

Repair Effect of Umbilical Cord Mesenchymal Stem Cells Embedded in Hydrogel on Mouse Insulinoma 6 Cells Injured by Streptozotocin
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cells Culture
2.3. Experimental Grouping
2.4. Repairing Effect of UC-MSCs in Combination with Hydrogel on STZ-Induced Injury in MIN-6 Cells
2.4.1. MDA and LDH Determination
2.4.2. SOD, T-AOC, CAT Determination
2.4.3. Inducible Nitric Oxide Synthase (iNOS), IL-1β, IL-10 Assay
2.4.4. Ki67, Pdx1, MafA Assay
2.4.5. Insulin Measurement
2.4.6. Determination of Glucose Content in Cell Supernatant
2.4.7. Live and Dead Cell Staining
2.5. Statistical Significance
3. Results and Discussion
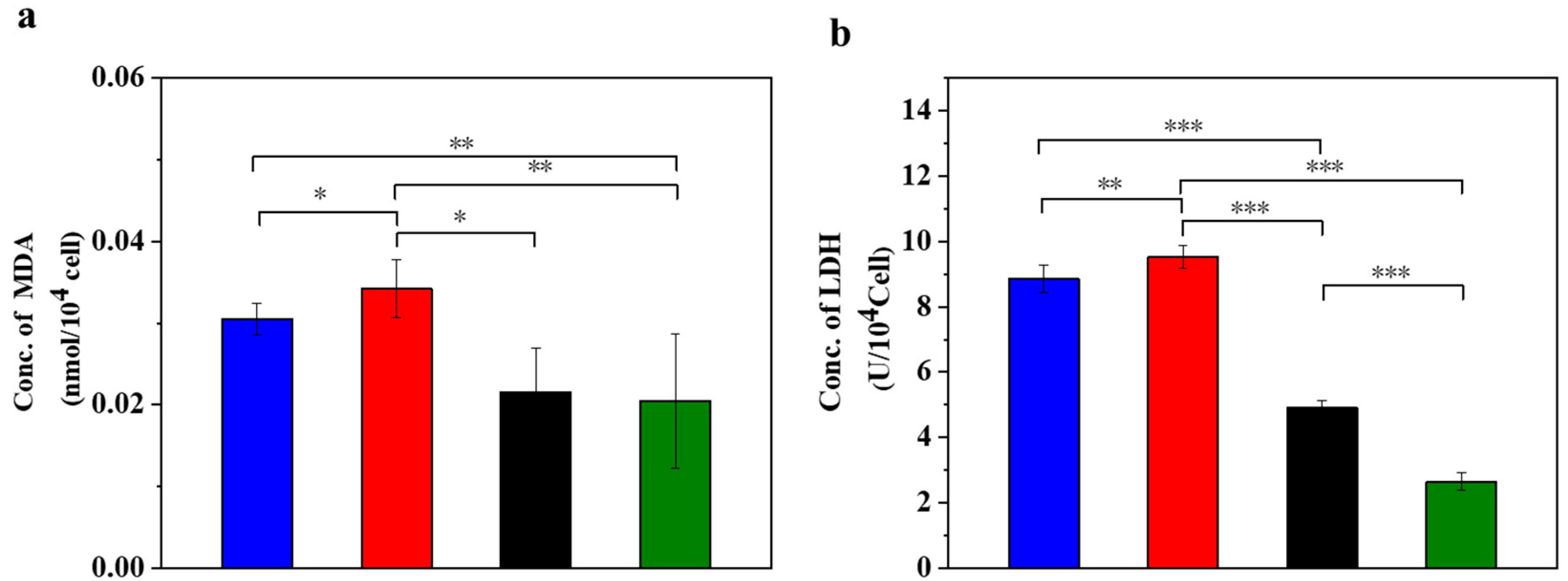
3.1. UC-MSCs + Hydrogel Reduced Oxidative Stress of Injured MIN-6 Cells
3.2. UC-MSCs + Hydrogel Enhanced Antioxidant Capacity of Injured MIN-6 Cells
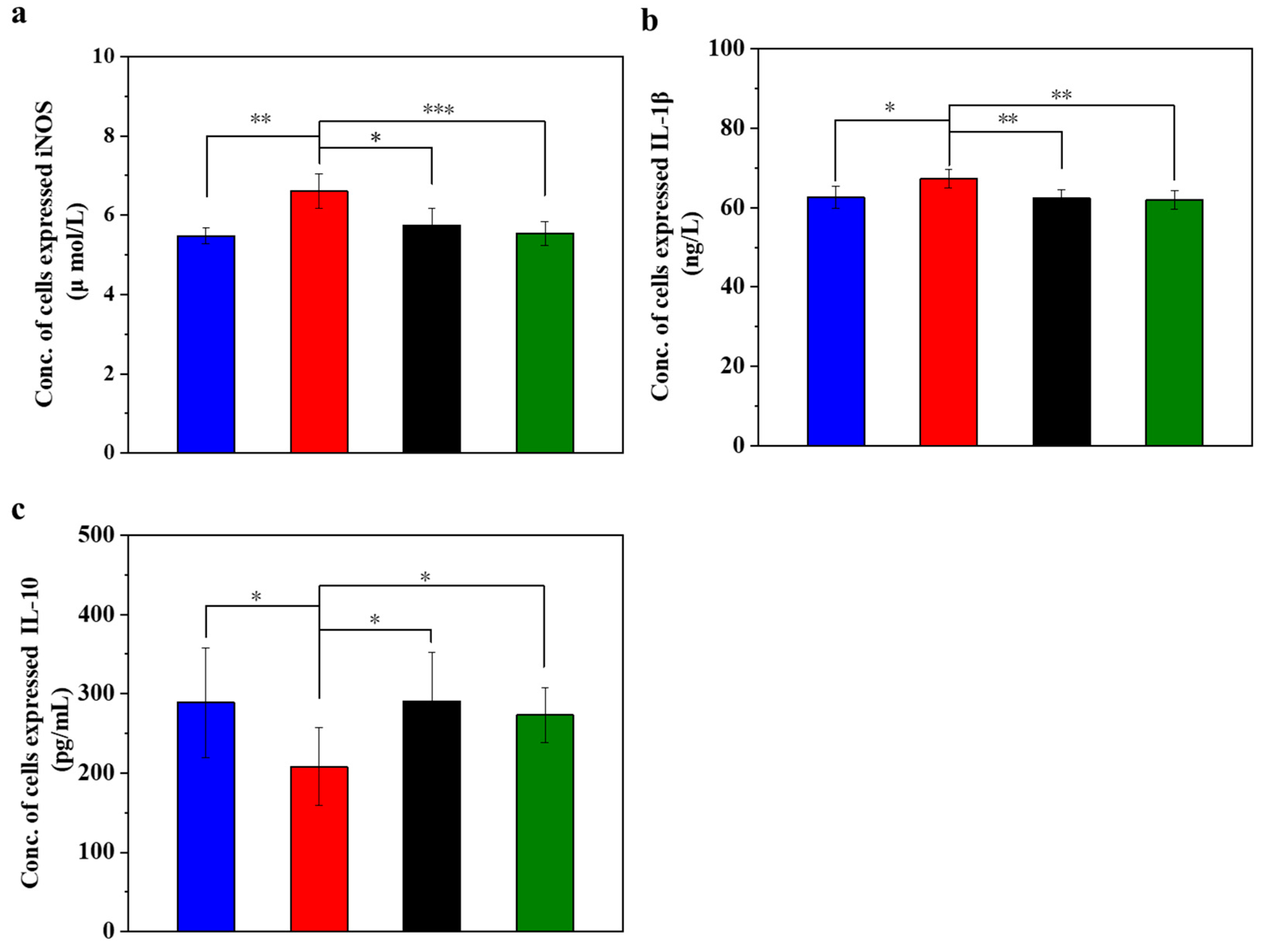
3.3. UC-MSCs + Hydrogel Reduced Inflammation of Injured MIN-6 Cells
3.4. UC-MSCs + Hydrogel Promoted Injured MIN-6 Cells Proliferation
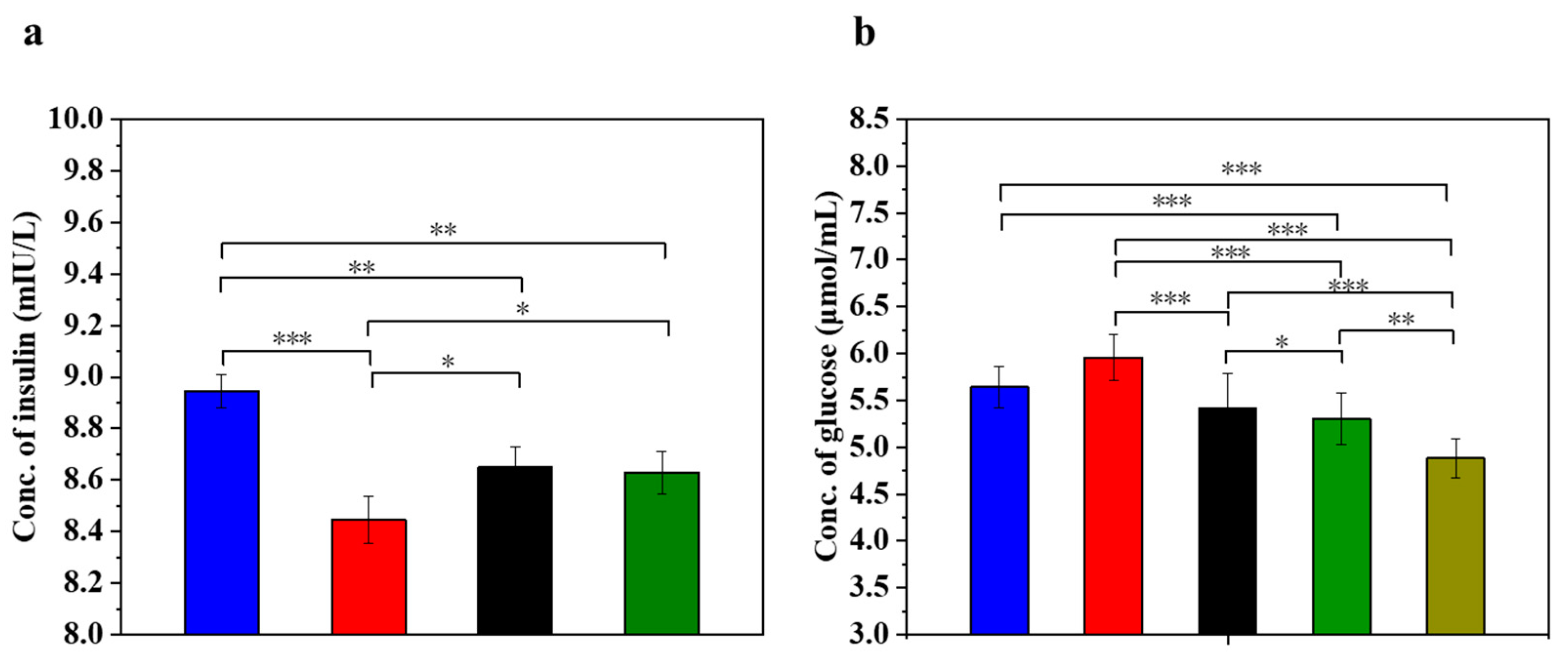
3.5. UC-MSCs + Hydrogel Enhanced Biological Function of Injured MIN-6 Cells
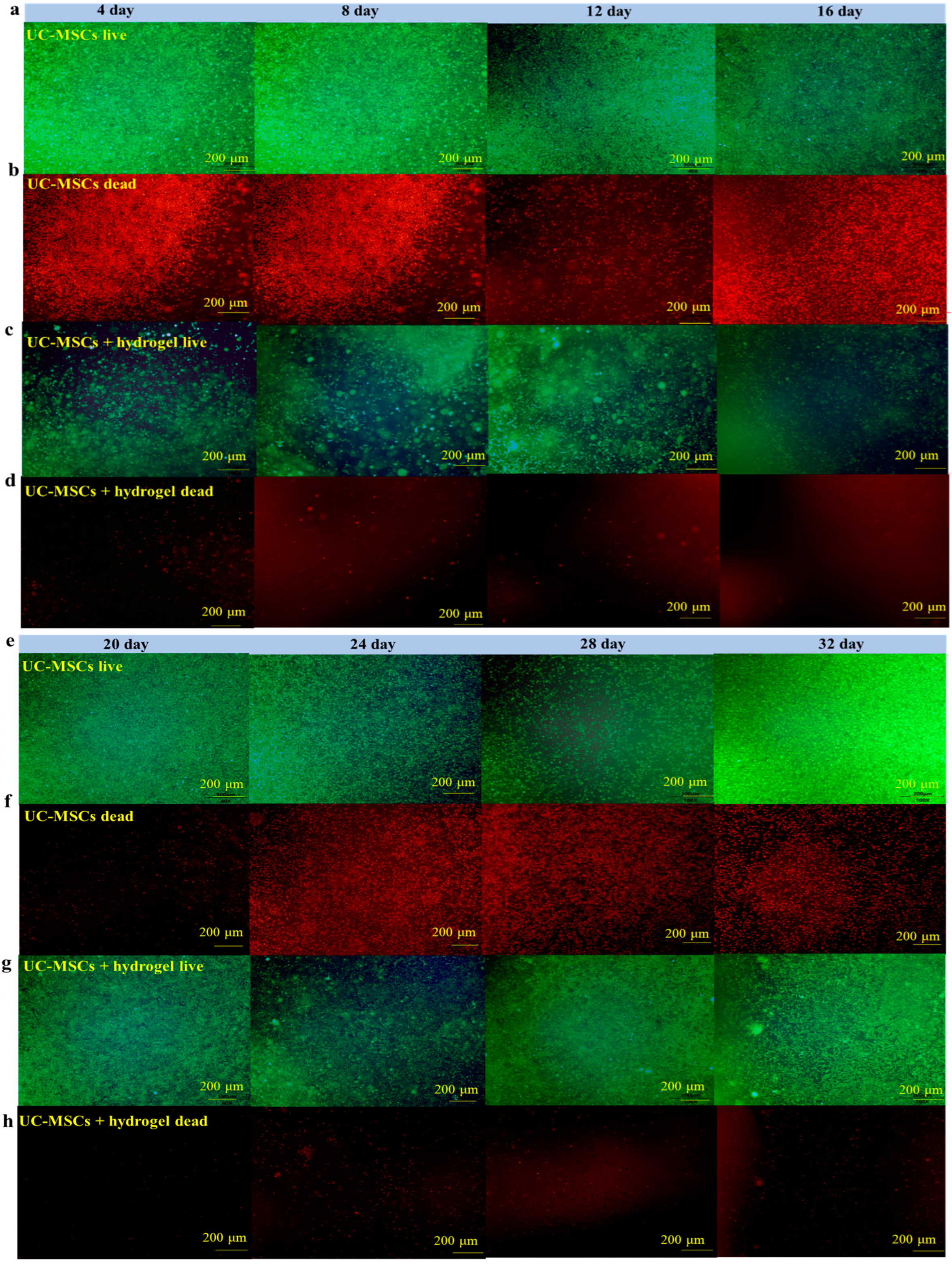
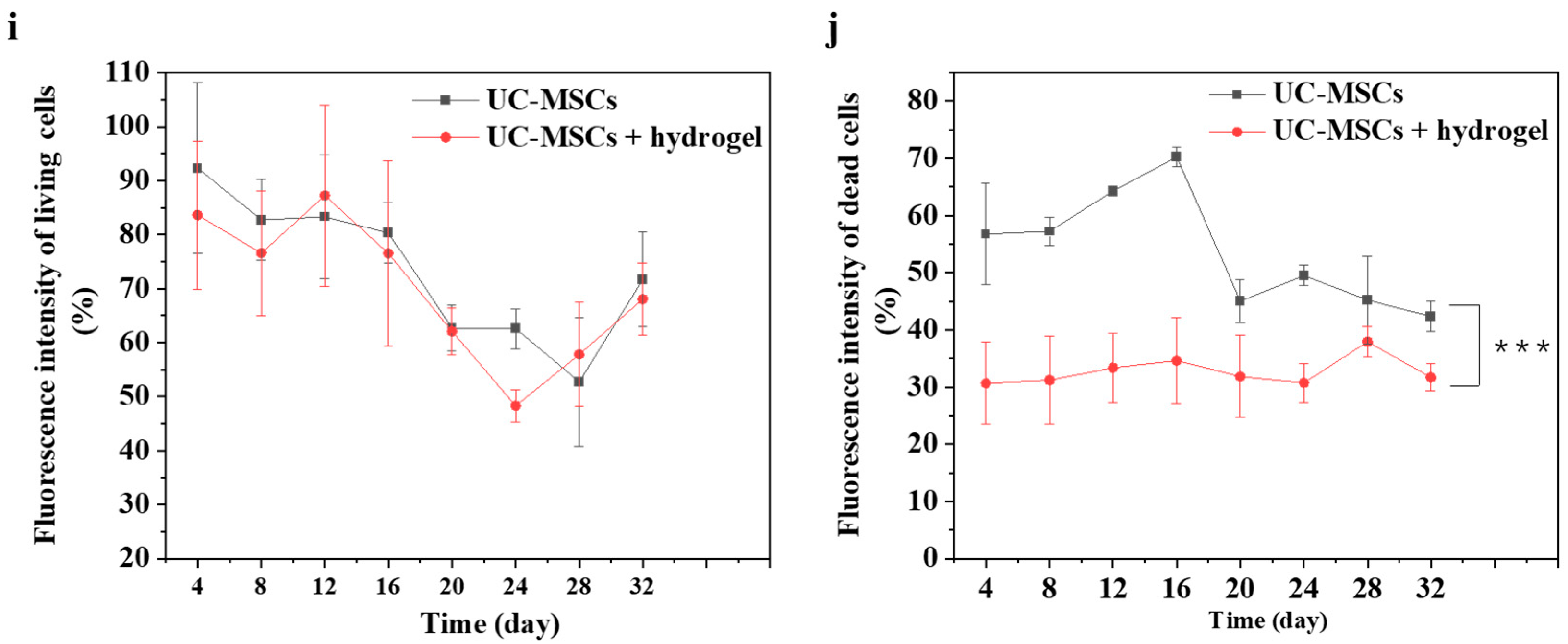
3.6. UC-MSCs + Hydrogel Enhanced Long-Term Survival of Injured MIN-6 Cells
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, H.; Mulya, A.; Nieuwoudt, S.; Vandanmagsar, B.; McDowell, R. GDF15 mediates the effect of skeletal muscle contraction on glucose-stimulated insulin secretion. Diabetes 2023, 72, 1070–1082. [Google Scholar] [CrossRef] [PubMed]
- Allawadhi, P.; Singh, V.; Govindaraj, K.; Khurana, I.; Sarode, L.P. Biomedical applications of polysaccharide nanoparticles for chronic inflammatory disorders: Focus on rheumatoid arthritis, diabetes and organ fibrosis. Carbohyd. Polym. 2022, 281, 118923. [Google Scholar] [CrossRef] [PubMed]
- El-Huneidi, W.; Anjum, S.; Mohammed, A.K.; Eshaq, S.B.; Abdrabh, S. Rosemarinic acid protects β-cell from STZ-induced cell damage via modulating NF-κβ pathway. Heliyon 2023, 9, e19234. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yao, Y.; Yang, J.; Zhou, B.; Zhu, Y.H. Inhibiting the SARM1-NAD+ axis reduces oxidative stress-induced damage to retinal and nerve cells. Int. Immunopharmacol. 2024, 134, 112193. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Sun, P.P.; Li, L.; Zou, Z.M.; Wu, Q.H. Heat stress suppresses MnSOD expression via p53-Sp1 interaction and induces oxidative stress damage in endothelial cells: Protective effects of MitoQ10 and Pifithrin-α. Heliyon 2023, 9, e22805. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.G.; Wei, T.; Gao, S.; Gao, N.; Chen, L.; Wu, Q.H. CD200R promotes high glucose-induced oxidative stress and damage in human retinal pigment epithelial cells by activating the mTOR signaling pathway. Tissue Cell 2024, 88, 102381. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhang, Y.; Mu, J.; Chen, J.; Zhang, C. Transplantation of human mesenchymal stem-cell-derived exosomes immobilized in an adhesive hydrogel for effective treatment of spinal cord injury. Nano Lett. 2020, 20, 4298–4305. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Xu, T.; Cheng, M.Y.; Fang, H.; Shen, X.J. Human umbilical cord mesenchymal stem cells regulate CD54 and CD105 in vascular endothelial cells and suppress inflammation in kawasaki disease. Exp. Cell Res. 2021, 409, 112941. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.Q.; Chang, Y.M.; Peng, C.Y.; Huang, S.; Li, G. Umbilical cord mesenchymal stem cells derived neurospheres promote long-term functional recovery but aggravate acute phase inflammation in experimental stroke. Neuroscience 2022, 480, 217–228. [Google Scholar] [CrossRef]
- Lee, Y.S.; Saha, S.K.; Lee, J.H.; Seo, K.-W.; Kang, K.-S. Human umbilical cord blood-derived mesenchymal stem cells ameliorate psoriasis-like skin inflammation in mice. Biochem. Biophys. Rep. 2017, 9, 281–288. [Google Scholar] [CrossRef]
- Yang, J.; Chen, Z.; Pan, D.; Li, H.; Shen, J. Umbilical cord-derived mesenchymal stem cell-derived exosomes combined pluronic F127 hydrogel promote chronic diabetic wound healing and complete skin regeneration. Int. J. Nanomed. 2020, 15, 5911–5926. [Google Scholar] [CrossRef] [PubMed]
- Kouhestani, M.; Ai, J.; Safari, M.; Hosseini, A.; Masoudifard, M. Study of anti-inflammatory and immunomodulatory potential of endometrial mesenchymal stem cells-derived exosomes with micro-ultrasound (μUS) guidance in regeneration β-islets on T1D animal model. J. Drug Deliv. Sci. Technol. 2023, 86, 104666. [Google Scholar] [CrossRef]
- Daga, D.R.; Marsaro, D.B.; Schaidt, B.; Fornazari, R.; Leite, L.M.B. The immunomodulatory effect of umbilical cord mesenchymal stem cells produced under xeno-free conditions. Cytotherapy 2021, 23, 35–36. [Google Scholar] [CrossRef]
- Zhou, C.; Yang, B.; Tian, Y.; Jiao, H.; Zheng, W. Immunomodulatory effect of human umbilical cord Wharton’s jelly-derived mesenchymal stem cells on lymphocytes. Cell Immunol. 2011, 272, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Xu, K.; Zhang, G.; Liu, Y.; Gao, J.F. Immunomodulatory effect of human umbilical cord mesenchymal stem cells on T lymphocytes in rheumatoid arthritis. Int. Immunopharmacol. 2019, 74, 105687. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Guo, Z.; Chen, K.; Yang, W.; Wan, X. Combined transplantation of mesenchymal stem cells and endothelial colony-forming cells accelerates refractory diabetic foot ulcer healing. Stem Cells Int. 2020, 2020, 8863649. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.; Ha, J.H.; Kim, I.; Bae, J.; Kim, S.W. The immunomodulatory effect of adipose-derived stem cells in xenograft transplantation model. Transplant. Proc. 2022, 54, 2388–2395. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.H.; Huang, X.; Ruan, G.P.; Pang, R.Q.; Chen, Q. Umbilical cord mesenchymal stem cells are able to undergo differentiation into functional islet-like cells in type 2 diabetic tree shrews. Mol. Cell. Probes 2017, 34, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Nie, Y.F.; Shang, J.M.; Liu, D.Q.; Meng, W.Q.; Ren, H.P. Apical papilla stem cell-derived exosomes regulate lipid metabolism and alleviate inflammation in the MCD-induced mouse NASH model. Biochem. Pharmacol. 2024, 222, 116073. [Google Scholar] [CrossRef]
- Lu, Z.L.; Li, Q.; Dai, Y.Y.; Pan, X.H. Association of co-exposure to polycyclic aromatic hydrocarbons and phthalates with oxidative stress and inflammation. Sci. Total Environ. 2024, 912, 169513. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, Y.; Shi, L.; Fang, L.; Xu, L. Neural stemness unifies cell tumorigenicity and pluripotent differentiation potential. J. Biol. Chem. 2022, 298, 102106. [Google Scholar] [CrossRef] [PubMed]
- Dadheecha, N.; Cuesta-Gomez, N.; Jasra, I.T.; Verhoeff, K.; Garza, B.M. Opportunities and impediments of human pluripotent stem cell-derived islets in the treatment of diabetes. J. Immunol. Regen. Med. 2022, 17, 100064. [Google Scholar] [CrossRef]
- Martín, P.G.; González, M.B.; Martínez, A.R.; Lara, V.G.; Naveros, B.C. Isolation and characterization of the environmental bacterial and fungi contamination in a pharmaceutical unit of mesenchymal stem cell for clinical use. Biologicals 2012, 40, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.; Han, F.; Xiong, N.; Wang, Y.; Feng, S. Standards of induced pluripotent stem cells derived clinical-grade neural stem cells preparation and quality control (2021 China version). J. Neurorestoratology 2021, 9, 13–30. [Google Scholar] [CrossRef]
- Wu, H.L.; Busman-Sahay, K.; Weber, W.C.; Waytashek, C.M.; Boyle, C.D. Allogeneic immunity clears latent virus following allogeneic stem cell transplantation in SIV-infected ART-suppressed macaques. Immunity 2023, 56, 1649–1663. [Google Scholar] [CrossRef] [PubMed]
- Spang, M.T.; Christman, K.L. Extracellular matrix hydrogel therapies: In vivo applications and development. Acta Biomater. 2018, 68, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Karimi, S.; Khorsandi, L.S.; Ai, J. Fabrication of bioartificial pancreas using decellularized rat testicular tissue. Acta Histochem. 2022, 124, 151928. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Xiao, Q.; Zhang, L.; Wang, X.; Huang, Q. CAPE-pNO2 attenuates diabetic cardiomyopathy through the NOX4/NF-κB pathway in STZ-induced diabetic mice. Biomed. Pharmacother. 2018, 108, 1640–1650. [Google Scholar] [CrossRef] [PubMed]
- Samaha, M.M.; Said, E.; Salem, H.A. Modulatory role of imatinib mesylate on pancreatic β-cells’ secretory functions in an STZ rat model of diabetes mellitus. Chem.-Biol. Interact. 2020, 328, 109197. [Google Scholar] [CrossRef]
- Adeleke, T.A.; Lawal, B.; Agwupuye, E.I.; Kuo, Y.; Eni, A.M. Apigetrin-enriched Pulmeria alba extract prevents assault of STZ on pancreatic β-cells and neuronal oxidative stress with concomitant attenuation of tissue damage and suppression of inflammation in the brain of diabetic rats. Biomed. Pharmacother. 2023, 162, 114582. [Google Scholar] [CrossRef]
- Amirshahrokhi, K.; Zohouri, A. Carvedilol prevents pancreatic β-cell damage and the development of type 1 diabetes in mice by the inhibition of proinflammatory cytokines, NF-κB, COX-2, iNOS and oxidative stress. Cytokine 2021, 138, 155394. [Google Scholar] [CrossRef]
- Hong, L.; Qin, W.; Shuzhen, G.; Juncheng, H.; Hanju, S. The protective effect of MT-α-glucan against streptozotocin (STZ)-induced NIT-1 pancreatic β-cell damage. Carbohyd. Polym. 2013, 92, 1211–1217. [Google Scholar] [CrossRef]
- Hu, C.M.; Li, J.S.; Cheah, K.P.; Lin, C.W.; Yu, W.Y. Effect of Sanguis draconis (a dragon’s blood resin) on streptozotocin- and cytokine-induced β-cell damage, in vitro and in vivo. Diabetes Res. Clin. Pract. 2011, 94, 417–425. [Google Scholar] [CrossRef]
- Li, H.; Tao, W.; Xu, X.; Chen, G.; Ma, W. Lycium barbarum polysaccharides alleviate pancreatic β-cells apoptosis through the inhibition of IFNγ pathway. J. Funct. Foods 2023, 107, 105706. [Google Scholar] [CrossRef]
- Ding, L.; Zhang, Y.; Wang, Y.; Wang, Y.; Tong, Z. Zhx2 maintains islet β-cell mass and function by transcriptionally regulating Pax6. iScience 2023, 26, 106871. [Google Scholar] [CrossRef]
- Lee, J.; Chung, J.O.; Park, S.Y.; Rajamohan, N.; Singh, A. Natural COA water inhibits mitochondrial ROS-mediated apoptosis through Plk3 downregulation under STZ diabetic stress in pancreatic β-cell lines. Biochem. Biophys. Rep. 2022, 30, 101247. [Google Scholar] [CrossRef]
- Bhatt, A.; Dhiman, N.; Giri, P.S.; Kasinathan, G.N.; Pati, F. Biocompatibility-on-a-chip: Characterization and evaluation of decellularized tendon extracellular matrix (tdECM) hydrogel for 3D stem cell culture in a microfluidic device. Int. J. Biol. Macromol. 2022, 213, 768–779. [Google Scholar] [CrossRef] [PubMed]
- Kalailingam, P.; Kannaian, B.; Tamilmani, E.; Kaliaperumal, R. Efficacy of natural diosgenin on cardiovascular risk, insulin secretion, and β-cells in streptozotocin (STZ)-induced diabetic rats. Phytomedicine 2014, 21, 1154–1161. [Google Scholar] [CrossRef]
- Espona-Noguera, A.; Ciriza, J.; Caño-Hernández, A.; Fernandez, L.; Ochoa, I. Tunable injectable alginate-based hydrogel for cell therapy in Type 1 Diabetes Mellitus. Int. J. Biol. Macromol. 2018, 107, 1261–1269. [Google Scholar] [CrossRef]
- Bal, T.; Nazli, C.; Okcu, A.; Duruksu, G.; Karaöz, E. Mesenchymal stem cells and ligand incorporation in biomimetic poly(ethylene glycol) hydrogels significantly improve insulin secretion from pancreatic islets. J. Tissue Eng. Regen. Med. 2014, 11, 694–703. [Google Scholar] [CrossRef]
- Dai, X.; Liu, Y.; Liu, T.; Zhang, Y.; Wang, S. SiJunZi decoction ameliorates bone quality and redox homeostasis and regulates advanced glycation end products/receptor for advanced glycation end products and WNT/β-catenin signaling pathways in diabetic mice. J. Ethnopharmacol. 2024, 319, 117167. [Google Scholar] [CrossRef] [PubMed]
- Lari, A.S.; Zahedi, P.; Ghourchian, H.; Khatibi, A. Microfluidic-based synthesized carboxymethyl chitosan nanoparticles containing metformin for diabetes therapy: In vitro and in vivo assessments. Carbohydr. Polym. 2021, 261, 117889. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.W.; Li, Y.; Jin, X.; Peng, Q.; Liu, X.F. Microwave-assisted synthesis of chitosan biguanidine hydrochloride and its antioxidant activity in vitro. J. Appl. Polym. Sci. 2016, 133, 43570. [Google Scholar] [CrossRef]
- Zhu, X.Q.; Shen, J.Y.; Feng, S.Y.; Huang, C.; Wang, H. Akkermansia muciniphila, which is enriched in the gut microbiota by metformin, improves cognitive function in aged mice by reducing the proinflammatory cytokine interleukin-6. Microbiome 2023, 11, 120. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, S.S.; Wang, L.; Liu, X.F.; Wu, Y.T. Chitooligosaccharide guanidine inhibits high glucose-induced activation of DAG/PKC pathway by regulating expression of GLUT2 in type 2 diabetic nephropathy rats. J. Funct. Foods 2018, 41, 41–47. [Google Scholar] [CrossRef]
- Zhang, S.S.; Zhang, H.; Wang, L.; Liu, X.F.; Wu, Y.T. Microwave-assisted synthesis of chitosan biguanidine hydrochloride and its regulation on InsR and GLUT2 in insulin resistant HepG2 cells. RSC Adv. 2017, 7, 10108–10117. [Google Scholar] [CrossRef]
- Gundala, N.K.V.; Naidu, V.G.M.; Das, U.N. Arachidonic acid and lipoxinA4 attenuate streptozotocin-induced cytotoxicity to RIN5 F cells in vitro and type 1 and type 2 diabetes mellitus in vivo. Nutrition 2017, 35, 61–80. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.K.; Mani, K.P. Endocan alters nitric oxide production in endothelial cells by targeting AKT/eNOS and NFkB/iNOS signaling. Nitric Oxide 2021, 117, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.Y.; Ko, Y.; Kim, M.J. Regulatory mechanisms of kaempferol on iNOS expression in RINm5F β-cells under exposure to interleukin-1β. Heliyon 2023, 9, e14818. [Google Scholar] [CrossRef] [PubMed]
- Xia, G.Q.; Zhu, M.P.; Li, J.W.; Huang, H. An alkaloid from Menispermum dauricum, dauricine mediates Ca2+ influx and inhibits NF-κB pathway to protect chondrocytes from IL-1β-induced inflammation and catabolism. J. Ethnopharmacol. 2024, 321, 117560. [Google Scholar] [CrossRef]
- Garg, M.; Johri, S.; Sagar, S.; Mundhada, A.; Agrawal, A.; Ray, P.; Chakraborty, K. Cardiolipin-mediated PPARγ S112 phosphorylation impairs IL-10 production and inflammation resolution during bacterial pneumonia. Cell Rep. 2021, 34, 108736. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Tang, J.; Li, C.; Liu, J.; Liu, H. Sulforaphane attenuates dextran sodium sulphate induced intestinal inflammation via IL-10/STAT3 signaling mediated macrophage phenotype switching. Food Sci. Hum. Wellness 2022, 11, 129–142. [Google Scholar] [CrossRef]
- Patel, R.; Parmar, N.; Rathwa, N.; Palit, S.P.; Li, Y. A novel therapeutic combination of sitagliptin and melatonin regenerates pancreatic β-cells in mouse and human islets. BBA Mol. Cell Res. 2022, 1869, 119263. [Google Scholar] [CrossRef] [PubMed]
- Babu, D.A.; Deering, T.G.; Mirmira, R.G. A feat of metabolic proportions: Pdx1 orchestrates islet development and function in the maintenance of glucose homeostasis. Mol. Genet. Metab. 2007, 92, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Zhang, T.; Lu, A.; Shiota, C.; Huard, M. Specific reprogramming of alpha cells to insulin-producing cells by short glucagon promoter-driven Pdx1 and MafA. Mol. Ther. Methods Clin. Dev. 2023, 28, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Yang, Z.; Ge, X.Y.; Gao, M.X.; Meng, R. Autonomous sensing of the insulin peptide by an olfactory G protein-coupled receptor modulates glucose metabolism. Cell Metab. 2022, 34, 240–255. [Google Scholar] [CrossRef] [PubMed]
- Schiano, E.; Novellino, E.; Fernández, M.M.G.; Lorinczova, H.T.; Tenore, G.C. Antioxidant and antidiabetic properties of a thinned-nectarine-based nanoformulation in a pancreatic β-cell line. Antioxidants 2023, 13, 63. [Google Scholar] [CrossRef]
- Neves, M.I.; Bidarra, S.J.; Magalhães, M.V.; Torres, A.L.; Moroni, L.; Barrias, C.C. Microstructured click hydrogels for cell contact guidance in 3D. Mater. Today Bio. 2023, 19, 100604. [Google Scholar] [CrossRef]
- Lee, K.; Chen, Y.; Li, X.; Kawazoe, N.; Yang, Y. Influence of viscosity on chondrogenic differentiation of mesenchymal stem cells during 3D culture in viscous gelatin solution-embedded hydrogels. J. Mater. Sci. Technol. 2021, 63, 1–8. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Liu, Y.; Wang, M.; Chen, S.; Miao, Q.; Liu, Z.; Zhang, B.; Deng, G. Repair Effect of Umbilical Cord Mesenchymal Stem Cells Embedded in Hydrogel on Mouse Insulinoma 6 Cells Injured by Streptozotocin. Polymers 2024, 16, 1845. https://doi.org/10.3390/polym16131845
Yang J, Liu Y, Wang M, Chen S, Miao Q, Liu Z, Zhang B, Deng G. Repair Effect of Umbilical Cord Mesenchymal Stem Cells Embedded in Hydrogel on Mouse Insulinoma 6 Cells Injured by Streptozotocin. Polymers. 2024; 16(13):1845. https://doi.org/10.3390/polym16131845
Chicago/Turabian StyleYang, Jia, Yang Liu, Minghui Wang, Shengqin Chen, Qingya Miao, Zhicong Liu, Bin Zhang, and Guodong Deng. 2024. "Repair Effect of Umbilical Cord Mesenchymal Stem Cells Embedded in Hydrogel on Mouse Insulinoma 6 Cells Injured by Streptozotocin" Polymers 16, no. 13: 1845. https://doi.org/10.3390/polym16131845
APA StyleYang, J., Liu, Y., Wang, M., Chen, S., Miao, Q., Liu, Z., Zhang, B., & Deng, G. (2024). Repair Effect of Umbilical Cord Mesenchymal Stem Cells Embedded in Hydrogel on Mouse Insulinoma 6 Cells Injured by Streptozotocin. Polymers, 16(13), 1845. https://doi.org/10.3390/polym16131845