Effect of Vanadium on the Uptake and Distribution of Organic and Inorganic Forms of Iodine in Sweetcorn Plants during Early-Stage Development


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Cultivation
2.2. Analysis of Fresh Plant Material Sample
2.3. Analysis of Dry Samples of Roots, Stems, and Leaves
2.4. Data Analysis
3. Results
3.1. Plant Growth
3.2. Uptake and Accumulation of Iodine in Maize Plants
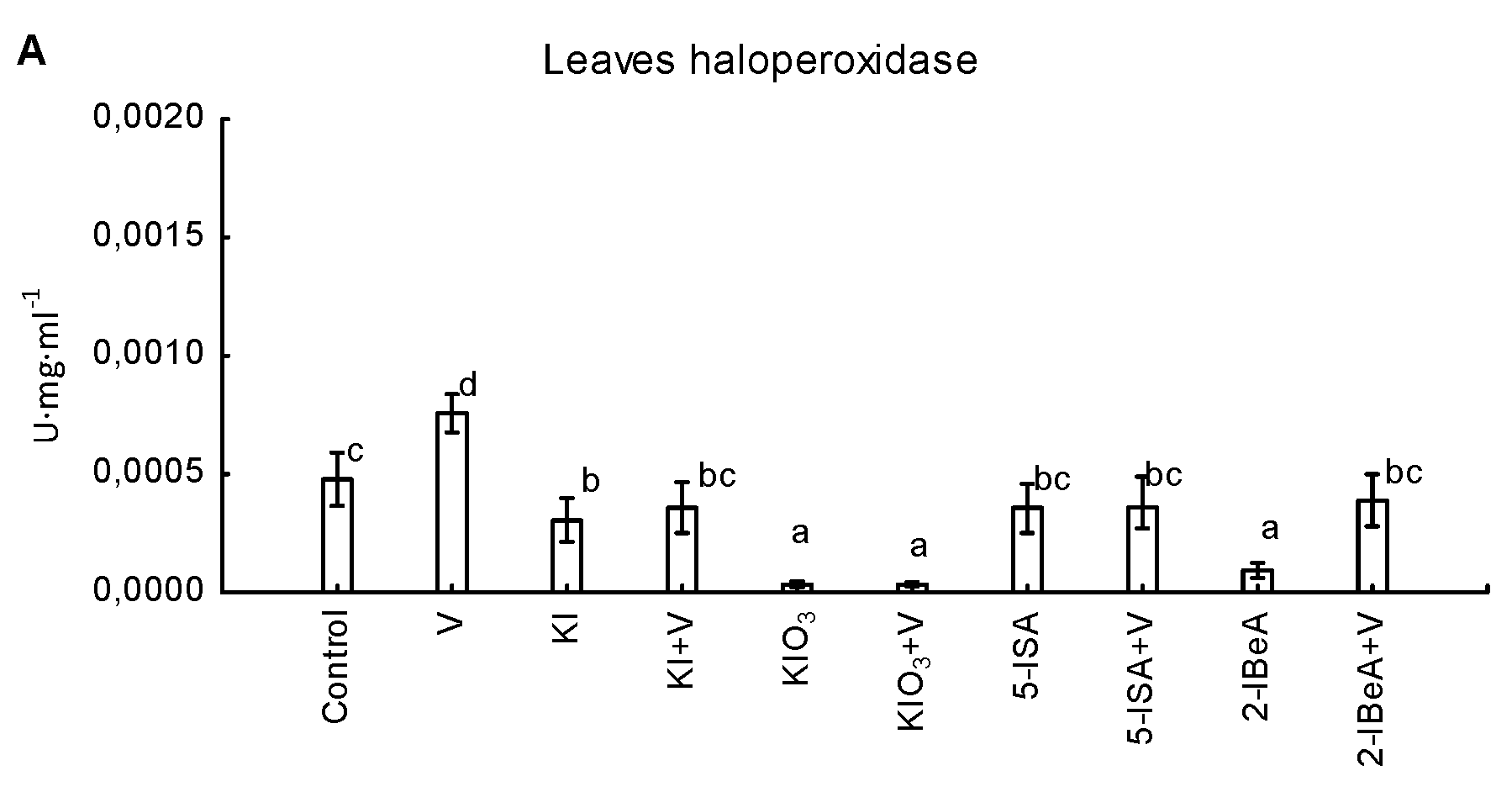
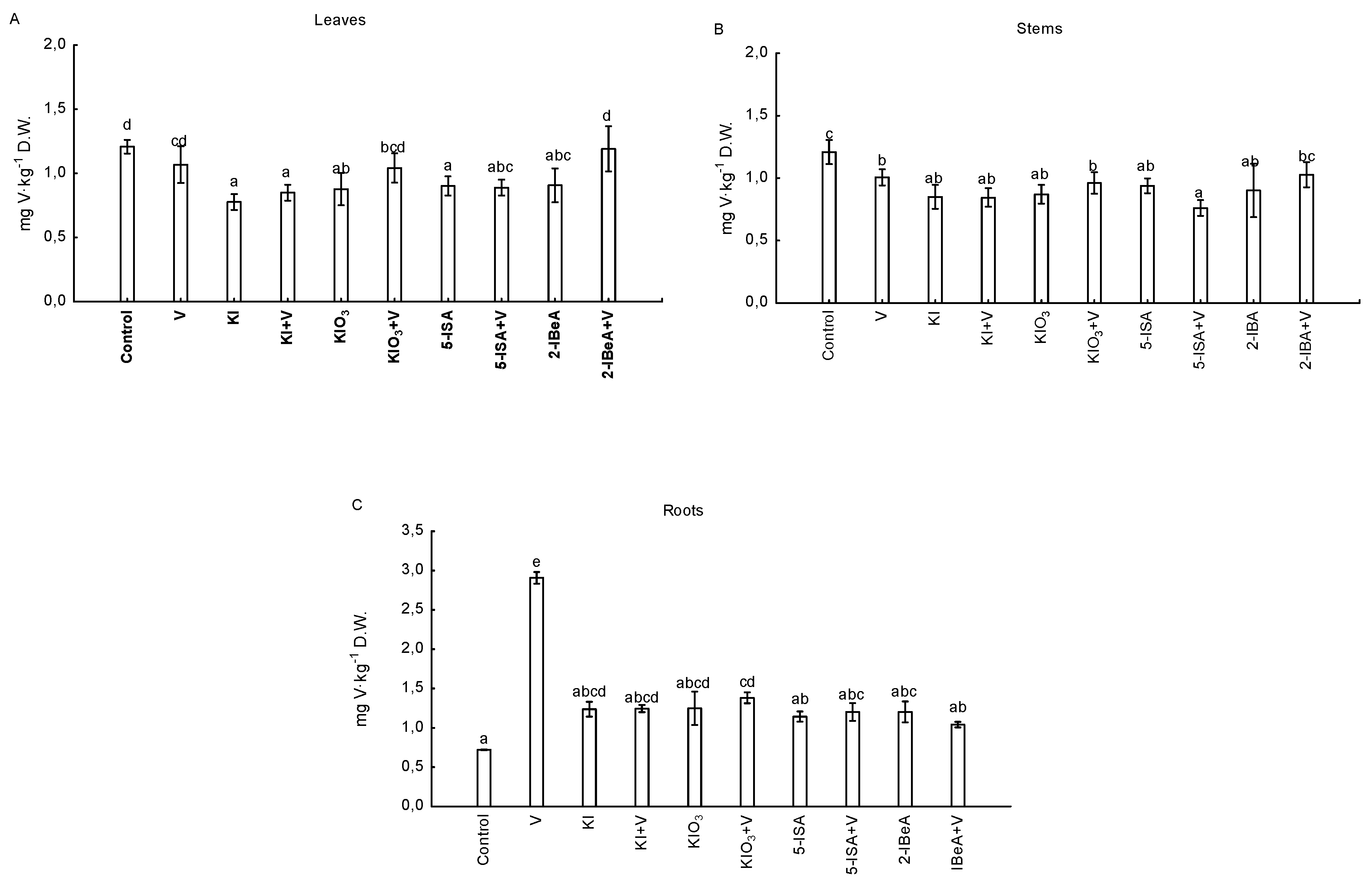
3.3. Activity of vHPO and Content and Uptake of Vanadium by Maize Plants
3.4. Contents of Macroelements and Microelements in Maize Plants
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lazarus, J.H. The importance of iodine in public health. Environ. Geochem. Health 2015, 37, 605–618. [Google Scholar] [CrossRef] [PubMed]
- Weng, H.T.; Hong, C.L.; Xia, T.H.; Baao, L.T.; Liu, H.P.; Li, D.W. Iodine biofortification of vegetable plants—An innovative method for iodine supplementation. Chin. Sci. Bull. 2013, 58, 2066–2072. [Google Scholar] [CrossRef]
- Zimmermann, M.B. Iodine Deficiency. Endocr. Rev. 2009, 30, 376–408. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, M.; Azizi, F.; Hedaya, M. Iodine deficiency status in the WHO Eastern MediterraneanRegion: A systematic review. Environ. Geochem. Health 2018, 40, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Blasco, B.; Rios, J.J.; Cervilla, L.M.; Sánchez-Rodrigez, E.; Ruiz, J.M.; Romero, L. Iodine biofortification and antioxidant capacity of lettuce: Potential benefits for cultivation and human health. Ann. Appl. Biol. 2008, 152, 289–299. [Google Scholar] [CrossRef]
- Smoleń, S.; Kowalska, I.; Sady, W. Assesment of biofortyfication with iodine and selenium of lettuce cultivated in the NFT hydroponic system. Sci. Hortic. 2014, 166, 9–16. [Google Scholar] [CrossRef]
- Zhu, Y.G.; Huang, Y.Z.; Hu, Y.; Liu, Y.X. Iodine uptake by spinach (Spinacia oleracea L.) plants grown in solution culture: Effect of iodine species and solution concentrations. Environ. Int. 2003, 29, 33–37. [Google Scholar] [CrossRef]
- Dai, J.L.; Zhu, Y.G.; Huang, Y.Z.; Zang, M.; Song, J.L. Availability of iodide and iodate to spinach (Spinacia oleracea L.) in relation to total iodine in soil solution. Plant Soil 2006, 289, 301–308. [Google Scholar] [CrossRef]
- Landini, M.; Gonzali, S.; Perata, P. Iodine biofortification in tomato. J. Plant Nutr. Soil Sci. 2011, 174, 480–486. [Google Scholar] [CrossRef]
- Weng, H.X.; Yan, A.L.; Hong, C.L.; Xie, L.L.; Qin, Y.C.; Cheng, C.Q. Uptake of different species of iodine by water spinach and its effect to growth. Biol. Trace Elem. Res. 2008, 124, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.L.; Zhu, Y.G.; Zhang, M.; Huang, Y.Z. Selecting iodine-enriched vegetables and the residual effect of iodate application to soil. Biol. Trace Elem. Res. 2004, 101, 265–276. [Google Scholar] [CrossRef]
- Smoleń, S.; Kowalska, I.; Skoczylas, Ł.; Liszka-Skoczylas, M.; Grzanka, M.; Halka, M.; Sady, W. The effect of salicylic acid on biofortification with iodine and selenium and the quality of potato cultivated in the NFT system. Sci. Hortic. 2018, 240, 530–543. [Google Scholar] [CrossRef]
- Maćkowiak, C.L.; Grossl, P.R. Iodate and iodide effects on iodine uptake and partitioning in rice (Oryza sativa L.) grown in solution culture. Plant Soil 1999, 212, 133–141. [Google Scholar]
- Cakmak, I.; Prom-u-Thai, C.; Guliherme, L.R.G.; Rashid, A.; Hora, K.H.; Yazici, A.; Savasli, E.; Kalayci, M.; Tutus, Y.; Phuphong, P.; et al. Iodine biofortification of wheat, rice and maize through fertilizer strategy. Plant Soil 2017, 418, 319–335. [Google Scholar] [CrossRef]
- Germ, M.; Stibilj, V.; Šircelj, H.; Jerše, A.; Kroflič, A.; Golob, A.; Maršić, N.K. Biofortification of common buckwheat microgreens and seeds with different forms of selenium and iodine. J. Sci. Food Agric. 2019, 99, 4353–4362. [Google Scholar] [CrossRef] [PubMed]
- Halka, M.; Smoleń, S.; Czernicka, M.; Klimek-Chodacka, M.; Pitala, J.; Tutaj, K. Iodine biofortification through expression of HMT, SAMT and S3H genes in Solanum lycopersicum L. Plant Physiol. Biochem. 2019, 144, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Sularz, O.; Smoleń, S.; Koronowicz, A.; Kowalska, I.; Leśzczyńska, T. Chemical composition of lettuce (Lactuca sativa L.) biofortified with iodine by KIO3, 5-iodo-, and 3.5-diiodosalicylic acid in a hydroponic cultivation. Agronomy 2020, 10, 1022. [Google Scholar] [CrossRef]
- Lawson, P.G.; Daum, D.; Czauderna, R.; Meuser, H.; Hartling, J.W. Soil versus foliar iodine fertilization as a biofortification strategy for field-grown vegetables. Front. Plant Sci. 2015, 6, 450. [Google Scholar] [CrossRef]
- Fuge, R. Geochemistry of iodine in relation to iodine deficiency diseases. Geol. Soc. 1996, 113, 201–211. [Google Scholar] [CrossRef]
- Leblanc, C.; Colin, C.; Cosse, A.; Delage, L.; la Barre, S.; Morin, P.; Fiévet, B.; Voiseux, C.; Ambroise, Y.; Verhaeghe, E.; et al. Iodine transfers in the coastal marine environment: The key role of brown algae and of their vanadium-dependent haloperoxidases. Biochemie 2006, 88, 1773–1785. [Google Scholar] [CrossRef]
- Küpper, F.C.; Schweigert, N.; Ar Gall, E.; Legendre, J.M.; Vilter, H.; Kloareg, B. Iodine uptake in Laminariales involves extracellular. haloperoxidase-mediated oxidation of iodide. Planta 1998, 207, 163–171. [Google Scholar]
- Tymon, T.M.; Miller, E.P.; Gonzalez, J.L.; Raab, A.; Kupper, F.C.; Carrano, C.J. Some aspects of the iodine metabolism of the giant kelp Macrocystis pyrifera (phaeophyceae). J. Inorg. Biochem. 2017, 177, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Butler, A.; Carter-Franklin, J.N. The role of vanadium bromoperoxidase in the biosynthesis of halogenated marine natural products. Nat. Prod. Rep. 2004, 21, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Medrano-Macías, J.; Leija-Martínez, P.; González-Morales, S.; Juárez-Maldonado, A.; Benavides-Mendoza, A. Use of iodine to biofortify and promote growth and stress tolerance in crops. Front. Plant Sci. 2016, 7, 1146. [Google Scholar] [CrossRef] [PubMed]
- Roberts, G.K.; Stout, M.D.; Sayers, B.; Fallacar, D.M.; Hejtmancik, M.R.; Waidyanatha, S.; Hooth, M. 14 day toxicity studies of tetravalent and pentavalent vanadium compounds in Harlan Sprague Dawley rats and B6C3F1/N mice via drinking water exposure. Toxicol. Rep. 2016, 3, 531–538. [Google Scholar] [CrossRef]
- Vachirapatama, N.; Jirakiattikul, Y.; Dicinoski, G.; Townsend, A.T.; Haddad, P.R. Effect of vanadium on plant growth and its accumulation in plant tissues. Songklanakarin J. Sci. Technol. 2011, 33, 255–261. [Google Scholar]
- Kaplan, D.I.; Adriano, D.C.; Carlson, C.L.; Sajwan, K.S. Vanadium: Toxicity and accumulation by beans. Water. Air. Soil Pollut. 1990, 49, 81–91. [Google Scholar] [CrossRef]
- Akoumianaki-Ioannidou, A.; Barouchas, P.E.; Ilia, E.; Kyramariou, A.; Moustakas, N.K. Effect of vanadium on dry matter and nutrient concentration in sweet basil (Ocimum basilicum L.). Aust. J. Crop Sci. 2016, 10, 199. [Google Scholar]
- Vara, F.; Serrano, R. Partial purification and properties of the proton-translocating ATPase of plant plasma membranes. J. Biol. Chem. 1982, 257, 12826–12830. [Google Scholar] [PubMed]
- Wdowikowska, A.; Kłobus, G. Plant P-type ATPases. Postępy Biochem. 2011, 57, 85–91. [Google Scholar]
- Karoń, B. Doświadczenie wazonowe nad wpływem wanadu na plon i skład chemiczny niektórych roślin. Rocz. Glebozn. 1977, 28, 155–172. [Google Scholar]
- Akoumianaki Ioannidou, A.; Barouchas, P.E.; Kyramariou, A.; Ilia, E.; Moustakas, N.K. Effect of vanadium on dry matter and nutrient concentration in pennyroyal (Mentha pulegium L). Bull. UASVM Hortic. 2015, 72, 295–298. [Google Scholar] [CrossRef][Green Version]
- Evangelou, A.M. Vanadium in cancer treatment. Crit. Rev. Oncol. Hematol. 2002, 42, 249–265. [Google Scholar] [CrossRef]
- Srivastava, A.K.; Mehdi, M.Z. Insulino-mimetic and anti-diabetic effects of vanadium compounds. Diabet. Med. 2005, 22, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Panchal, S.K.; Wanyonyi, S.; Brown, L. Selenium, Vanadium, and Chromium as Micronutrients to Improve Metabolic Syndrome. Curr. Hypertens. Rep. 2017, 19, 10. [Google Scholar] [CrossRef] [PubMed]
- Urban, J.; Antonowicz-Juchniewicz, J.; Andrzejak, R. WANAD—ZAGROŻENIA I NADZIEJE. Med. Pr. 2001, 52, 125–133. [Google Scholar] [PubMed]
- Barceloux, D.G.; Barceloux, D. Vanadium. J. Toxicol. Clin. Toxicol. 1999, 37, 265–278. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, B.; Patra, B.; Mahapatra, S.; Banerjee, P.; Tiwari, A.; Chatterjee, M. Vanadium—An element of atypical biological significance. Toxicol. Lett. 2009, 150, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Gruzewska, K.; Michno, A.; Pawelczyk, T.; Bielarczyk, H. Essentiality and toxicity of vanadium supplements in health and pathology. J. Physiol. Pharmacol. 2014, 65, 603–611. [Google Scholar]
- Boden, G.; Chen, X.; Ruiz, J.; van Rossum, G.D.V.; Turco, S. Effects of vanadyl sulfate on carbohydrate and lipid metabolism in patients with non-insulin-dependent diabetes mellitus. Metabolism 1996, 45, 1130–1135. [Google Scholar] [CrossRef]
- Nielsen, F.H.; Uthus, E.O. The Essentiality and Metabolism of Vanadium. In Physiology and Biochemistry; Kluwer Academic Publishers: Amsterdam, The Netherlands, 1990; pp. 51–62. [Google Scholar]
- WHO. Vanadium Pentoxide and Other Inorganic Vanadium Compounds; The World Health Organization: Geneva, Switzerland, 2001. [Google Scholar]
- Thumbo, P.; Yates, A.A.; Schlicker, S.; Poos, M. Dietary Reference Intakes: Vitamin, A, viramin K, Arsenic, Boron, Chromium, Copper, Iodine, Iron. Manganese, Molybdenum, Nickle, Silicon, Vanadium, and Zinc. J. Am. Diet. Assoc. 2001, 3, 294–301. [Google Scholar]
- Leyva, R.; Sánchez-Rodríguez, E.; Ríos, J.J.; Rubio-Wilhelmi, M.M.; Romero, L.; Ruiz, J.M.; Blasco, B. Beneficial effects of exogenous iodine in lettuce plants subjected to salinity stress. Plant Sci. 2011, 181, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Bajpai, M.; Majumdar, R.; Mishra, P. Response of iodine on antioxidant levels of Glycine max L. Grown under Cd2+ stress. Adv. Biol. Res. 2015, 9, 40–48. [Google Scholar]
- Matsugo, S.; Kanamori, K.; Sugiyama, H.; Misu, H.; Takamura, T. Physiological roles of peroxido-vanadium complexes: Leitmotif as their signal transduction pathway. J. Inorg. Biochem. 2015, 147, 9398. [Google Scholar] [CrossRef] [PubMed]
- Caffagni, A.; Pecchioni, N.; Meriggi, P.; Bucci, V.; Sabatini, E.; Acciarri, N.; Ciriaci, T.; Pulcini, L.; Felicioni, N.; Beretta, M.; et al. Iodine uptake and distribution in horticultural and fruit tree species. Ital. J. Agron. 2012, 7, e32. [Google Scholar] [CrossRef]
- Gonzali, S.; Kiferle, C.; Perata, P. Iodine biofortification of crops: Agronomic biofortification, metabolic engineering and iodine bioavailability. Curr. Opin. Biotechnol. 2017, 44, 16–26. [Google Scholar] [CrossRef]
- Hong, C.L.; Weng, H.X.; Yan, A.L.; Islam, E.U. The fate of exogenous iodine in pot soil cultivated with vegetables. Environ. Geochem. Health 2009, 31, 99–108. [Google Scholar] [CrossRef]
- McLauchlana, C.C.; Murakamib, H.A.; Wallacea, C.A.; Crans, D.C. Coordination environment changes of the vanadium in vanadium-dependenthaloperoxidase enzymes. J. Inorg. Biochem. 2018, 186, 267–279. [Google Scholar] [CrossRef]
- Timmins, A.; Visser, S.P. Enzymatic halogenases and haloperoxidases: Computational studies on mechanism and function. Adv. Protein Chem. Struct. Biol. 2015, 100, 113–151. [Google Scholar]
- Smoleń, S.; Kowalska, I.; Halka, M.; Ledwożyw-Smoleń, I.; Grzanka, M.; Skoczylas, Ł.; Czernicka, M.; Pitala, J. Selected aspects of iodate and iodosalicylate metabolism in lettuce including the activity of vanadium dependent haloperoxidases as affected by exogenous vanadium. Agronomy 2019, 10, 1. [Google Scholar] [CrossRef]
- Smoleń, S.; Ledwożyw-Smoleń, I.; Sady, W. The role of exogenous humic and fulvic acids in iodine biofortification in spinach (Spinacia oleracea L.). Plant Soil 2016, 402, 129–143. [Google Scholar]
- Pasławski, P.; Migaszewski, Z.M. The quality of element determinations in plant materials by instrumental methods. Pol. J. Environ. Stud. 2006, 15, 154–164. [Google Scholar]
- Taylor, M.J.C.; Saden, J.F. Spectrophotometric determination of vanadium(IV) and vanadium(V) in each other’s presence. Analyst 1994, 119, 1263–1276. [Google Scholar] [CrossRef]
- Yang, J.; Wang, M.; Jia, Y.; Gou, M.; Zeyer, J. Toxicity of vanadium in soil on soybean at different growth stages. Environ. Pollut. 2017, 231, 48e58. [Google Scholar] [CrossRef] [PubMed]
- Halka, M.; Klimek-Chodacka, M.; Smoleń, S.; Baranski, R.; Ledwożyw-Smoleń, I.; Sady, W. Organic iodine supply affects tomato plants differently than inorganic iodine. Physiol. Plant. 2018, 164, 290–306. [Google Scholar] [CrossRef] [PubMed]
- Sarma, H. Metal hyperaccumulation in plants: A revive focusing on phytoremediation technology. J. Environ. Sci. Technol. 2011, 4, 118–138. [Google Scholar] [CrossRef]
- Voogt, W.; Holwerda, H.T.; Khodabaks, R. Biofortification of lettuce (Lactuca sativa L.) with iodine: The effect of iodine form and concentration in the nutrient solution on growth. development and iodine uptake of lettuce grown in water culture. J. Sci. Food Agric. 2010, 90, 906–913. [Google Scholar] [CrossRef]
- Welch, R.M.; Huffman, E.W. Vanadium and plant nutrition: The growth of lettuce (Lactuca sativa L.) and Tomato (Lycopersicon esculentum Mill.) plants in nutrient solutions low in vanadium. Plant Physiol. 1973, 52, 183–185. [Google Scholar] [CrossRef]
- Imtiaz, M.; Ashraf, M.; Rizwan, M.S.; Nawaz, M.A.; Rizwan, M.; Mehmood, S.; Ali, M. Vanadium toxicity in chickpea (Cicer arietinum L.) grown in red soil: Effects on cell death. ROS and antioxidative systems. Ecotoxicol. Environ. Saf. 2018, 158, 139–144. [Google Scholar] [CrossRef]
- Saco, D.; Martín, S.; San José, P. Vanadium distribution in roots and leaves of Phaseolus vulgaris: Morphological and ultrastructural effects. Biol. Plant. 2013, 57, 128–132. [Google Scholar] [CrossRef]
- Chongkid, B.; Vachirapatama, N.; Jirakiattikul, Y. Effects of V on rice growth and vandium accumulation in rice tissues. Kasetsart J. 2007, 41, 28–33. [Google Scholar]
- Krzepiłko, A.; Zych-Wężyk, I.; Święciło, A.; Molas, J.; Skwaryło-Bednarz, B. Effect of iodine biofortification of lettuce seedlings on their mineral composition and biological quality. J. Elem. 2016, 21, 1071–1080. [Google Scholar] [CrossRef]
- Smoleń, S.; Sady, W. Influence of iodine form and application method on the effectiveness of iodine biofortification, nitrogen metabolism as well as the content of mineral nutrients and heavy metals in spinach plants (Spinacia oleracea L.). Sci. Hortic. 2012, 143, 176–183. [Google Scholar]
- Imtiaz, M.; Rizwan, M.S.; Xiong, S.; Li, H.; Ashraf, M.; Shahzad, S.M.; Shahzad, M.; Rizwan, M.; Tu, S. Vanadium, recent advancements and research prospects: A review. Environ. Int. 2015, 80, 79–88. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dose of Applied Compounds (μM) | ||
|---|---|---|
| Treatments | Iodine | Vanadium |
| Control | - | - |
| V | - | 0.1 μM |
| KI | 10 μM | - |
| KI+V | 10 μM | 0.1 μM |
| KIO3 | 10 μM | - |
| KIO3+V | 10 μM | 0.1 μM |
| 5-ISA | 10 μM | - |
| 5-ISA+V | 10 μM | 0.1 μM |
| 2-IBeA | 10 μM | - |
| 2-IBeA+V | 10 μM | 0.1 μM |
| Treatment | Root Length (cm) | Plant Height (Aerial Part of the Plant) (cm) | Root Weight (g) | Plant Weight (Aerial Part of the Plant) (g) | Shoot/Root Ratio |
|---|---|---|---|---|---|
| Control | 12.08 ± 1.39 a | 58.17 ± 0.64 b,c | 5.11 ± 0.71 a | 26.82 ± 2.21 a | 5.1 ± 0.61 a,b,c |
| V | 16.83 ± 1.75 b | 58.17 ± 1.37 b,c | 7.17 ± 1.15 a,b | 34.43 ± 4.42 a,b | 5.2 ± 0.88 a,b,c |
| KI | 22.50 ± 2.10 c,d | 61.50 ± 1.95 d | 14.37 ± 3.21 c,d | 54.06 ± 5.98 c,d | 5.1 ± 1.20 a,b,c |
| KI+V | 16.46 ± 2.53 b | 65.50 ± 1.00 e | 14.52 ± 3.43 c,d | 62.27 ± 5.56 d | 6.0 ± 1.38 b,c |
| KIO3 | 17.58 ± 2.94 b | 62.33 ± 1.36 d | 10.07 ± 2.33 a,b,c | 50.65 ± 6.51 c,d | 7.0 ± 1.42 c |
| KIO3+V | 20.83 ± 1.71 c,d | 57.92 ± 2.29 b | 8.52 ± 1.03 a,b,c | 43.48 ± 5.69 b,c | 5.0 ± 0.50 a,b,c |
| 5-ISA | 20.33 ± 1.92 c | 60.96 ± 2.19 c,d | 12.61 ± 1.50 b,c,d | 40.14 ± 2.71 b,c | 3.2 ± 0.32 a,b |
| 5-ISA+V | 26.83 ± 0.37 e | 59.50 ± 1.98 b,c,d | 18.95 ± 2.13 d,e | 49.95 ± 6.04 c,d | 2.7 ± 0.23 a |
| 2-IBeA | 23.00 ± 1.49 d | 57.38 ± 3.30 b | 23.02 ± 5.36 e | 44.45 ± 8.08 b,c | 2.5 ± 0.36 a |
| 2-IBeA+V | 21.33 ± 2.18 c,d | 49.00 ± 3.43 a | 9.24 ± 1.48 a,b,c | 30.69 ± 3.48 a,b | 4.0 ± 0.94 a,b,c |
| Leaves (g kg−1 d.w) | |||||
|---|---|---|---|---|---|
| Treatment | Ca | K | Mg | P | S |
| Control | 1.13 ± 0.025 d | 4.68 ± 0.133 f | 0.31 ± 0.003 d | 0.68 ± 0.046 e | 0.24 ± 0.005 a |
| V | 1.06 ± 0.026 c,d | 4.41 ± 0.143 e | 0.27 ± 0.002 c | 0.58 ± 0.031 d | 0.25 ± 0.002 a,b,c |
| KI | 0.79 ± 0.010 a | 4.02 ± 0.140 c,d | 0.21 ± 0.004 a | 0.47 ± 0.015 b | 0.25 ± 0.009 a,b |
| KI+V | 0.92 ± 0.006 b | 3.79 ± 0.087 b,c | 0.22 ± 0.004 a,b | 0.43 ± 0.018 a,b | 0.26 ± 0.006 b,c |
| KIO3 | 0.90 ± 0.016 b | 4.27 ±0.160 d,e | 0.24 ± 0.008 b | 0.51 ± 0.028 c | 0.25 ± 0.005 a,b,c |
| KIO3+V | 1.12 ± 0.035 d | 4.22 ± 0.128 d,e | 0.30 ± 0.004 d | 0.59 ± 0.028 d | 0.26 ± 0.005 b,c |
| 5-ISA | 0.93 ± 0.051 b | 4.13 ± 0.041 d | 0.23 ± 0.006 a,b | 0.52 ± 0.002 c | 0.26 ± 0.015 c |
| 5-ISA+V | 0.92 ± 0.041 b | 3.61 ± 0.028 a,b | 0.24 ± 0.004 b | 0.43 ± 0.002 a | 0.26 ± 0.013 b,c |
| 2-IBeA | 0.98 ± 0.073 b,c | 3.49 ± 0.141 a | 0.22 ± 0.011 a,b | 0.45 ± 0.014 a,b | 0.26 ± 0.023 b,c |
| 2-IBeA+V | 1.23 ± 0.076 e | 3.85 ± 0.190 b,c | 0.26 ± 0.014 c | 0.56 ± 0.027 d | 0.26 ± 0.013 b,c |
| Stems (g kg−1 d.w) | |||||
| Control | 0.95 ± 0.031 f | 8.15 ± 0.282 d | 0.41 ± 0.013 e | 0.68 ± 0.008 e | 0.19 ± 0.011 a,b |
| V | 0.84 ± 0.021 e | 8.33 ± 0.491 d | 0.37 ± 0.006 d | 0.64 ± 0.017 d,e | 0.21 ± 0.013 c |
| KI | 0.70 ± 0.029 a | 6.61 ± 0.643 a,b,c | 0.29 ± 0.026 a,b | 0.49 ± 0.011 a,b | 0.19 ± 0.01 a |
| KI+V | 0.71 ± 0.013 a,b | 5.63 ± 0.802 a | 0.26 ± 0.022 a | 0.42 ± 0.044 a | 0.19 ± 0.02 a |
| KIO3 | 0.75 ± 0.028 c,d | 7.37 ± 0.703 b,c,d | 0.33 ± 0.031 c,d | 0.54 ± 0.01 b,c | 0.20 ± 0.01 a,b,c |
| KIO3+V | 0.83 ± 0.041 e | 7.87 ± 0.66 d | 0.37 ± 0.024 d | 0.62 ± 0.007 d,e | 0.19 ± 0.003 a |
| 5-ISA | 0.78 ± 0.017 d | 7.44 ± 0.46 c,d | 0.31 ± 0.013 b,c | 0.58 ± 0.013 c,d | 0.20 ± 0.014 a,b,c |
| 5-ISA+V | 0.73 ± 0.024 a,b,c | 6.25 ± 0.492 a,b | 0.31 ± 0.014 b,c | 0.49 ± 0.01 a,b | 0.19 ± 0.012 a,b,c |
| 2-IBeA | 0.74 ± 0.049 b,c | 5.94 ± 0.596 a | 0.29 ± 0.021 a,b | 0.51 ± 0.004 b,c | 0.19 ± 0.021 a |
| 2-IBeA+V | 0.86 ± 0.010 e | 7.33 ± 0.443 b,c,d | 0.35 ± 0.022 d | 0.67 ± 0.017 e | 0.21 ± 0.01 b,c |
| Roots (mg kg−1 d.w) | |||||
| Control | 0.94 ± 0.004 a,b,c | 2.95 ± 0.019 d | 0.20 ± 0.001 b,c | 0.23 ± 0.001 f | 0.48 ± 0.003 c,d |
| V | 0.87 ± 0.006 a | 2.89 ± 0.031 d | 0.17 ± 0.002 a | 0.18 ± 0.002 d | 0.46 ± 0.003 b,c,d |
| KI | 1.04 ± 0.002 d | 2.57 ± 0.003 b | 0.22 ± 0.001 c | 0.18 ± 0.001 d | 0.45 ± 0.002 b,c |
| KI+V | 0.99 ± 0.005 b,c,d | 2.37 ± 0.007 a,b | 0.22 ± 0.001 c | 0.15 ± 0.001 b | 0.38 ± 0.002 a |
| KIO3 | 1.00 ± 0.033 b,c,d | 2.59 ± 0.084 b,c | 0.20 ± 0.008 c | 0.16 ± 0.005 c | 0.46 ± 0.015 b,c,d |
| KIO3+V | 0.97 ± 0.002 b,c,d | 2.82 ± 0.001 c,d | 0.18 ± 0.001 a,b | 0.22 ± 0.001 e | 0.45 ± 0.002 b,c,d |
| 5-ISA | 0.84 ± 0.006 a | 2.91 ± 0.013 d | 0.18 ± 0.001 a,b | 0.20 ± 0.002 d | 0.41 ± 0.003 a,b |
| 5-ISA+V | 0.90 ± 0.004 a,b | 2.14 ± 0.010 a | 0.20 ± 0.001 b,c | 0.14 ± 0.000 a | 0.38 ± 0.002 a |
| 2-IBeA | 1.03 ± 0.051 c,d | 2.41 ± 0.112 b | 0.25 ± 0.013 d | 0.15 ± 0.007 b | 0.50 ± 0.024 d,e |
| 2-IBeA+V | 1.03 ± 0.004 c,d | 3.61 ± 0.026 e | 0.20 ± 0.001 b,c | 0.25 ± 0.001 g | 0.55 ± 0.002 f |
| Leaves (mg kg−1 d.w) | ||||||
|---|---|---|---|---|---|---|
| Treatment | B | Cu | Fe | Mn | Zn | Mo |
| Control | 0.85 ± 0.14 a | 15.77 ± 0.92 f | 134.91 ± 4.91 d | 104.87 ± 0.46 e | 48.60 ± 4.54 d | 0.04 ± 0.03 a,b |
| V | 1.56 ± 0.41 b | 14.70 ± 0.75 e | 118.64 ± 1.21 c | 92.65 ± 0.37 d | 40.51 ± 2.00 c | 0.86 ± 0.12 a,b |
| KI | 2.29 ± 0.54 c | 11.53 ± 0.87 a,b | 114.09 ± 3.03 c | 65.83 ± 5.67 a | 35.22 ± 0.94 a,b,c | 1.31 ± 0.40 b |
| KI+V | 1.80 ± 0.28 b,c | 11.49 ± 0.85 a | 100.55 ± 2.45 a | 74.22 ± 3.73 b,c | 30.53 ± 1.15 a | 1.16 ± 0.27 b |
| KIO3 | 1.99 ± 0.46 b,c | 12.50 ± 0.79 c | 112.65 ± 2.76 b,c | 80.73 ± 6.08 c | 35.15 ± 1.22 a,b | 1.08 ± 0.30 a,b |
| KIO3+V | 1.81 ± 0.42 b,c | 14.20 ± 0.93 d,e | 117.84 ± 3.34 c | 95.77 ± 0.50 d | 38.96 ± 1.51 b,c | 0.56 ± 0.18 a,b |
| 5-ISA | 1.99 ± 0.49 b,c | 12.43 ± 0.80 b,c | 100.93 ± 2.10 a | 77.35 ± 2.37 b,c | 35.07 ± 1.28 a,b | 0.79 ± 0.25 a,b |
| 5-ISA+V | 1.42 ± 0.17 a,b | 11.61 ± 0.91 a,b,c | 103.91 ± 1.82 a,b | 71.44 ± 1.76 a,b | 32.87 ± 0.43 a | 0.59 ± 0.40 a,b |
| 2-IBeA | 1.51 ± 0.28 a,b | 11.37 ± 1.49 a | 98.69 ± 3.95 a | 71.25 ± 2.08 a,b | 33.15 ± 2.09 a | 0.87 ± 0.32 a,b |
| 2-IBeA+V | 1.92 ± 0.39 b,c | 13.49 ± 1.18 d | 110.52 ± 5.28 b,c | 105.93 ± 4.69 e | 46.69 ± 2.80 d | <0.04 a |
| Stems (mg kg−1 d.w) | ||||||
| Control | 1.67 ± 0.46 g | 12.88 ± 0.91 g | 101.77 ± 4.09 d | 54.54 ± 6.93 e | 91.1 ± 77.31 g | <0.04 |
| V | 1.46 ± 0.35 d,e | 10.73 ± 0.59 f | 93.19 ± 1.19 c,d | 41.78 ± 2.59 d | 77.89 ± 1.17 f | <0.04 |
| KI | 1.33 ± 0.37 a–d | 8.29 ± 0.95 b,c | 76.07 ± 6.52 a | 32.78 ± 2.40 b,c | 51.55 ± 3.06 a,b | <0.04 |
| KI+V | 1.37 ± 0.31 c,d,e | 8.24 ± 0.66 b,c | 78.36 ± 5.21 a | 33.52 ± 0.67 b,c | 46.92 ± 3.24 a | <0.04 |
| KIO3 | 1.34 ± 0.33 b–e | 9.16 ± 0.78 d | 94.52 ± 0.59 c,d | 34.47 ± 1.41 c | 66.45 ± 5.72 e | <0.04 |
| KIO3+V | 1.52 ± 0.35 e,f | 10.08 ± 0.77 e | 73.41 ± 5.44 a | 34.94 ± 1.94 c | 64.33 ± 3.86 d,e | <0.04 |
| 5-ISA | 1.30 ± 0.31 a–d | 8.72 ± 0.56 c,d | 80.17 ± 2.52 a,b | 31.23 ± 0.25 b | 59.43 ± 3.48 c,d | <0.04 |
| 5-ISA+V | 1.18 ± 0.29 a,b,c | 7.71 ± 0.51 a | 71.09 ± 6.99 a | 28.65 ± 0.40 a | 49.65 ± 2.05 a | <0.04 |
| 2-IBeA | 1.15 ± 0.28 a,b | 7.89 ± 0.98 a,b | 89.24 ± 2.13 b,c | 31.46 ± 1.90 b | 55.77 ± 5.56 b,c | <0.04 |
| 2-IBeA+V | 1.13 ± 0.28 a | 9.97 ± 0.77 e | 94.20 ± 4.54 c,d | 41.45 ± 0.61 d | 94.22 ± 1.44 g | <0.04 |
| Roots (mg kg−1 d.w) | ||||||
| Control | 1.74 ± 0.03 b | 36.74 ± 0.40 f | 146.66 ± 10.51 e,f | 52.62 ± 0.33 d | 63.91 ± 0.32 d | 1.11 ± 0.04 a,b,c |
| V | 1.49 ± 0.03 a | 20.42 ± 0.19 e | 116.55 ± 1.20 b,c | 32.39 ± 0.21 b | 96.25 ± 0.93 e | 1.36 ± 0.05 c |
| KI | 1.66 ± 0.02 a,b | 17.32 ± 0.15 c | 138.62 ± 0.43 d,e,f | 50.45 ± 2.12 d | 61.58 ± 0.33 c,d | 1.18 ± 0.15 b,c |
| KI+V | 1.59 ± 0.04 a,b | 14.50 ± 0.04 a | 153.51 ± 0.78 f | 35.24 ± 0.34 b | 59.24 ± 0.40 c | 0.76 ± 0.06 a |
| KIO3 | 1.50 ± 0.04 a | 19.41 ± 0.69 d,e | 129.04 ± 4.64 c,d,e | 41.60 ± 1.42 c | 44.18 ± 1.34 b | 0.95 ± 0.05 a,b,c |
| KIO3+V | 1.66 ± 0.07 a,b | 18.33 ± 0.07 c,d | 120.39 ± 0.95 b,c,d | 43.28 ± 0.36 c | 46.95 ± 0.27 b | 0.90 ± 0.04 a,b |
| 5-ISA | 1.63 ± 0.02 a,b | 14.15 ± 0.11 a | 100.09 ± 0.46 a,b | 31.01 ± 0.20 b | 44.78 ± 0.49 b | 0.88 ± 0.06 a,b |
| 5-ISA+V | 1.45 ± 0.09 a | 15.55 ± 0.06 a,b | 108.35 ± 1.00 a,b,c | 23.06 ± 0.22 a | 37.76 ± 0.31 a | 1.03 ± 0.10 a,b,c |
| 2-IBeA | 1.48 ± 0.04 a | 14.61 ± 0.67 a | 122.83 ± 6.42 c,d | 32.19 ± 1.57 b | 35.42 ± 1.37 a | 1.00 ± 0.14 a,b,c |
| 2-IBeA+V | 1.46 ± 0.03 a | 16.71 ± 0.12 b,c | 91.29 ± 3.11 a | 89.66 ± 0.50 e | 43.45 ± 0.19 b | 0.70 ± 0.06 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grzanka, M.; Smoleń, S.; Kováčik, P. Effect of Vanadium on the Uptake and Distribution of Organic and Inorganic Forms of Iodine in Sweetcorn Plants during Early-Stage Development. Agronomy 2020, 10, 1666. https://doi.org/10.3390/agronomy10111666
Grzanka M, Smoleń S, Kováčik P. Effect of Vanadium on the Uptake and Distribution of Organic and Inorganic Forms of Iodine in Sweetcorn Plants during Early-Stage Development. Agronomy. 2020; 10(11):1666. https://doi.org/10.3390/agronomy10111666
Chicago/Turabian StyleGrzanka, Marlena, Sylwester Smoleń, and Peter Kováčik. 2020. "Effect of Vanadium on the Uptake and Distribution of Organic and Inorganic Forms of Iodine in Sweetcorn Plants during Early-Stage Development" Agronomy 10, no. 11: 1666. https://doi.org/10.3390/agronomy10111666
APA StyleGrzanka, M., Smoleń, S., & Kováčik, P. (2020). Effect of Vanadium on the Uptake and Distribution of Organic and Inorganic Forms of Iodine in Sweetcorn Plants during Early-Stage Development. Agronomy, 10(11), 1666. https://doi.org/10.3390/agronomy10111666