III | 1.827 | 2.052 | |||||||||
| POB-T-100 | I | 1.958 | 1.677 | 2.246 | |||||||
| II | 2.452 | 1.845 | 1.791 | ||||||||
| III | 2.918 | 1.696 | 2.733 | ||||||||
| POB-T-107 | I | 1.831 | 1.62 | 1.355 | 2.023 | ||||||
| II | 3.038 | 2.403 | 1.622 | 2.783 | |||||||
| III | 3.522 | 2.419 | 2.475 | 3.099 | |||||||
| Skald | I | 3.122 | 3.214 | 2.464 | 3.605 | 2.244 | |||||
| II | 3.443 | 3.058 | 2.064 | 3.26 | 1.739 | ||||||
| III | 3.225 | 3.265 | 2.132 | 3.645 | 2.288 | ||||||
| Egida | I | 1.596 | 1.717 | 1.1 | 2.613 | 1.901 | 2.606 | ||||
| II | 1.874 | 1.984 | 1.804 | 2.414 | 3.059 | 2.533 | |||||
| III | 3.03 | 2.329 | 3.337 | 2.132 | 3.493 | 3.601 | |||||
| Emma | I | 3.508 | 3.613 | 2.831 | 4.288 | 2.636 | 1.564 | 2.902 | |||
| II | 2.682 | 2.388 | 1.859 | 2.554 | 2.301 | 2.447 | 2.349 | ||||
| III | 3.138 | 3.916 | 2.32 | 4.661 | 3.805 | 3.119 | 4.996 | ||||
| Skala | I | 1.225 | 1.652 | 2.237 | 2.624 | 2.592 | 3.04 | 2.13 | 3.6 | ||
| II | 1.558 | 1.761 | 2.595 | 2.369 | 3.889 | 4.049 | 1.965 | 3.416 | |||
| III | 1.689 | 3.076 | 2.776 | 4.319 | 4.505 | 4.365 | 4.508 | 3.371 | |||
| Skaut | I | 2.178 | 2.155 | 3.236 | 3.587 | 3.002 | 3.896 | 2.985 | 4.079 | 2.23 | |
| II | 2.37 | 2.491 | 3.157 | 3.746 | 3.91 | 3.868 | 2.467 | 4.01 | 2.167 | ||
| III | 2.414 | 2.929 | 3.511 | 4.337 | 3.867 | 4.067 | 3.897 | 4.51 | 2.588 | ||
| Karta | I | 1.988 | 2.236 | 2.749 | 3.546 | 2.863 | 3.109 | 2.298 | 3.138 | 1.681 | 1.664 |
| II | 1.433 | 1.634 | 2.377 | 3.069 | 3.333 | 3.403 | 1.888 | 3.193 | 1.825 | 1.335 | |
| III | 1.8 | 1.756 | 2.869 | 3.016 | 3.172 | 3.485 | 2.665 | 4.288 | 2.871 | 1.62 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radkowski, A.; Bocianowski, J.; Nowosad, K.; Piwowarczyk, E.; Bakinowska, E.; Radkowska, I.; Wolski, K. Comparison of the Yield and Chemical Composition of Eleven Timothy (Phleum pratense L.) Genotypes under Three Locations in Poland. Agronomy 2020, 10, 1743. https://doi.org/10.3390/agronomy10111743
Radkowski A, Bocianowski J, Nowosad K, Piwowarczyk E, Bakinowska E, Radkowska I, Wolski K. Comparison of the Yield and Chemical Composition of Eleven Timothy (Phleum pratense L.) Genotypes under Three Locations in Poland. Agronomy. 2020; 10(11):1743. https://doi.org/10.3390/agronomy10111743
Chicago/Turabian StyleRadkowski, Adam, Jan Bocianowski, Kamila Nowosad, Elżbieta Piwowarczyk, Ewa Bakinowska, Iwona Radkowska, and Karol Wolski. 2020. "Comparison of the Yield and Chemical Composition of Eleven Timothy (Phleum pratense L.) Genotypes under Three Locations in Poland" Agronomy 10, no. 11: 1743. https://doi.org/10.3390/agronomy10111743
APA StyleRadkowski, A., Bocianowski, J., Nowosad, K., Piwowarczyk, E., Bakinowska, E., Radkowska, I., & Wolski, K. (2020). Comparison of the Yield and Chemical Composition of Eleven Timothy (Phleum pratense L.) Genotypes under Three Locations in Poland. Agronomy, 10(11), 1743. https://doi.org/10.3390/agronomy10111743