Distribution of Pigeon Pea Cyst Nematode and Root-Knot Nematodes in Major Sesame Growing Areas in Myanmar

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
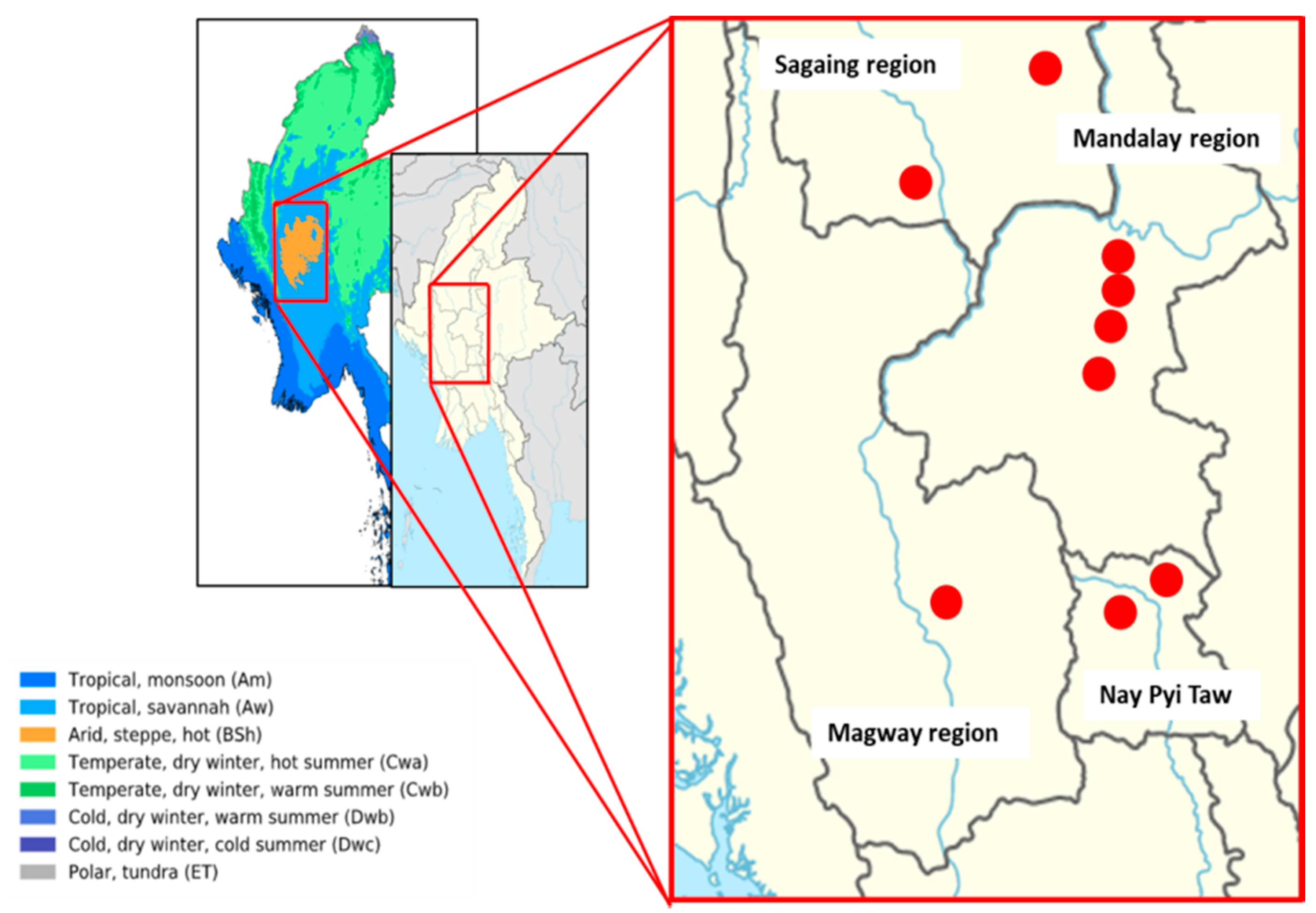
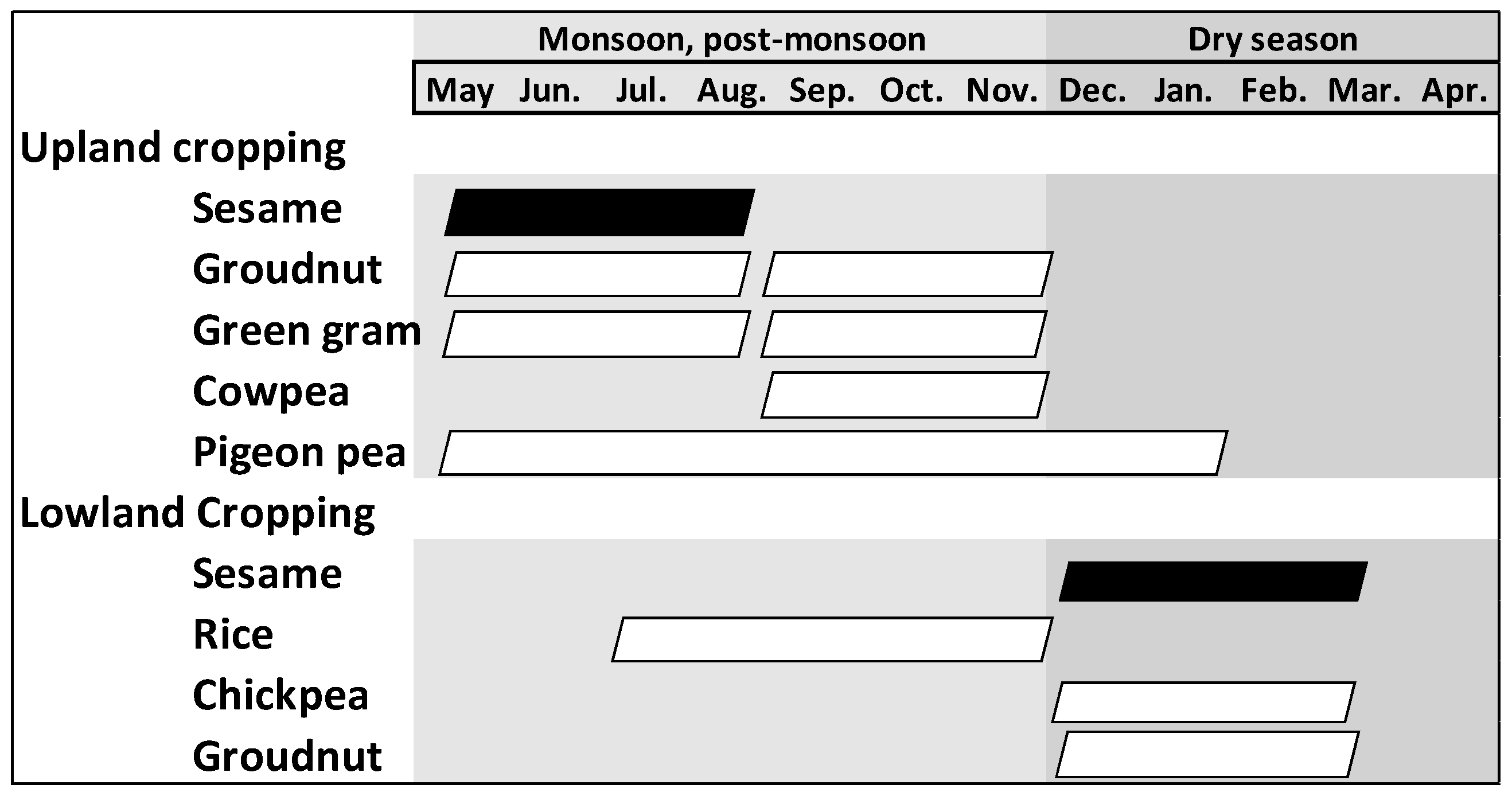
2.1. Description of the Soil Sampling Sites and the Sesame-Based Cropping
2.2. Soil Sampling
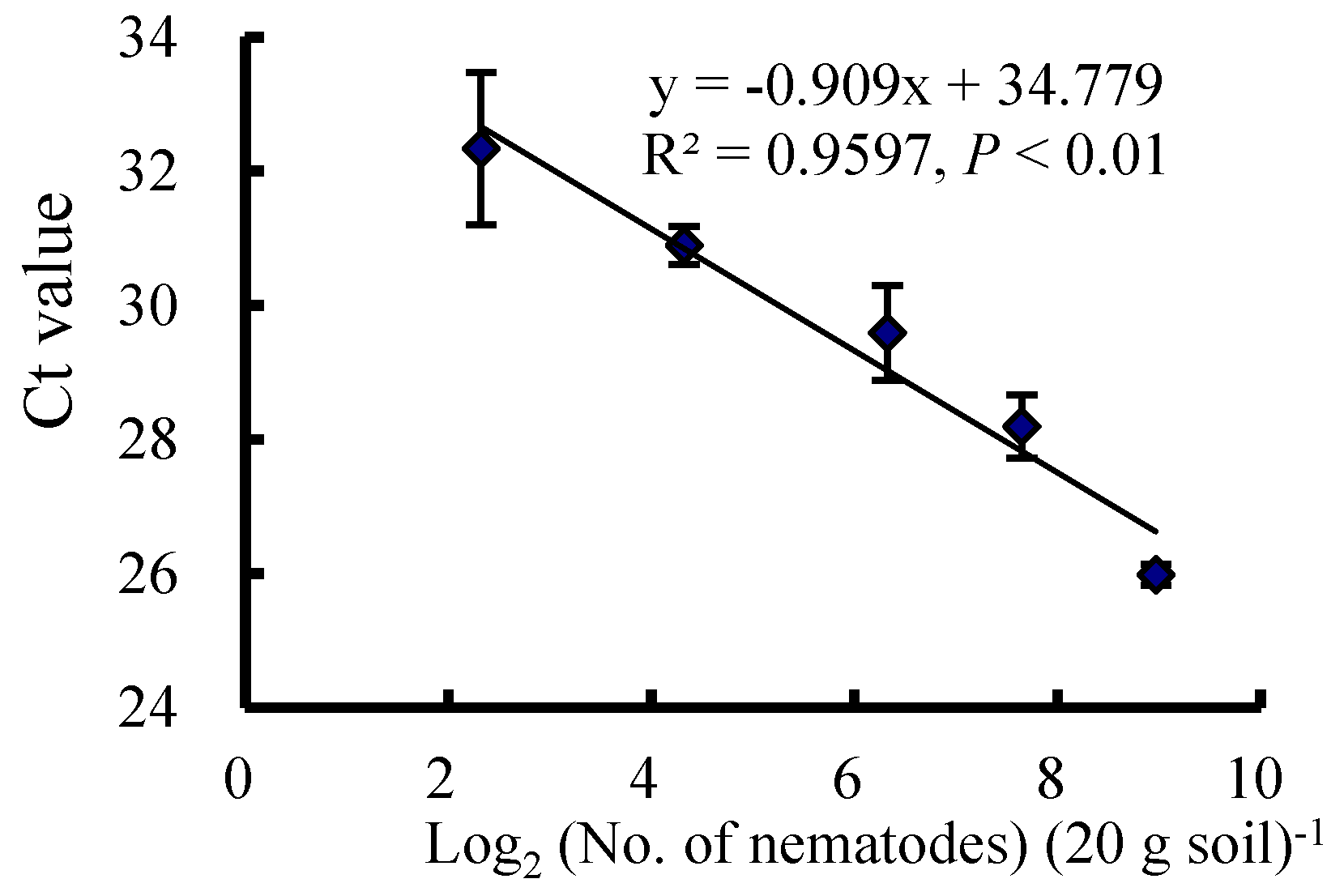
2.3. Nematode Quantification
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- MOALI. Myanmar Agriculture at a Glance 2016; Department of Planning, Ministry of Agriculture, Livestock and Irrigation: Nay Pyi Taw, Myanmar, 2018; 166p. [Google Scholar]
- FAOSTAT. Food and Agricultural Organization of the United Nations Statistics. 2018. Available online: http://www.fao.org/documents/card/en (accessed on 31 August 2019).
- Koshy, P.K.; Swarup, G. Susceptibility of plants to pigeon-pea cyst nematode, Heterodera cajani. Ind. J. Nematol. 1972, 2, 1–6. [Google Scholar]
- Senthamiz, K.; Rajendran, G.; Subramanian, S. Host range, biology and races of Heterodera cajani Koshy, 1967 occurring in Tamil Nadu (India). Ind. J. Nematol. 2005, 2, 187–191. [Google Scholar]
- Sharma, S.B.; Remanandan, P.; Jain, K.C. Resistance to cyst nematode (Heterodera cajani) in pigeonpea cultivars and in wild relatives of Cajanus. Ann. Appl. Biol. 1993, 123, 75–81. [Google Scholar] [CrossRef]
- Myint, Y.Y.; Lwin, T.; Thwe, K.H.; Min, Y.Y.; Aye, S.S.; Lin, M.; Kyi, P.P.; Maung, Z.T.Z.; Than, P.P. New record on the occurrence of cyst nematode, Heterodera cajani Koshy, 1967 on sesame, Sesamum indicum, in Myanmar. In Proceedings of the Fourth Agricultural Research Conference in Commemoration of the Ruby Jubilee of Yezin Agricultural University, Nay Pyi Taw, Myanmar, 17–18 February 2005. [Google Scholar]
- Hadden, R.L. The Geology of Burma: An Annotated Bibliography of Burma’s Geology, Geography and Earth Science; US Army Corps of Engineers: Alexandria, VA, USA, 2008; p. 19. Available online: https://apps.Dtic.mil/dtic/tr/fulltext/u2/a487552.pdf (accessed on 13 September 2020).
- Data Collection on Agriculture Sector in the Republic of the Union of Myanmar. Available online: https://www.themimu.info/sites/themimu.info/files/documents/Assesst_Survey_on_Agriculture_JICA_Dec2013.pdf (accessed on 13 September 2020).
- Myanmar-Climate/Britannica. Available online: https://www.britannica.com/place/Myanmar/Climate (accessed on 30 August 2020).
- Herridge, D.F.; Win, M.M.; New, K.M.M.; Kyu, K.L.; Win, S.S. The cropping systems of the Central Dry Zone of Myanmar: Productivity constraints and possible solutions. Agric. Syst. 2019, 169, 31–40. [Google Scholar] [CrossRef]
- Katsuta, A.; Toyota, K.; Min, Y.Y.; Maung, T.T. Development of real-time PCR primers for the quantification of Meloidogyne graminicola, Hirschmanniella oryzae and Heterodera cajani, pests of the major crops in Myanmar. Nematology 2016, 18, 257–263. [Google Scholar] [CrossRef]
- Invasive Species Compendium. Available online: https://www.cabi.org/isc/datasheet/40612 (accessed on 30 August 2020).
- Köppen–Geiger Climate Classification Map for Myanmar. Available online: https://en.m.wikipedia.org/wiki/File:Koppen-Geiger_Map_MMR_present.svg (accessed on 10 September 2020).
- Thuzar, L. Value Chain Analysis of Sesame in Magway Township; Research Working Paper Series; Paper NO. 5/2013; Mekong Institute, Newzeland Foreign Affair and Trade Aids Programme: Khon Kaen, Thailand, 2013. [Google Scholar]
- Min, Y.Y.; Toyota, K.; Sato, E. A novel nematode diagnostic method using the direct quantification of major plant parasitic in soil by real time PCR. Nematology 2012, 14, 265–276. [Google Scholar] [CrossRef]
- Nemaplex. Available online: http://nemaplex.ucdavis.edu/Uppermnus/topmnu.htm (accessed on 30 August 2020).
- Min, Y.Y.; Toyota, K. Plant-parasitic nematodes in some economically important crops in Myanmar—Species, possible damage and control measures. Nematology 2018, 20, 1–13. [Google Scholar] [CrossRef]
- Toyota, K.; Shirakashi, T.; Sato, E.; Wada, S.; Min, Y.Y. Development of a real-time PCR method for the potato cyst nematode Globodera rostochiensis and the root-knot nematode Meloidogyne incognita. Soil Sci. Plant Nutr. 2008, 54, 72–76. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, M.; Saito, K. Ecological control of Heterodera glycines injurious to azuki bean. Bull. Tochigi Agric. Exp. 1993, 40, 47–58. [Google Scholar]
- Castillo, P.; Navas-Cortés, J.A.; Landa, B.B.; Jiménez-Díaz, R.M.; Nicola, V. Plant-parasitic nematodes attacking chickpea and their in planta interactions with rhizobia and phytopathogenic fungi. Plant Dis. 2008, 92, 840–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viaene, C.N.; Coyne, D.L.; Kerry, B.R. Biological and cultural management. In Plant Nematology; Perry, R.N., Moens, M., Eds.; CABI: Wallingford, UK, 2006; pp. 346–369. [Google Scholar]
- Namu, J.; Karuri, H.; Alakonya, A.; Nyaga, J.; Njeri, E. Distribution of parasitic nematodes in Kenyan rice fields and their relation to edaphic factors, rainfall and temperature. Trop. Plant Pathol. 2018, 43, 128–137. [Google Scholar] [CrossRef]
- Tanda, A.S.; Atwal, A.S.; Bajaj, Y.P.S. In Vitro inhibition of root-knot nematode Meloidogyne incognita by sesame root exudate and its amino acids. Nematologica 1989, 35, 115–124. [Google Scholar] [CrossRef]
- Walker, J.T.; Melin, J.B.; Davis, J. Response of Sesamum indicum and S. radiatum. Accessions to root-knot nematode, Meloidogyne incognita. J. Nematol. 1998, 30, 611–615. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Nematode Species for Sequence Comparison (Accession No.) | Sequences (5′–3′) in the Positions of Specific Primer Set | |
|---|---|---|
| Forward (Mel-f) | Reverse (Mel-r) | |
| Meloidogyne chitwoodi KP901059 | ATCA-ACTT--GACGGGAGCATAATC | CTCG---AGCCGAGAGTGGGTAA |
| M. enterolobii MN661336 | ATCA-ACTT--GACGGGAGCATAATC | CTCG---AGCCGAGAGTGGGTAA |
| M. ethiopica LN626932 | ATCA-ACTT--GACGGGAGCATAATC | CTCG---AGCCGAGAGTGGGTAA |
| M. exigua AF442200 | ATCA-ACTT--GACGGGAGCATAATC | CTCG---AGCCGAGAGTGGGTAA |
| M. floridensis AY942621 | ATCA-ACTT--GACGGGAGCATAATC | CTCG---AGCCGAGAGTGGGTAA |
| M. graminicola MK301102 | ATCA-ACTT--GACGGGAGCATAATC | CTCG---AGCCGAGAGTGGGTAA |
| M. graminis KP901056 | ATCA-ACTT--GACGGGAGCATAATC | CTCG---AGCCGAGAGTGGGTAA |
| M. hapla MH011983 | ATCA-ACTT--GACGGGAGCATAATC | CTCG---AGCCGAGAGTGGGTAA |
| M. incognita MN661338 | ATCA-ACTT--GACGGGAGCATAATC | CTCG---AGCCGAGAGTGGGTAA |
| M. mayaguensis AY942629 | ATCA-ACTT--GACGGGAGCATAATC | CTCG---AGCCGAGAGTGGGTAA |
| M. paranaensis AY942622 | ATCA-ACTT--GACGGGAGCATAATC | CTCG---AGCCGAGAGTGGGTAA |
| M. aberrans MF278756 | ATCA-ACCT--GATGGTAGCATAAAC | CTCG---TGCCGAGAGTGGGTAA |
| M. ichinohei EU669953 | ATCATACTT--GATGTTAGTATCAGT | CTCG---TACCGAGAGTGGGTAA |
| M. duytsi AF442197 | ATCA-ACTT--GACGGGAGCATAACC | CTCG---AGCCGAGAGTGGGTAA |
| M. microtyla AF442198 | ATCA-ACTT--GACGGGAGCATAATT | CTCG---AGCCGAGAGTGGGTAA |
| M. artiellia KC875392 | ATCA-ACTT--GACGGGAGTATAATC | CTCG---TGCCGAGAGTGGGTAA |
| M. mali KJ636400 | ATCA-ACTT--GATGGGAGCATAATT | CTCG---TGCCGAGAGTGGGTAA |
| M. oleae MH011980 | ATCA-ACTT--GACGGGAGCATAACC | CTCG---AACCGAGAGTGGGTAA |
| M. africana KY433422 | ATCA-ACTT--GAAGGGAGCATAATC | CTCT---AGCCGAGAGTGGGTAA |
| M. coffeicola HE667739 | ATCA-ACTT--GAAGGGAGCATAATT | CTCG---TGCCGAGAGTGGGTAA |
| Hirschmanniella oryzae KF366906 | ATCA-ACTTTCGATGGTAGTGTATCT | CTCA---CTCTGAGAGTGGGTAA |
| Ditylenchus destructor AY593912 | ATCA-ACTTTCGATGGTAGTGTATGT | CTCA---TTCTGAGAGTGGGTAA |
| Tylenchus arcuatus EU306349 | ATCA-ACTTTCGATGGTAGTGTATCT | CTCAATGCTCTGAGAGTGGGTAA |
| Hoplolaimus columbus KJ934149 | ATCA-ACTTTCGATGGTAGTATACAG | CTCA---TCTTGAGAGTGGGTAA |
| Pratylenchus neglectus EU130802 | ATCA-ACTTTCGATGGTAGTGTATGT | CTCA---AATCGAGAGTGGGTAA |
| Heterodera glycines AY043247 | ATCA-ACTTTCGATGGTAGTGTATCT | CTCG---CCTTGAGAGTGGGTAA |
| Helicotylenchus dihystera AJ966486 | ATCA-ACTTTCGATGGTAGTGTACCT | CTCG---CCTTGAGAGTGGGTAA |
| Area | The Density * in Each Field ** | No. of Infested Fields (%) | No. of Fields | Average Density * (Range) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Magway | 100 | 0 | 7 | 0 | 61 | 13 | 15 | 0 | 12 | 15 | 12 (63.2) | 19 | 22.6 (0–100) |
| 2 | 0 | 78 | 0 | 0 | 36 | 0 | 13 | 77 | |||||
| Mandalay | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 8 (40.0) | 20 | 9.9 (0–125) |
| 6 | 1 | 13 | 1 | 0 | 125 | 21 | 25 | 0 | 7 | ||||
| Sagaing | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 (0.0) | 14 | 0.0 (0) |
| 0 | 0 | 0 | 0 | ||||||||||
| Nay Pyi Taw | 0 | 0 | 0 | 26 | 0 | 0 | 0 | 0 | 7 | 0 | 8 (18.6) | 43 | 21.0 (0–223) |
| 1 | 0 | 0 | 0 | 0 | 131 | 0 | 0 | 0 | 0 | ||||
| 0 | 0 | 0 | 0 | 47 | 0 | 0 | 0 | 0 | 180 | ||||
| 166 | 223 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 122 | ||||
| 0 | 0 | 0 | |||||||||||
| Area | No. of Fields Infested with RKN (%) | No. of Fields | Average Density * of RKN (Range) |
|---|---|---|---|
| Magway | 0 (0.0) | 19 | |
| Mandalay | 0 (0.0) | 20 | |
| Sagaing | 0 (0.0) | 14 | |
| Nay Pyi Taw | 11 (25.6) | 43 | 34.3 (0.8–137.2) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Min, Y.Y.; Naing, T.H.; Htun, N.N.; Myint, A.K.; Ichinose, Y.; Perry, R.N.; Yoshimura, A.; Toyota, K. Distribution of Pigeon Pea Cyst Nematode and Root-Knot Nematodes in Major Sesame Growing Areas in Myanmar. Agronomy 2020, 10, 1457. https://doi.org/10.3390/agronomy10101457
Min YY, Naing TH, Htun NN, Myint AK, Ichinose Y, Perry RN, Yoshimura A, Toyota K. Distribution of Pigeon Pea Cyst Nematode and Root-Knot Nematodes in Major Sesame Growing Areas in Myanmar. Agronomy. 2020; 10(10):1457. https://doi.org/10.3390/agronomy10101457
Chicago/Turabian StyleMin, Yu Yu, Thu Htet Naing, Nwe Nwe Htun, Aung Kyaw Myint, Yuri Ichinose, Roland N. Perry, Atsushi Yoshimura, and Koki Toyota. 2020. "Distribution of Pigeon Pea Cyst Nematode and Root-Knot Nematodes in Major Sesame Growing Areas in Myanmar" Agronomy 10, no. 10: 1457. https://doi.org/10.3390/agronomy10101457
APA StyleMin, Y. Y., Naing, T. H., Htun, N. N., Myint, A. K., Ichinose, Y., Perry, R. N., Yoshimura, A., & Toyota, K. (2020). Distribution of Pigeon Pea Cyst Nematode and Root-Knot Nematodes in Major Sesame Growing Areas in Myanmar. Agronomy, 10(10), 1457. https://doi.org/10.3390/agronomy10101457