1. Introduction
The introduction of hybrid cultivars with high early vigor, better weed control and increased use of nitrogen (N) fertilizers has improved canola (
Brassica napus L.) production in Australia [
1]. Annual production reached 3.9 Mt in 2017–2018 from approximately 1.8 Mt in 2008–2009 [
2]. However, there is still a large gap between actual and attainable yields [
1,
3,
4,
5]. Initially, in farming systems of southern Australian canola was grown after legume-based pastures to use high-mineral nitrogen (N) in the soil and to break the cereal root disease cycle. However, with the intensification of the cropping system and increased popularity of canola, canola is grown most often in continuous cropping systems where it is often grown after cereals in soils with low N status and where high rates of N are required to achieve high yields. Expansion of canola into new areas and changes to crop rotations and tillage practices has meant that the nutrition of canola has been reexamined in recent times [
6], with a strong focus on N management. In the Mediterranean environment of South Australia, water and N availability are the most critical factors for sustaining crop productivity [
5,
7,
8,
9], but often water-use efficiency (WUE) and nitrogen-use efficiency (NUE) are low.
Nitrogen is the nutrient most limiting to canola yield [
6]. Campbell et al. [
10] stated that N yield (N uptake) is a function of plant-available water as water is a major driver of yield in rainfed systems. Moreover, water deficits at critical growth stages can limit N uptake and utilization in plants [
11] and can reduce crop responses to N fertilizers [
12]. The critical phase of growth for yield in canola is just after the start of flowering [
13] when seed number is largely determined. Management practices that reduce the risk of N and water stress during this period will be important for achieving high yields.
Nitrogen is an expensive and difficult-to-manage input in environments where the availability of moisture is a major limitation to yield. Recovery of N in crops is generally less than 50% [
14], which cannot be justified from environmental and economic perspectives [
15]. Canola has a high nitrogen requirement and optimal nitrogen (N) management in environments with variable rainfall is a challenge. Norton for example, estimated that canola requires 80 kg N ha
−1 for each ton of grain yield [
6]. Therefore, for yields of 2–3 t ha
−1, which are commonly achieved in the medium rainfall zones of southern and western Australia [
16], 160–240 kg N ha
−1 needs to be supplied from soil N reserves and fertilizer. These values are similar to the total N required for canola (220–245 kg N ha
−1) estimated from simulation modeling for the medium and high rainfall zones [
17]. Applying all the N at the start of the season has inherent risks because the difficulty in predicting yield and hence the N requirement of crops and also because of canola establishment and seedling growth are sensitive to high rates of N fertilizer at sowing [
18,
19,
20]. Even in growing seasons with above-average rainfall, applying all the N fertilizer at the seedling stage could result in poor N efficiency by mismatching N supply with crop N demand.
Delaying or splitting applications of N is a reasonable strategy for managing risk. However, responses to timing of N have been variable: some studies have shown yield improvements with split N applications compared with a single application of N [
21,
22], whereas other studies found no improvement in yield with split N application than a single application [
21,
23,
24]. On average there may be little difference in yield between N applied at sowing or with a split application although the analysis of Norton [
6] and other recent work suggest that the post-application moisture availability has a large influence on the variation in responses [
25]. However, the interaction between N management and moisture availability has been little studied. It is often suggested that growers should manage fertilizer N in response to water availability and crop demand in these environments [
7,
9,
26], but often N rate and timing are selected based on logistics of N application rather than based on an understanding of crop demand of N at different phenological growth stages.
Applying N at different growth stages may have different influences on yield by its differential effect on seed number m
−2 (sink size) and seed weight and the degree of compensation between the two. Canola is an indeterminate crop and seed set and seed growth overlap, which complicates the source–sink relationship compared to determinate crops like wheat. Seed yield in canola can be affected by both sink and source limitations depending on the growth stage [
27] which can be influenced by the timing of N. Some studies based on the plant N status at different growth stages showed the importance of N at the rosette to green-bud growth stages of canola [
25,
28,
29] which would largely influence seed number. In contrast, Dreccer et al. [
30] argued that yield of oilseed rape was source-limited during the pod filling stages, and this limitation could be tackled by a targeted application of N at the pod filling stage. Based upon a simulation study, Habekotté [
31] suggested improving the source and sink capacity simultaneously to raise the potential yield of winter canola. In a recent study, it was reported that increasing the sink capacity with improved pre-flowering biomass production can improve the seed yield of canola and mustard [
25].
It is widely accepted that the management of fertilizer inputs is one of the most important tools for the improvement of yield, NUE and WUE in rainfed environments [
32,
33,
34], but there have been few analyses of the interaction between moisture and N management in canola. In the trials reported here, the effect of moisture was examined in two ways: irrigation was used to manipulate the water supply at one site and the response to N management was compared at two sites with different rainfall. Consequently, we hypothesize that post-sowing split applications of N at key growth stages can improve the yield, NUE and WUE of canola over its single application under different water regimes.
4. Discussion
Significant responses to N of between 16% and 69% were measured across both sites. The greatest increase occurred with 100 kg N ha
−1 and there was little further yield increase at 200 kg N ha
−1. This is consistent with the estimates of fertilizer N requirement based on an N balance described by Norton [
47] which predicted fertilizer rates of between 70 kg N ha
−1 and 100 kg N ha
−1 for canola yielding 3 t ha
−1, which was achieved under irrigation at Roseworthy and at Tarlee. However, a significant response to 100 kg N ha
−1 was also measured in the rainfed trial at Roseworthy, which the N balance predicted to be nonresponsive to N. This suggest that in-season mineralization of N was overestimated in the N balance and/or the recovery of mineral N by the crop was poor in the rainfed treatment. Half the estimated soil mineral N at sowing was found in the top 20 cm at Roseworthy and there was approximately 30 kg N/ha below 40 cm, which remained dry throughout the season. Therefore, this subsoil mineral may not have been able to be used effectively.
The responses in yield to N were affected mainly by the rate of N rather than the timing of N application. There was no significant effect of the time of N on the yield response to N within the range of yields in the current experiments (approximately 2–3 t ha
−1) irrespective of the moisture availability because similar effects were found under rainfed conditions at medium and high rainfall sites and in the rainfed and irrigated treatments at Roseworthy. The timing of N application did not have a significant effect on total crop water use, water-use efficiency
, total N uptake or N-use efficiency. The results are consistent with earlier work which found similar yields for N applied at sowing or split between sowing and the first appearance of buds (GS51) [
6,
25]. Therefore, in terms of N management in the medium-high rainfall zone, estimating and applying the correct rate of N is more critical to yield and NUE than the time of application.
The improvements in yield from additional N were due to changes in crop dry matter because there was very little difference in the HI among the treatments or between the two sites. Interestingly, irrigation at the rosette stage at Roseworthy improved yield by 49% with an increase of 41% in total shoot dry matter without any considerable improvement in HI, which highlights the importance of crop biomass production to yield.
The independence of yield from HI in canola observed in this study was also reported for different
Brassica species by Lewis and Thurling [
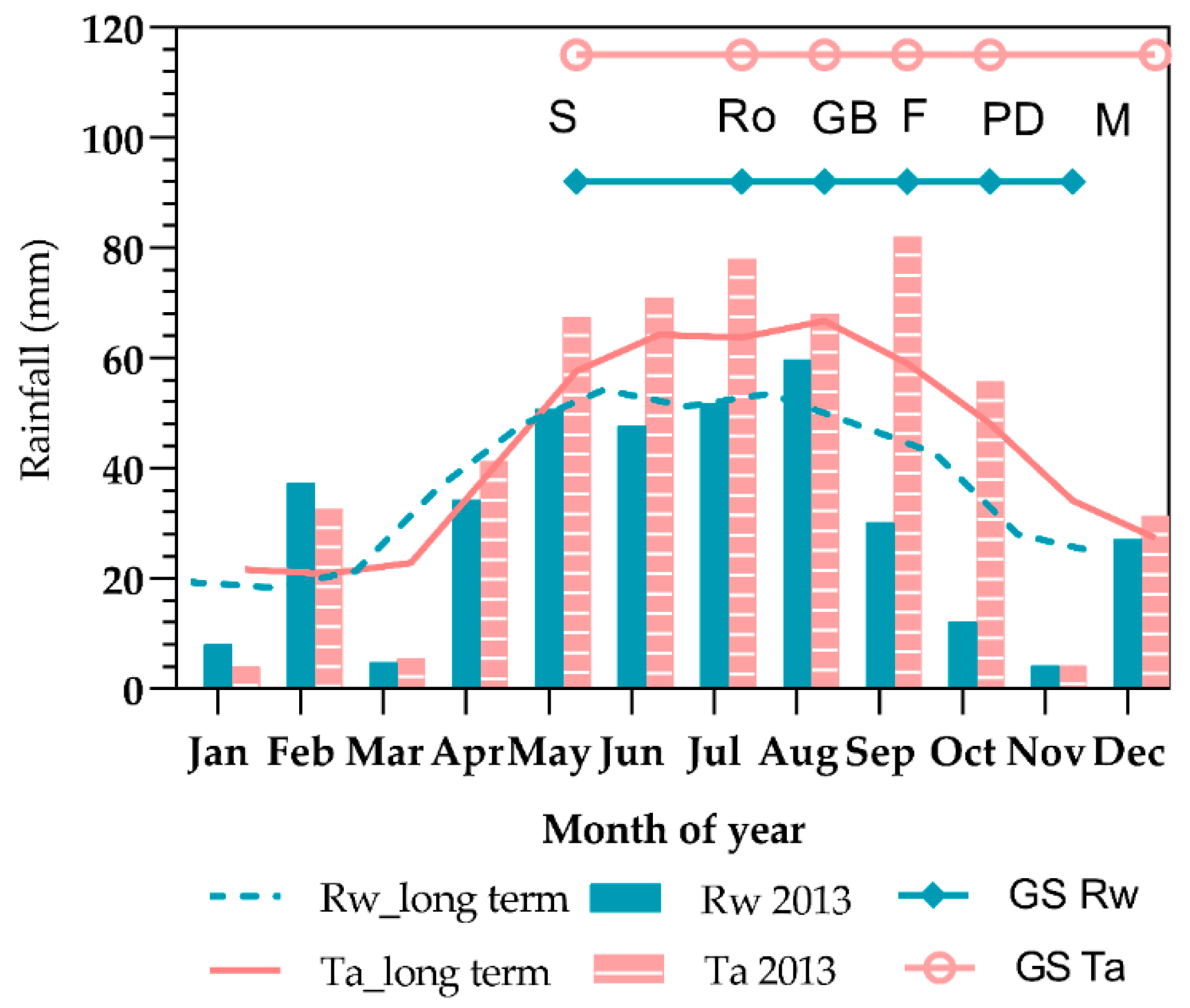
48]. Differences in total dry matter and yield between the two sites and different water regimes were attributed not only to water availability, but also to the different rainfall patterns. Tarlee received around 30% more rainfall than Roseworthy from sowing to mid-flowering period (May–August), but rainfall was 2.5 times higher during the late-flowering and pod initiation period (September) and around three times higher during the pod-development phase (October) (
Figure 1). The lower dry matter and yield for the rainfed treatments at Roseworthy compared to Tarlee was due to lower available soil water during the flowering and pod-development phases, which reduced crop dry matter production and sink development leading to low N uptake and utilization [
49,
50]. Richards and Thurling [
51] also found a significant reduction in canola dry matter and yield components due to water stress at flowering and pod development.
While differences in yield and responses to N were driven more by crop biomass rather than by HI, the values reported in these and other experiments in the region tend to be low and may be limiting yield and responsiveness to N. The average values in the current experiments were 0.24 (Roseworthy) and 0.27 (Tarlee) which are lower than the average HI reported for canola in Australia (0.28; [
52]), but comparable to the average HI for commercial crops in South Australia and western Victoria (0.24) [
53]. By comparison HI for canola in trials from SE New South Wales are often substantially higher; for example, Hocking and Stapper reported HIs of 0.29–0.37 for crops sown in April and May at two sites [
44] and they only fell to values comparable to those reported in the current experiments (0.25–0.28) when the crops were sown late and exposed to a higher level of stress due to the short growing season. Previously, Hocking et al. measured HIs of 0.27–0.34 [
54], while more recent work confirmed this difference with canola grown in South Australia having a HI of 0.27 compared to a HI of 0.34 in an experiment grown at Wagga Wagga in New South Wales [
13]. This was associated with a 25% lower yield at the South Australian site. Both sites received well above-average rainfall and total dry matter production was similar at the two sites (only 7% lower in South Australia). The lower HI for crops in the South Australia and western Victoria indicates that growth during pod set and pod fill may be occurring under higher levels of environmental stress compared to SE New South Wales.
The mean oil concentration for canola was greater than 42% at both the sites. Oil concentration in canola seed was unaffected by the site or irrigation regime. Nitrogen application slightly reduced the oil content of seed at Tarlee compared to the control, which is consistent with the inverse relationship between oil and protein reported in the literature [
24,
54,
55]. Split applications of N improved the oil content compared to the single N application at the high rainfall site, but the difference between the treatments was less than 0.5%. At Roseworthy, split application improved seed oil content in irrigated conditions, but not in rainfed environment. High oil content in the irrigated crop at Roseworthy may be attributed to a longer period of seed development and slower seed maturity than in the rainfed crop due to higher water availability.
Seed yield formation involves complex interactions between yield components and yield can be both sink-limited (related to pod and seed numbers) and source-limited (reflected in seed size) because of the indeterminate growth pattern of canola [
27]. The importance of sink size to yield has been demonstrated in several studies that have reported pod number to be the main factor responsible for yield [
29,
56,
57,
58]. Irrigation increased pod numbers in the Roseworthy experiment (from 6700 to 8200 pods m
−2), but N rate was the more important influence, increasing pods m
−2 × 131% at Roseworthy under rainfed conditions and by 210% under irrigated conditions and by 130% at Tarlee. While producing large number of pods was important to yield, there was no benefit from setting more than 8000 pods m
−2 under the rainfed conditions at either site or more than 10,000 pods m
−1 under irrigation. The limit to the yield from high pod number in the rainfed treatments was due to a reduction in seeds pod
−1, which may reflect the increasing contribution from pods on the branches and the higher level of competition for assimilate at seed set. Seed set in pods on the branches show greater sensitivity to stress than the seed set on the main stem pods [
13].
The strong relationship between seed number and yield for both the rainfed and irrigated treatments at Roseworthy suggests that yields were sink-limited at this site. The dry spring severely curtailed growth during flowering and early pod set, a growth stage critical to seed number and yield [
13,
59] yet additional N still increased seed number. In contrast, there was evidence that the crops at Tarlee were more strongly source-limited. Despite producing more seeds m
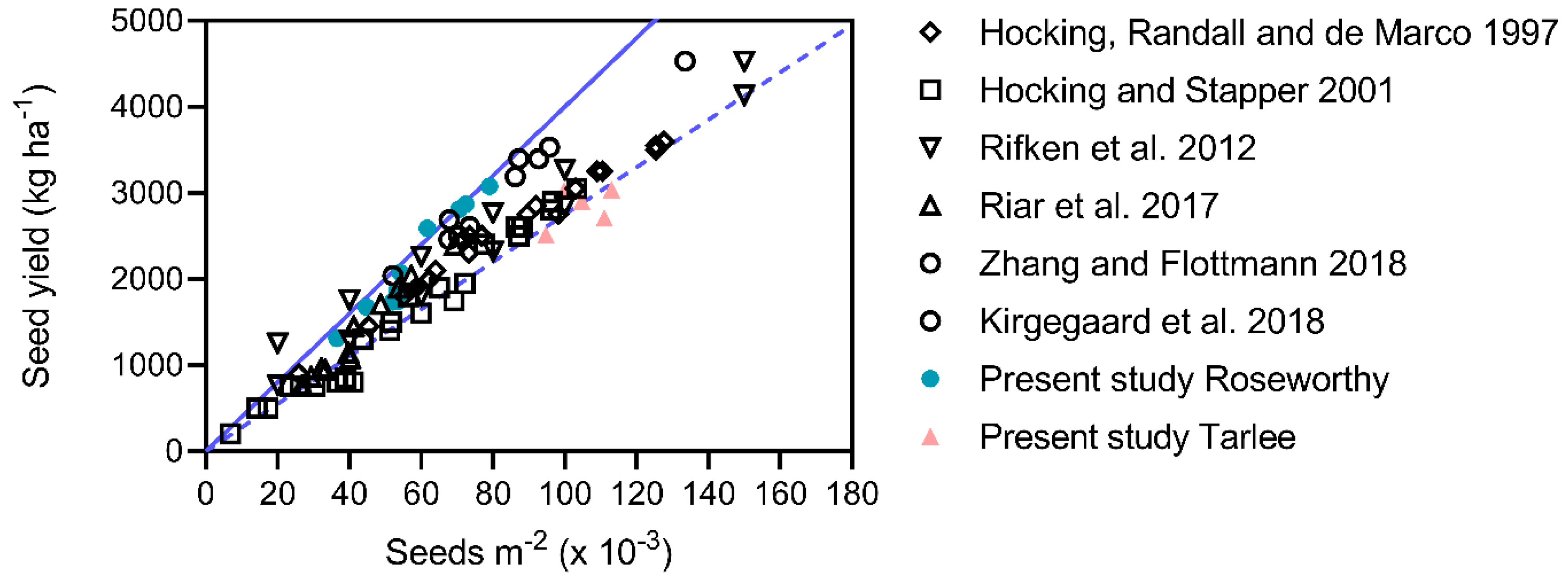
−2 than irrigated canola at Roseworthy, maximum yields were similar because of smaller seed weight. Moreover, the yields at Tarlee are generally lower than those achieved at other sites with comparable seeds m
−2 (
Figure 2).
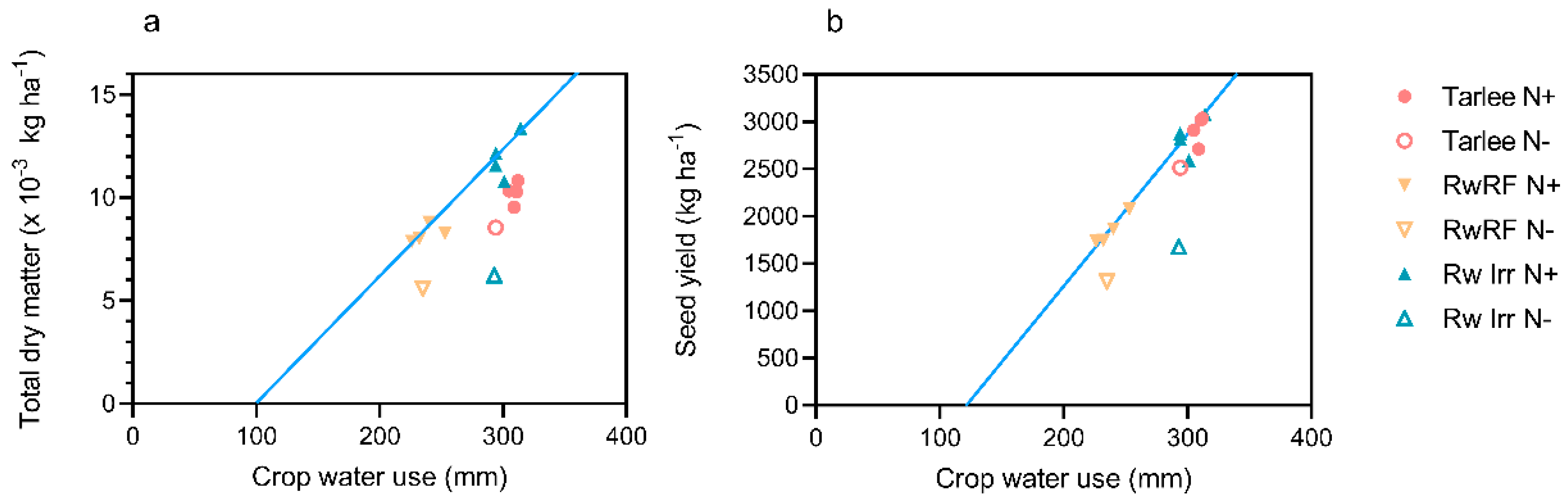
Responses to water and N are linked and improvements in WUE are influenced by N supply. The seasonal marginal WUE
GY indicates the water-limited yield in these experiments. The value derived from the trials (16 kg ha
−1 mm
−1) was not significantly different from the WUE
GY estimated for canola over a much wider range of environments by Houshmandfer et al. (17 ± 1 kg ha
−1 mm
−1) [
16], but higher than earlier reports of WUE
GY by Nuttal and Armstrong (approximately 12 kg ha
−1 mm
−1, based on the marginal WUE in their Figure 7) [
53] and Robertson and Kirkegaard (14 kg ha
−1 mm
−1), suggesting there has been an overall improvement in crop WUE
GY in canola [
60]. However, the current experiments demonstrated that an adequate supply of N is crucial to high WUE
GY and the crop achieving its water-limited yield potential.
The greater response to N with irrigation at Roseworthy compared to the rainfed treatment clearly demonstrated how an increase in available moisture increases crop water use, N recovery, yield and WUE. Dependence of N uptake on plant available water has also been previously reported in wheat [
10]. However, crop water use at Tarlee was the same as the irrigated crop at Roseworthy despite receiving 46% more rainfall than water received from rainfall plus irrigation at Roseworthy. The patterns of water extraction at Tarlee indicated the effective root depth was 60–80 cm and the increase in soil moisture below 80 cm suggested the crops were unable to use available subsoil moisture effectively. Canola roots can grow rapidly in moist soil and have been reported to reach more than 2 m from May-sown crops in deep nonconstrained soils [
61], so potentially this moisture should have been available to the crop if there was no impediment to root growth. However, in soil with subsoil constraints, canola roots may largely be limited to the top 80 cm [
53]. When adequate N was supplied, the WUE at Tarlee was high (
Figure 3) and so arguably the limitation to higher yields at Tarlee and responses to N was the inability to use available soil moisture in the subsoil completely rather than use the available water inefficiently. Kirkegaard et al. reported a marginal water-use efficiency of 38 kg ha
−1 mm
−1 from the additional use of subsoil moisture [
61], which is about four times the seasonal WUE
GY measured at Tarlee, so even improving the depth of soil water extraction to 1 m could potentially have a large effect on yield. Norton and Wachsmann also showed that small changes in crop water use had a large effect on yield in canola [
9]. While a full chemical analysis was not done on the soil at Tarlee, the chemical properties of the nearby reference profile indicates the subsoil is sodic with high chloride and boron concentrations (
Table S1), and the depth of root growth was likely limited by these soil properties.
The subsoil at Roseworthy was alkaline and had a much higher EC than Tarlee. The reference profile also indicates the subsoil also has high boron and is sodic (
Table S1). The dry soil profile during the experiment makes interpretation of the changes in soil moisture difficult, but the high subsoil EC, boron and exchangeable sodium% may have contributed to a lower crop water use. Similar to Tarlee, when N was supplied the WUE was high (
Figure 3), so the major limitation to high yields and WUE was the availability of moisture. However, comparison of water use in the irrigated and rainfed treatments also allowed an estimate of the marginal WUE
GY (the increased in yield per mm of additional water use from irrigation) to be calculated and this suggested that even when the water supply was increased, the soil properties may impose a limit on WUE compared to less constrained soils. Irrigation in winter increased subsoil moisture resulting in a marginal WUE
GY from irrigation of 13.6 kg ha
−1 mm
−1. However, this is only one-third of the value reported for canola crops in New South Wales (38 kg ha
−1 mm
−1), suggesting that while water was being extracted at depth the efficiency with which it was used was low. High salinity can reduce WUE
GY of canola [
62] and surveys of commercial crops indicate that canola is more sensitive to the limitations of sodic and saline soils than wheat or barley [
53]. There has been little work examining the effect of the soil constraints associated with the calcareous and sodic soils in the region on canola growth and water use, but limitations to the ability of crops to extract subsoil water or use it efficiently may be a bottleneck to future improvements in NUE in canola on many of the sodic soils that are common in southern Australia. Poor use of subsoil moisture may also help explain the tendency for crops in the region to have lower HIs since low post flowering water use can reduce HI [
52].
At Roseworthy, irrigation improved N uptake by 1.4-fold compared to the rainfed crop, which improved N uptake efficiency and N-use efficiency by 40%. Furthermore, irrigation resulted in a more than 2.0-fold improvement in agronomic efficiency and N-utilization efficiency. Physiological efficiency, apparent recovery and NHI were unaffected by irrigation suggesting that N uptake is a major limitation to yield and NUE in these environments [
63]. Improving N uptake will not only be influenced by soil moisture, but the ability of the crop to use available soil moisture.







{kind=link}
{kind=link}
{kind=link}
{kind=link}