Arbuscular Mycorrhizal Fungi and Associated Microbiota as Plant Biostimulants: Research Strategies for the Selection of the Best Performing Inocula

 ,
,  ,
, {kind=link}
Abstract
1. Introduction
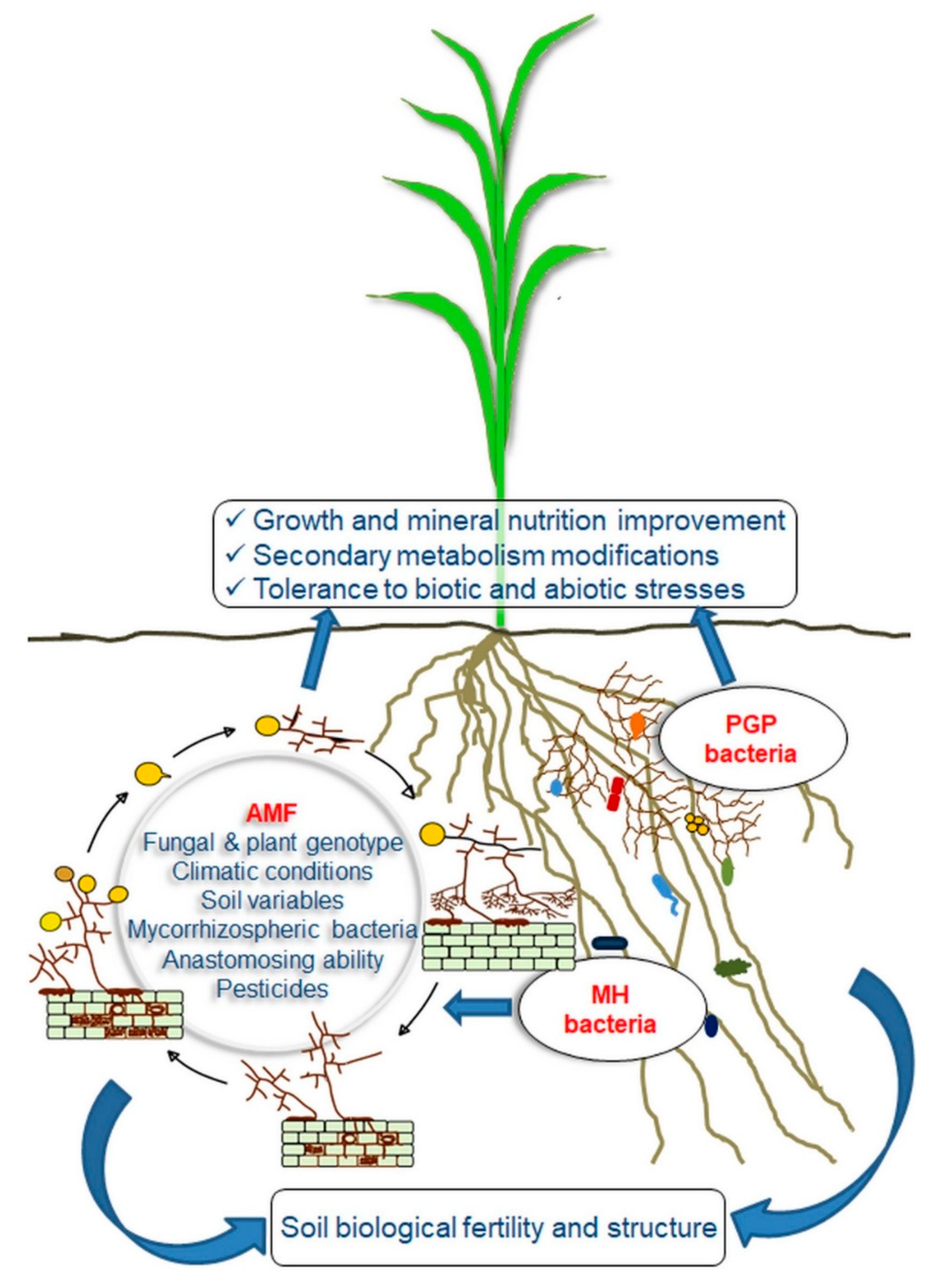
2. Arbuscular Mycorrhizal Fungi
3. AMF Functional Diversity: Colonization Ability and Efficiency
3.1. Colonization Ability
3.2. Efficiency
4. AMF Efficiency in the Enhancement of Plant Health-Promoting Compounds
5. Mycorrhizospheric Bacteria and Their Functional Significance
6. Conclusions and Perspectives for Future Studies
Author Contributions
Funding
Conflicts of Interest
References
- Food and Agriculture Organization. A Policymaker’s Guide to the Sustainable Intensification of Smallholder Crop Production; FAO: Rome, Italy, 2011; Available online: http://www.fao.org/3/a-i2215e.pdf (accessed on 7 January 2020).
- Philippot, L.; Raaijmakers, J.M.; Lemanceau, P.; Van Der Putten, W.H. Going back to the roots: The microbial ecology of the rhizosphere. Nat. Rev. Microbiol. 2013, 11, 789–799. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008. [Google Scholar]
- Casieri, L.; Ait Lahmidi, N.; Doidy, J.; Veneault-Fourrey, C.; Migeon, A.; Bonneau, L.; Courty, P.E.; Garcia, K.; Charbonnier, M.; Delteil, A.; et al. Biotrophic transportome in mutualistic plant-fungal interactions. Mycorrhiza 2013, 23, 597–625. [Google Scholar] [CrossRef]
- Gianinazzi, S.; Gollotte, A.; Binet, M.N.; van Tuinen, D.; Redecker, D.; Wipf, D. Agroecology the key role of arbuscular mycorrhizas in ecosystem services. Mycorrhiza 2010, 20, 519–530. [Google Scholar] [CrossRef]
- Sbrana, C.; Avio, L.; Giovannetti, M. Beneficial mycorrhizal symbionts affecting the production of health-promoting phytochemicals. Electrophoresis 2014, 35, 1535–1546. [Google Scholar] [CrossRef]
- Avio, L.; Turrini, A.; Giovannetti, M.; Sbrana, C. Designing the ideotype mycorrhizal symbionts for the production of healthy food. Front. Plant Sci. 2018, 9, 1089. [Google Scholar] [CrossRef]
- Barea, J.M.; Azcón, R.; Azcón-Aguilar, C. Mycorrhizosphere interactions to improve plant fitness and soil quality. Antonie Leeuwenhoek 2002, 81, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Franken, P.; Schneider, C.; Schwarz, D.; Giovannetti, M.; Agnolucci, M.; De Pascale, S.; Bonini, P.; Colla, G. Arbuscular mycorrhizal fungi act as biostimulants in horticultural crops. Sci. Hortic. 2015, 196, 91–108. [Google Scholar] [CrossRef]
- Turrini, A.; Avio, L.; Giovannetti, M.; Agnolucci, M. Functional complementarity of arbuscular mycorrhizal fungi and associated microbiota: The challenge of translational research. Front. Plant Sci. 2018, 9, 1407. [Google Scholar] [CrossRef] [PubMed]
- Giovannetti, M.; Avio, L. Biotechnology of arbuscular mycorrhizas. In Applied Mycology and Biotechnology: Agriculture and Food Production; Khachatourians, G.G., Arora, D.K., Eds.; Elsevier: Amsterdam, The Netherlands, 2002; Volume 2, pp. 275–310. [Google Scholar]
- Giovannetti, M.; Sbrana, C.; Avio, L.; Citernesi, A.S.; Logi, C. Differential hyphal morphogenesis in arbuscular mycorrhizal fungi during pre-infection stages. New Phytol. 1993, 125, 587–593. [Google Scholar] [CrossRef]
- Akiyama, K.; Matsuzaki, K.I.; Hayashi, H. Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi. Nature 2005, 435, 824–827. [Google Scholar] [CrossRef]
- Sbrana, C.; Giovannetti, M. Chemotropism in the arbuscular mycorrhizal fungus Glomus mosseae. Mycorrhiza 2005, 15, 539–545. [Google Scholar] [CrossRef]
- Kiers, E.T.; Duhamel, M.; Beesetty, Y.; Mensah, J.A.; Franken, O.; Verbruggen, E.; Fellbaum, C.R.; Kowalchuk, G.A.; Hart, M.M.; Bago, A.; et al. Reciprocal rewards stabilize cooperation in the mycorrhizal symbiosis. Science 2011, 333, 880–882. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, W.; Xie, Q.; Liu, N.; Liu, L.; Wang, D.; Zhang, X.; Yang, C.; Chen, X.; Tang, D.; et al. Plants transfer lipids to sustain colonization by mutualistic mycorrhizal and parasitic fungi. Science 2017, 356, 1172–1175. [Google Scholar] [CrossRef]
- Luginbuehl, L.H.; Menard, G.N.; Kurup, S.; Van Erp, H.; Radhakrishnan, G.V.; Breakspear, A.; Oldroyd, G.E.D.; Eastmond, P.J. Fatty acids in arbuscular mycorrhizal fungi are synthesized by the host plant. Science 2017, 356, 1175–1178. [Google Scholar] [CrossRef]
- Smith, F.A.; Smith, S.E. Tansley Review No. 96 structural diversity in (vesicular)–arbuscular mycorrhizal symbioses. New Phytol. 1997, 137, 373–388. [Google Scholar] [CrossRef]
- Giovannetti, M.; Sbrana, C.; Avio, L.; Strani, P. Patterns of below-ground plant interconnections established by means of arbuscular mycorrhizal networks. New Phytol. 2004, 164, 175–181. [Google Scholar] [CrossRef]
- Mikkelsen, B.L.; Rosendahl, S.; Jakobsen, I. Underground resource allocation between individual networks of mycorrhizal fungi. New Phytol. 2008, 180, 890–898. [Google Scholar] [CrossRef]
- Giovannetti, M.; Avio, L.; Sbrana, C. Fungal spore germination and pre-symbiotic mycelial growth–physiological and genetic aspects. In Arbuscular Mycorrhizas: Physiology and Function; Koltai, H., Kapulnik, Y., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 3–32. [Google Scholar] [CrossRef]
- de Novais, C.B.; Sbrana, C.; Júnior, O.J.S.; Siqueira, J.O.; Giovannetti, M. Vegetative compatibility and anastomosis formation within and among individual germlings of tropical isolates of arbuscular mycorrhizal fungi (Glomeromycota). Mycorrhiza 2013, 23, 325–331. [Google Scholar] [CrossRef]
- Roesti, D.; Ineichen, K.; Braissant, O.; Redecker, D.; Wiemken, A.; Aragno, M. Bacteria associated with spores of the arbuscular mycorrhizal fungi Glomus geosporum and Glomus constrictum. Appl. Environ. Microbiol. 2005, 71, 6673–6679. [Google Scholar] [CrossRef]
- Long, L.; Zhu, H.; Yao, Q.; Ai, Y. Analysis of bacterial communities associated with spores of Gigaspora margarita and Gigaspora rosea. Plant Soil 2008, 310, 1–9. [Google Scholar] [CrossRef]
- Agnolucci, M.; Battini, F.; Cristani, C.; Giovannetti, M. Diverse bacterial communities are recruited on spores of different arbuscular mycorrhizal fungal isolates. Biol. Fertil. Soils 2015, 51, 379–389. [Google Scholar] [CrossRef]
- Giovannetti, M.; Turrini, A.; Strani, P.; Sbrana, C.; Avio, L.; Pietrangeli, B. Mycorrhizal fungi in ecotoxicological studies: Soil impact of fungicides, insecticides and herbicides. Prev. Today 2006, 2, 47–61. [Google Scholar]
- Buysens, C.; Dupré de Boulois, H.; Declerck, S. Do fungicides used to control Rhizoctonia solani impact the non-target arbuscular mycorrhizal fungus Rhizophagus irregularis? Mycorrhiza 2015, 25, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Hage-Ahmed, K.; Rosner, K.; Steinkellner, S. Arbuscular mycorrhizal fungi and their response to pesticides. Pest Manag. Sci. 2019, 75, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Sbrana, C.; Fortuna, P.; Giovannetti, M. Plugging into the network: Belowground connections between germlings and extraradical mycelium of arbuscular mycorrhizal fungi. Mycologia 2011, 103, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Giovannetti, M.; Azzolini, D.; Citernesi, A.S. Anastomosis formation and nuclear and protoplasmic exchange in arbuscular mycorrhizal fungi. Appl. Environ. Microbiol. 1999, 65, 5571–5575. [Google Scholar] [CrossRef]
- Voets, L.; De La Providencia, I.E.; Declerck, S. Glomeraceae and Gigasporaceae differ in their ability to form hyphal networks. New Phytol. 2006, 172, 185–188. [Google Scholar] [CrossRef]
- Chiocchio, V.; Venedikian, N.; Martinez, A.E.; Ana Menendez, A.M.; Ocampo, J.A.; Godeas, A. Effect of the fungicide benomyl on spore germination and hyphal length of the arbuscular mycorrhizal fungus Glomus mosseae. Int. Microbiol. 2000, 3, 173–175. [Google Scholar]
- de Novais, C.B.; Giovannetti, M.; de Faria, S.M.; Sbrana, C. Two herbicides, two fungicides and spore-associated bacteria affect Funneliformis mosseae extraradical mycelium structural traits and viability. Mycorrhiza 2019, 29, 341–349. [Google Scholar] [CrossRef]
- Buee, M.; Rossignol, M.; Jauneau, A.; Ranjeva, R.; Bécard, G. The pre-symbiotic growth of arbuscular mycorrhizal fungi is induced by a branching factor partially purified from plant root exudates. Mol. Plant Microbe Interact. 2000, 13, 693–698. [Google Scholar] [CrossRef]
- Giovannetti, M.; Citernesi, A.S. Time-course of appressorium formation on host plants by arbuscular mycorrhizal fungi. Mycol. Res. 1993, 97, 1140–1142. [Google Scholar] [CrossRef]
- Mosse, B. The regular germination of resting spores and some observations on the growth requirements of an Endogone sp. causing vesicular-arbuscular mycorrhiza. Trans. Br. Mycol. Soc. 1959, 42, 273–286. [Google Scholar] [CrossRef]
- Breuninger, M.; Requena, N. Recognition events in AM symbiosis: Analysis of fungal gene expression at the early appressorium stage. Fungal Genet. Biol. 2004, 41, 794–804. [Google Scholar] [CrossRef] [PubMed]
- Turrini, A.; Sbrana, C.; Pitto, L.; Ruffini Castiglione, M.; Giorgetti, L.; Briganti, R.; Bracci, T.; Evangelista, M.; Nuti, M.P.; Giovannetti, M. The antifungal Dm-AMP1 protein from Dahlia merckii expressed in Solanum melongena is released in root exudates and differentially affects pathogenic fungi and mycorrhizal symbiosis. New Phytol. 2004, 163, 393–403. [Google Scholar] [CrossRef]
- Morley, C.D.; Mosse, B. Abnormal vesicular-arbuscular mycorrhizal infections in white clover induced by lupin. Trans. Br. Mycol. Soc. 1976, 67, 510–513. [Google Scholar] [CrossRef]
- Fortuna, P.; Citernesi, S.; Morini, S.; Giovannetti, M.; Loreti, F. Infectivity and effectiveness of different species of arbuscular mycorrhizal fungi in micropropagated plants of Mr S 2/5 plum rootstock. Agronomie 1992, 12, 825–829. [Google Scholar] [CrossRef][Green Version]
- Staehelin, C.; Charon, C.; Boller, T.; Crespi, M.; Kondorosi, Á. Medicago truncatula plants overexpressing the early nodulin gene enod40 exhibit accelerated mycorrhizal colonization and enhanced formation of arbuscules. Proc. Natl. Acad. Sci. USA 2001, 98, 15366–15371. [Google Scholar] [CrossRef]
- Tawaraya, K.; Hashimoto, K.; Wagatsuma, T. Effect of root exudate fractions from P-deficient and P-sufficient onion plants on root colonisation by the arbuscular mycorrhizal fungus Gigaspora margarita. Mycorrhiza 1998, 8, 67–70. [Google Scholar] [CrossRef]
- Turrini, A.; Giordani, T.; Avio, L.; Natali, L.; Giovannetti, M.; Cavallini, A. Large variation in mycorrhizal colonization among wild accessions, cultivars, and inbreds of sunflower (Helianthus annuus L.). Euphytica 2016, 207, 331–342. [Google Scholar] [CrossRef]
- De Vita, P.; Avio, L.; Sbrana, C.; Laidò, G.; Marone, D.; Mastrangelo, A.M.; Cattivelli, L.; Giovannetti, M. Genetic markers associated to arbuscular mycorrhizal colonization in durum wheat. Sci. Rep. 2018, 8, 10612. [Google Scholar] [CrossRef]
- Battini, F.; Grønlund, M.; Agnolucci, M.; Giovannetti, M.; Jakobsen, I. Facilitation of phosphorus uptake in maize plants by mycorrhizosphere bacteria. Sci. Rep. 2017, 7, 4686. [Google Scholar] [CrossRef] [PubMed]
- Giovannetti, M.; Fortuna, P.; Citernesi, A.S.; Morini, S.; Nuti, M.P. The occurrence of anastomosis formation and nuclear exchange in intact arbuscular mycorrhizal networks. New Phytol. 2001, 151, 717–724. [Google Scholar] [CrossRef]
- Friese, C.F.; Allen, M.F. The spread of VA mycorrhizal fungal hyphae in the soil: Inoculum types and external hyphal architecture. Mycologia 1991, 83, 409–418. [Google Scholar] [CrossRef]
- Bago, B.; Azcón-Aguilar, C.; Goulet, A.; Piché, Y. Branched absorbing structures (BAS): A feature of the extraradical mycelium of symbiotic arbuscular mycorrhizal fungi. New Phytol. 1998, 139, 375–388. [Google Scholar] [CrossRef]
- Giovannetti, M.; Avio, L.; Sbrana, C. Functional significance of anastomosis in arbuscular mycorrhizal networks. In Mycorrhizal Networks; Horton, T., Ed.; Springer: Dordrecht, The Netherlands, 2015; Volume 224, pp. 41–67. [Google Scholar]
- Purin, S.; Morton, J.B. In situ analysis of anastomosis in representative genera of arbuscular mycorrhizal fungi. Mycorrhiza 2011, 21, 505–514. [Google Scholar] [CrossRef]
- Purin, S.; Morton, J.B. Anastomosis behavior differs between asymbiotic and symbiotic hyphae of Rhizophagus clarus. Mycologia 2013, 105, 589–602. [Google Scholar] [CrossRef]
- Pepe, A.; Giovannetti, M.; Sbrana, C. Different levels of hyphal self incompatibility modulate interconnectedness of mycorrhizal networks in three arbuscular mycorrhizal fungi within the Glomeraceae. Mycorrhiza 2016, 26, 325–332. [Google Scholar] [CrossRef]
- Avio, L.; Pellegrino, E.; Bonari, E.; Giovannetti, M. Functional diversity of arbuscular mycorrhizal fungal isolates in relation to extraradical mycelial networks. New Phytol. 2006, 172, 347–357. [Google Scholar] [CrossRef]
- Helgason, T.; Daniell, T.J.; Husband, R.; Fitter, A.H.; Young, J.P.W. Ploughing up the wood-wide web? Nature 1998, 394, 431. [Google Scholar] [CrossRef]
- Simard, S.W.; Durrell, D.M. Mycorrhizal networks: A review of their extent, function and importance. Can. J. Bot. 2004, 82, 1140–1165. [Google Scholar] [CrossRef]
- Avio, L.; Castaldini, M.; Fabiani, A.; Bedini, S.; Sbrana, C.; Turrini, A.; Giovannetti, M. Impact of nitrogen fertilization and soil tillage on arbuscular mycorrhizal fungal communities in a Mediterranean agroecosystem. Soil Biol. Biochem. 2013, 67, 285–294. [Google Scholar] [CrossRef]
- Njeru, E.M.; Avio, L.; Sbrana, C.; Turrini, A.; Bocci, G.; Bàrberi, P.; Giovannetti, M. First evidence for a major cover crop effect on arbuscular mycorrhizal fungi and organic maize growth. Agron. Sustain. Dev. 2014, 34, 841–848. [Google Scholar] [CrossRef]
- de Novais, C.B.; Avio, L.; Giovannetti, M.; de Faria, S.M.; Siqueira, J.O.; Sbrana, C. Interconnectedness, length and viability of arbuscular mycorrhizal mycelium as affected by selected herbicides and fungicides. Appl. Soil Ecol. 2019, 143, 144–152. [Google Scholar] [CrossRef]
- Schubert, A.; Marzachi, C.; Mazzitelli, M.; Cravero, M.C.; Bonfante-Fasolo, P. Development of total and viable extraradical mycelium in the vesicular–arbuscular mycorrhizal fungus Glomus clarum Nicol. & Schenck. New Phytol. 1987, 107, 183–190. [Google Scholar] [CrossRef]
- Hamel, C.; Fyles, H.; Smith, D.L. Measurement of development of endomycorrhizal mycelium using three different vital stains. New Phytol. 1990, 115, 297–302. [Google Scholar] [CrossRef]
- Pepe, A.; Giovannetti, M.; Sbrana, C. Lifespan and functionality of mycorrhizal fungal mycelium are uncoupled from host plant lifespan. Sci. Rep. 2018, 8, 10235. [Google Scholar] [CrossRef]
- Sanders, F.E.; Tinker, P.B. Phosphate flow into mycorrhizal roots. Pestic. Sci. 1973, 4, 385–395. [Google Scholar] [CrossRef]
- Ferrol, N.; Azcón-Aguilar, C.; Pérez-Tienda, J. Arbuscular mycorrhizas as key players in sustainable plant phosphorus acquisition: An overview on the mechanisms involved. Plant Sci. 2018, 280, 441–447. [Google Scholar] [CrossRef]
- Pepe, A.; Sbrana, C.; Ferrol, N.; Giovannetti, M. An in vivo whole-plant experimental system for the analysis of gene expression in extraradical mycorrhizal mycelium. Mycorrhiza 2017, 7, 659–668. [Google Scholar] [CrossRef]
- Johnson, I.T. Glucosinolates: Bioavailability and importance to health. Int. J. Vitam. Nutr. Res. 2002, 71, 26–31. [Google Scholar] [CrossRef]
- Duthie, S.J. Plant polyphenols in cancer and heart disease: Implications as nutritional antioxidants. Nutr. Res. Rev. 2000, 13, 79–106. [Google Scholar] [CrossRef] [PubMed]
- Lund, E. Non-nutritive bioactive constituents of plants: Dietary sources and health benefits of glucosinolates. Int. J. Vitam. Nutr. Res. 2003, 73, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Copetta, A.; Lingua, G.; Berta, G. Effects of three AM fungi on growth, distribution of glandular hairs, and essential oil production in Ocimum basilicum L. var. Genovese. Mycorrhiza 2006, 16, 485–494. [Google Scholar] [CrossRef]
- Toussaint, J.P.; Kraml, M.; Nell, M.; Smith, S.E.; Smith, F.A.; Steinkellner, S.; Schmiderer, C.; Vierheilig, H.; Novak, J. Effect of Glomus mosseae on concentrations of rosmarinic and caffeic acids and essential oil compounds in basil inoculated with Fusarium oxysporum f. sp. basilici. Plant Pathol. 2008, 57, 1109–1116. [Google Scholar] [CrossRef]
- Battini, F.; Bernardi, R.; Turrini, A.; Agnolucci, M.; Giovannetti, M. Rhizophagus intraradices or its associated bacteria affect gene expression of key enzymes involved in the rosmarinic acid biosynthetic pathway of basil. Mycorrhiza 2016, 26, 699–707. [Google Scholar] [CrossRef] [PubMed]
- Gualandi, R.J., Jr. Fungal Endophytes Enhance Growth and Production of Natural Products in Echinacea Purpurea (Moench.). Master’s Thesis, University of Tennessee, Knoxville, TN, USA, 2010. Available online: https://trace.tennessee.edu/utk_gradthes/713 (accessed on 7 January 2020).
- Tavarini, S.; Passera, B.; Martini, A.; Avio, L.; Sbrana, C.; Giovannetti, M.; Angelini, L.G. Plant growth, steviol glycosides and nutrient uptake as affected by arbuscular mycorrhizal fungi and phosphorous fertilization in Stevia rebaudiana Bert. Ind. Crops Prod. 2018, 111, 899–907. [Google Scholar] [CrossRef]
- Zubek, S.; Stojakowska, A.; Anielska, T.; Turnau, K. Arbuscular mycorrhizal fungi alter thymol derivative contents of Inula ensifolia L. Mycorrhiza 2010, 20, 497–504. [Google Scholar] [CrossRef]
- Rambelli, A. The rhizosphere of mycorrhizae. In Ectomycorrhizae: Their Ecology and Physiology; Marks, G.C., Kozlowski, T.T., Eds.; Academic Press: New York, NY, USA, 1973; pp. 299–343. [Google Scholar]
- Walley, F.L.; Germida, J.J. Failure to decontaminate Glomus clarum NT4 spores is due to spore wall-associated bacteria. Mycorrhiza 1996, 6, 43–49. [Google Scholar] [CrossRef]
- Filippi, C.; Bagnoli, G.; Citernesi, A.S.; Giovannetti, M. Ultrastructural spatial distribution of bacteria associated with sporocarps of Glomus mosseae. Symbiosis 1998, 24, 1–12. [Google Scholar]
- Mosse, B. Honey-coloured, sessile Endogone spores: II. Changes in fine structure during spore development. Arch. Mikrobiol. 1970, 74, 129–145. [Google Scholar] [CrossRef]
- MacDonald, R.M.; Chandler, M.R. Bacterium-like organelles in vesicular-arbuscular mycorrizal fungus Glomus caledonium. New Phytol. 1981, 89, 241–246. [Google Scholar] [CrossRef]
- MacDonald, R.M.; Chandler, M.R.; Mosse, B. The occurrence of bacterium-like organelles in vesicular–arbuscular mycorrizal fungi. New Phytol. 1982, 90, 659–663. [Google Scholar] [CrossRef]
- Bianciotto, V.; Bandi, C.D.; Minerdi, M.; Sironi, H.; Tichy, V.; Bonfante, P. An obligately endosymbiotic mycorrhizal fungus itself harbors obligately intracellular bacteria. Appl. Environ. Microbiol. 1996, 62, 3005–3010. [Google Scholar] [CrossRef]
- Mayo, K.; Davis, R.E.; Motta, J. Stimulation of germination of spores of Glomus versiforme by spore-associated bacteria. Mycologia 1986, 78, 426–431. [Google Scholar] [CrossRef]
- Xavier, L.J.C.; Germida, J.J. Bacteria associated with Glomus clarum spores influence mycorrhizal activity. Soil Biol. Biochem. 2003, 35, 471–478. [Google Scholar] [CrossRef]
- Bharadwaj, D.P.; Lundquist, P.O.; Persson, P.; Alström, S. Evidence for specificity of cultivable bacteria associated with arbuscular mycorrhizal fungal spores. FEMS Microbiol. Ecol. 2008, 65, 310–322. [Google Scholar] [CrossRef]
- Cruz, A.F.; Horii, S.; Ochiai, S.; Yasuda, A.; Ishii, T. Isolation and analysis of bacteria associated with spores of Gigaspora margarita. J. Appl. Microbiol. 2008, 104, 1711–1717. [Google Scholar] [CrossRef]
- Battini, F.; Cristani, C.; Giovannetti, M.; Agnolucci, M. Multifunctionality and diversity of culturable bacterial communities strictly associated with spores of the plant beneficial symbiont Rhizophagus intraradices. Microbiol. Res. 2016, 183, 68–79. [Google Scholar] [CrossRef]
- Frey-Klett, P.; Garbaye, J.A.; Tarkka, M. The mycorrhiza helper bacteria revisited. New Phytol. 2007, 176, 22–36. [Google Scholar] [CrossRef]
- Mugnier, J.; Mosse, B. Spore germination and viability of a vesicular arbuscular mycorrhizal fungus, Glomus mosseae. Trans. Br. Mycol. Soc. 1987, 88, 411–413. [Google Scholar] [CrossRef]
- Tylka, G.L.; Hussey, R.S.; Roncadori, R.W. Axenic germination of vesicular–arbuscular mycorrhizal fungi: Effects of selected Streptomyces species. Phytopathology 1991, 81, 754–759. [Google Scholar] [CrossRef]
- Carpenter-Boggs, L.; Loynachan, T.E.; Stahl, P.D. Spore germination of Gigaspora margarita stimulated by volatiles of soil-isolated actinomycetes. Soil Biol. Biochem. 1995, 27, 1445–1451. [Google Scholar] [CrossRef]
- Ames, R.N.; Mihara, K.L.; Bayne, H.G. Chitin-decomposing actynomycetes associated with a vesicular–arbuscular mycorrhizal fungus from a calcareous soil. New Phytol. 1989, 111, 67–71. [Google Scholar] [CrossRef]
- Will, M.E.; Sylvia, D.M. Interaction of rhizosphere bacteria, fertilizer, and vesicular-arbuscular mycorrhizal fungi with sea oats. Appl. Environ. Microbiol. 1990, 56, 2073–2079. [Google Scholar] [CrossRef]
- Calvet, C.; Barea, J.M.; Pera, J. In vitro interactions between the vesicular-arbuscular mycorrhizal fungus Glomus mosseae and some saprophytic fungi isolated from organic substrates. Soil Biol. Biochem. 1992, 24, 775–780. [Google Scholar] [CrossRef]
- Hildebrandt, U.; Ouziad, F.; Marner, F.-J.J.; Bothe, H. The bacterium Paenibacillus validus stimulates growth of the arbuscular mycorrhizal fungus Glomus intraradices up to the formation of fertile spores. FEMS Microbiol. Lett. 2006, 254, 258–267. [Google Scholar] [CrossRef]
- Pivato, B.; Offre, P.; Marchelli, S.; Barbonaglia, B.; Mougel, C.; Lemanceau, P. Bacterial effects on arbuscular mycorrhizal fungi and mycorrhiza development as influenced by the bacteria, fungi, and host plant. Mycorrhiza 2009, 19, 81–90. [Google Scholar] [CrossRef]
- Bidondo, L.F.; Silvani, V.; Colombo, R.; Pérgola, M.; Bompadre, J.; Godeas, A. Pre-symbiotic and symbiotic interactions between Glomus intraradices and two Paenibacillus species isolated from AM propagules. In vitro and in vivo assays with soybean (AG043RG) as plant host. Soil Biol. Biochem. 2011, 43, 1866–1872. [Google Scholar] [CrossRef]
- Ordoñez, Y.M.; Fernandez, B.R.; Lara, L.S.; Rodriguez, A.; Uribe-Vélez, D.; Sanders, I.R. Bacteria with phosphate solubilizing capacity alter mycorrhizal fungal growth both inside and outside the root and in the presence of native microbial communities. PLoS ONE 2016, 11, e0154438. [Google Scholar] [CrossRef]
- Ravnskov, S.; Jakobsen, I. Effects of Pseudomonas fluorescens DF57 on growth and P uptake of two arbuscular mycorrhizal fungi in symbiosis with cucumber. Mycorrhiza 1999, 8, 329–334. [Google Scholar] [CrossRef]
- Ravnskov, S.; Larsen, J.; Jakobsen, I. Phosphorus uptake of an arbuscular mycorrhizal fungus is not effected by the biocontrol bacterium Burkholderia cepacia. Soil Biol. Biochem. 2002, 34, 1875–1881. [Google Scholar] [CrossRef]
- Zhang, L.; Fan, J.; Ding, X.; He, X.; Zhang, F.; Feng, G. Hyphosphere interactions between an arbuscular mycorrhizal fungus and a phosphate solubilizing bacterium promote phytate mineralization in soil. Soil Biol. Biochem. 2014, 74, 177–183. [Google Scholar] [CrossRef]
- Khalid, A.; Arshad, M.; Kahir, Z.A. Screening plant growth promoting rhizobacteria for improving growth and yield of wheat. J. Appl. Microbiol. 2004, 96, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Aloni, R.; Aloni, E.; Langhans, M.; Ullrich, C.I. Role of cytokinin and auxin in shaping root architecture: Regulating vascular differentiation, lateral root initiation, root apical dominance and root gravitropism. Ann. Bot. 2006, 97, 883–893. [Google Scholar] [CrossRef] [PubMed]
- Duca, D.; Lorv, J.; Patten, C.L.; Rose, D.; Glick, B.R. Indole-3- acetic acid in plant–microbe interactions. Antonie Leeuwenhoek 2014, 106, 85–125. [Google Scholar] [CrossRef] [PubMed]
- Budi, S.W.; van Tuinen, D.; Martinotti, G.; Gianinazzi, S. Isolation from Sorghum bicolor mycorrhizosphere of a bacterium compatible with arbuscular mycorrhiza development and antagonistic towards soil-borne fungal pathogens. Appl. Environ. Microbiol. 1999, 65, 5148–5150. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Ravnskov, S.; Xie, G.; Larsen, J. Biocontrol of Pythium damping-off in cucumber by arbuscular mycorrhiza-associated bacteria from the genus Paenibacillus. Biocontrol 2007, 52, 863–875. [Google Scholar] [CrossRef]
- Bharadwaj, D.P.; Lundquist, P.O.; Alström, S. Arbuscular mycorrhizal fungal spore-associated bacteria affect mycorrhizal colonization, plant growth and potato pathogens. Soil Biol. Biochem. 2008, 40, 2494–2501. [Google Scholar] [CrossRef]
- Kobayashi, T.; Nishizawa, N.K. Iron uptake, translocation, and regulation in higher plants. Ann. Rev. Plant Biol. 2012, 63, 131–152. [Google Scholar] [CrossRef]
- Robin, A.; Vansuyt, G.; Hinsinger, P.; Meyer, J.M.; Briat, J.F.; Lemanceau, P. Iron dynamics in the rhizosphere: Consequences for plant health and nutrition. Adv. Agron. 2008, 99, 183–225. [Google Scholar] [CrossRef]
- Agnolucci, M.; Avio, L.; Pepe, A.; Turrini, A.; Cristani, C.; Bonini, P.; Cirino, V.; Colosimo, F.; Ruzzi, M.; Giovannetti, M. Bacteria associated with a commercial mycorrhizal inoculum: Community composition and multifunctional activity as assessed by Illumina sequencing and culture-dependent tools. Front. Plant Sci. 2018, 9, 1956. [Google Scholar] [CrossRef] [PubMed]
- Le Cocq, K.; Gurr, S.J.; Hirsch, P.R.; Mauchline, T.H. Exploitation of endophytes for sustainable agricultural intensification. Mol. Plant Pathol. 2017, 18, 469–473. [Google Scholar] [CrossRef]
- Agnolucci, M.; Palla, M.; Cristani, C.; Cavallo, N.; Giovannetti, M.; De Angelis, M.; Gobbetti, M.; Minervini, F. Beneficial Plant Microorganisms Affect the Endophytic Bacterial Communities of Durum Wheat Roots as Detected by Different Molecular Approaches. Front. Microbiol. 2019, 10, 2500. [Google Scholar] [CrossRef] [PubMed]
- Compant, S.; Clément, C.; Sessitsch, A. Plant growth-promoting bacteria in the rhizo- and endosphere of plants: Their role, colonization, mechanisms involved and prospects for utilization. Soil Biol. Biochem. 2010, 42, 669–678. [Google Scholar] [CrossRef]
- Hardoim, P.R.; van Overbeek, P.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The hidden world within plants: Ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [PubMed]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giovannini, L.; Palla, M.; Agnolucci, M.; Avio, L.; Sbrana, C.; Turrini, A.; Giovannetti, M. Arbuscular Mycorrhizal Fungi and Associated Microbiota as Plant Biostimulants: Research Strategies for the Selection of the Best Performing Inocula. Agronomy 2020, 10, 106. https://doi.org/10.3390/agronomy10010106
Giovannini L, Palla M, Agnolucci M, Avio L, Sbrana C, Turrini A, Giovannetti M. Arbuscular Mycorrhizal Fungi and Associated Microbiota as Plant Biostimulants: Research Strategies for the Selection of the Best Performing Inocula. Agronomy. 2020; 10(1):106. https://doi.org/10.3390/agronomy10010106
Chicago/Turabian StyleGiovannini, Luca, Michela Palla, Monica Agnolucci, Luciano Avio, Cristiana Sbrana, Alessandra Turrini, and Manuela Giovannetti. 2020. "Arbuscular Mycorrhizal Fungi and Associated Microbiota as Plant Biostimulants: Research Strategies for the Selection of the Best Performing Inocula" Agronomy 10, no. 1: 106. https://doi.org/10.3390/agronomy10010106
APA StyleGiovannini, L., Palla, M., Agnolucci, M., Avio, L., Sbrana, C., Turrini, A., & Giovannetti, M. (2020). Arbuscular Mycorrhizal Fungi and Associated Microbiota as Plant Biostimulants: Research Strategies for the Selection of the Best Performing Inocula. Agronomy, 10(1), 106. https://doi.org/10.3390/agronomy10010106