
Recent Development in Biomedical Applications of Oligonucleotides with Triplex-Forming Ability




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
2.1. Oligonucleotides with Triplex-Forming Ability in Targeted Delivery
2.1.1. Substantial RNA–DNA Interactions
2.1.2. PNA
2.2. The Impact of TFOs on Gene Expression
Triplex-Mediated Modulation of Gene Transcription
2.3. Biosensor Based on Aptamers
2.4. G-Quadruplex Forming Aptamer
2.5. Nucleic Acid Motifs with pH Sensitivity
2.6. Triplex Cations
3. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hu, Y.; Cecconello, A.; Idili, A.; Ricci, F.; Willner, I. Triplex DNA nanostructures: From basic properties to applications. Angew. Chem. Int. Ed. 2017, 56, 15210–15233. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, A.R.; Rusling, D.A. Triplex-forming oligonucleotides: A third strand for DNA nanotechnology. Nucleic Acids Res. 2018, 46, 1021–1037. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Wang, G.; Vasquez, K.M. DNA triple helices: Biological consequences and therapeutic potential. Biochimie 2008, 90, 1117–1130. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, Y.; Sasaki, S. Enhancements in the utilization of antigene oligonucleotides in the nucleus by booster oligonucleotides. Org. Biomol. Chem. 2012, 10, 8336–8341. [Google Scholar] [CrossRef] [PubMed]
- Graham, M.K.; Brown, T.R.; Miller, P.S. Targeting the human androgen receptor gene with platinated triplex-forming oligonucleotides. Biochemistry 2015, 54, 2270–2282. [Google Scholar] [CrossRef]
- Goñi, J.R.; De La Cruz, X.; Orozco, M. Triplex-forming oligonucleotide target sequences in the human genome. Nucleic Acids Res. 2004, 32, 354–360. [Google Scholar] [CrossRef]
- Robles, J.; Grandas, A.; Pedroso, E.; Luque, F.; Eritja, R.; Orozco, M. Nucleic acid triple helices: Stability effects of nucleobase modifications. Curr. Org. Chem. 2002, 6, 1333–1368. [Google Scholar] [CrossRef]
- Soyfer, V.N.; Potaman, V.N. Triple-Helical Nucleic Acids; Springer: New York, NY, USA, 1996. [Google Scholar]
- Giovannangeli, C.; Helene, C. Triplex technology takes off. Nat. Biotechnol. 2000, 18, 1245–1246. [Google Scholar] [CrossRef]
- Ebbinghaus, S.W.; Vigneswaran, N.; Miller, C.R.; Chee-Awai, R.A.; Mayeld, C.A.; Curiel, D.T.; Miller, D.M. Efficient delivery of triplex forming oligonucleotides to tumor cells by adenovirus-polylysine complexes. Gene Ther. 1996, 3, 287–297. [Google Scholar]
- Zendegui, J.G.; Vasquez, K.M.; Tinsley, J.H.; Kessler, D.J.; Hogan, M.E. In vivo stability and kinetics of absorption and disposition of 3’phosphopropyl amine oligonucleotides. Nucleic Acids Res. 1992, 20, 307–314. [Google Scholar] [CrossRef]
- Bates, A.D.; Maxwell, A. DNA Topology; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- Fogg, J.M. Bullied no more: When and how DNA shoves proteins around. Q. Rev. Biophys. 2012, 45, 257–299. [Google Scholar] [CrossRef] [PubMed]
- Mirkin, S.M. DNA topology: Fundamentals. Encycl. Life Sci. 2001, 111. [Google Scholar] [CrossRef]
- Schvartzman, J.B.; Hernández, P.; Krimer, D.B.; Dorier, J.; Stasiak, A. Closing the DNA replication cycle: From simple circular molecules to supercoiled and knotted DNA catenanes. Nucleic Acids Res. 2019, 47, 7182–7198. [Google Scholar] [CrossRef] [PubMed]
- Sobetzko, P. Transcription-coupled DNA supercoiling dictates the chromosomal arrangement of bacterial genes. Nucleic Acids Res. 2016, 44, 1514–1524. [Google Scholar] [CrossRef] [PubMed]
- Newton, M.D. DNA stretching induces Cas9 off-target activity. Nat. Struct. Mol. Biol. 2019, 26, 185–192. [Google Scholar] [CrossRef]
- Travers, A.; Muskhelishvili, G. A common topology for bacterial and eukaryotic transcription initiation? EMBO Rep. 2007, 8, 147–151. [Google Scholar] [CrossRef]
- Van Aelst, K.; Martínez-Santiago, C.J.; Cross, S.J.; Szczelkun, M.D. The effect of DNA topology on observed rates of R-loop formation and DNA strand cleavage by CRISPR Cas12a. Genes 2019, 10, 169. [Google Scholar] [CrossRef]
- Holoch, D.; Moazed, D. RNA-mediated epigenetic regulation of gene expression. Nat. Rev. Genet. 2015, 16, 71–84. [Google Scholar] [CrossRef]
- Oo, J.A.; Brandes, R.P.; Leisegang, M.S. Long non-coding RNAs: Novel regulators of cellular physiology and function. Pflügers Arch.-Eur. J. Physiol. 2021, 474, 191–204. [Google Scholar] [CrossRef]
- Takemata, N.; Ohta, K. Role of non-coding RNA transcription around gene regulatory elements in transcription factor recruitment. RNA Biol. 2017, 14, 1–5. [Google Scholar] [CrossRef]
- Andric, V.; Nevers, A.; Hazra, D.; Auxilien, S.; Menant, A.; Graille, M.; Palancade, B.; Rougemaille, M. A scaffold lncRNA shapes the mitosis to meiosis switch. Nat. Commun. 2021, 12, 770. [Google Scholar] [CrossRef] [PubMed]
- Shimada, Y.; Mohn, F.; Bühler, M. The RNA-induced transcriptional silencing complex targets chromatin exclusively via interacting with nascent transcripts. Genes Dev. 2016, 30, 2571–2580. [Google Scholar] [CrossRef] [PubMed]
- Csankovszki, G.; Nagy, A.; Jaenisch, R. Synergism of Xist RNA, DNA methylation, and histone hypoacetylation in maintaining X chromosome inactivation. J. Cell Biol. 2001, 153, 773–784. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.Y.; Zheng, K.w.; Xiao, S. Mechanism, and manipulation of DNA: RNA hybrid G-quadruplex formation in the transcription of G-rich DNA. J. Am. Chem. Soc. 2014, 136, 1381–1390. [Google Scholar] [CrossRef]
- García Muse, T.; Aguilera, A. R loops: From physiological to pathological roles. Cell 2019, 179, 604–618. [Google Scholar] [CrossRef]
- Santos Pereira, J.M.; Aguilera, A. R loops: New modulators of genome dynamics and function. Nat. Rev. Genet. 2015, 16, 583–597. [Google Scholar] [CrossRef]
- Crossley, M.P.; Bocek, M.; Cimprich, K.A. R-loops as cellular regulators and genomic threats. Mol. Cell 2019, 73, 398–411. [Google Scholar] [CrossRef]
- Felsenfeld, G.; Davies, D.R.; Rich, A. Formation of a three-stranded polynucleotide molecule. J. Am. Chem. Soc. 1957, 79, 2023–2024. [Google Scholar] [CrossRef]
- Escudeé, C.; Francçois, J.C.; Sun, J.-S. Stability of triple helices containing RNA and DNA strands: Experimental and molecular modeling studies. Nucleic Acids Res. 1993, 21, 5547–5553. [Google Scholar] [CrossRef]
- Mondal, T.; Subhash, S.; Vaid, R.; Enroth, S.; Uday, S.; Reinius, B.; Mitra, S.; Mohammed, A.; James, A.R.; Hoberg, E.; et al. MEG3 long noncoding RNA regulates the TGF-β pathway genes through formation of RNA–DNA triplex structures. Nat. Commun. 2015, 6, 7743. [Google Scholar] [CrossRef]
- Chu, C.; Qu, K.; Zhong, F.L.; Artandi, S.E.; Chang, H.Y. Genomic maps of long noncoding RNA occupancy reveal principles of RNA-chromatin interactions. Mol. Cell 2011, 44, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Vrba, L.; Futscher, B.W. Epigenetic silencing of lncRNA MORT in 16 TCGA cancer types. F1000Research 2018, 7, 211. [Google Scholar] [CrossRef]
- Jiang, M.C.; Ni, J.J.; Cui, W.Y. Emerging roles of lncRNA in cancer and therapeutic opportunities. Am. J. Cancer Res. 2019, 9, 1354. [Google Scholar]
- Zhou, Z.; Giles, K.E.; Felsenfeld, G. DNA· RNA triple helix formation can function as a cis-acting regulatory mechanism at the human β-globin locus. Proc. Natl. Acad. Sci. USA 2019, 116, 6130–6139. [Google Scholar] [CrossRef] [PubMed]
- Garratt, H.; Ashburn, R.; Sopic, M. Long Non-Coding RNA Regulation of Epigenetics in Vascular Cells. Non-Coding RNA 2021, 7, 62. [Google Scholar] [CrossRef]
- Maldonado, R.; Filarsky, M.; Grummt, I. Purine–and pyrimidine–triple-helix-forming oligonucleotides recognize qualitatively different target sites at the ribosomal DNA locus. RNA 2018, 24, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Buske, F.A.; Bauer, D.C.; Mattick, J.S.; Bailey, T.L. Triplexator: Detecting nucleic acid triple helices in genomic and transcriptomic data. Genome Res. 2012, 22, 1372–1381. [Google Scholar] [CrossRef]
- Kuo, C.-C.; Hänzelmann, S.; Cetin, N.S.; Frank, S.; Zajzon, B.; Derks, J.-P.; Akhade, V.S.; Ahuja, G.; Kanduri, C.; Grummt, I.; et al. Detection of RNA–DNA binding sites in long noncoding RNAs. Nucleic Acids Res. 2019, 47, e32. [Google Scholar] [CrossRef]
- He, S.; Zhang, H.; Liu, H.; Zhu, H. LongTarget: A tool to predict lncRNA DNA-binding motifs and binding sites via Hoogsteen base-pairing analysis. Bioinformatics 2015, 31, 178–186. [Google Scholar] [CrossRef]
- Antonov, I.V.; Mazurov, E.; Borodovsky, M. Prediction of lncRNAs and their interactions with nucleic acids: Benchmarking bioinformatics tools. Brief. Bioinform. 2019, 20, 551–564. [Google Scholar] [CrossRef]
- Zhang, Y.; Long, Y.; Kwoh, C.K. Deep learning-based DNA: RNA triplex forming potential prediction. Brief. Bioinform. 2020, 21, 522. [Google Scholar] [CrossRef] [PubMed]
- Sentürk Cetin, N.; Kuo, C.C.; Ribarska, T.; Li, R.; Costa, I.G.; Grummt, I. Isolation, and genome-wide characterization of cellular DNA: RNA triplex structures. Nucleic Acids Res. 2019, 47, 2306–2321. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Li, X.; Luo, D. GRID-seq for comprehensive analysis of global RNA–chromatin interactions. Nat. Protoc. 2019, 14, 2036–2068. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.; Sridhar, B.; Rivas Astroza, M. Mapping RNA-chromatin interactions. FASEB J. 2018, 32, 525.2. [Google Scholar] [CrossRef]
- Wu, W.; Yan, Z.; Nguyen, T.C. Mapping RNA–chromatin interactions by sequencing with iMARGI. Nat. Protoc. 2019, 14, 3243–3272. [Google Scholar] [CrossRef]
- Bell, J.C.; Jukam, D.; Teran, N.A. Chromatin-associated RNA sequencing (ChAR-seq) maps genome-wide RNA-to-DNA contacts. Elife 2018, 7, e27024. [Google Scholar] [CrossRef]
- Bonetti, A.; Agostini, F.; Suzuki, A.M. RADICL-seq identifies general and cell type–specific principles of genome-wide RNA-chromatin interactions. Nat. Commun. 2020, 11, 1018. [Google Scholar] [CrossRef]
- Gavrilov, A.A.; Zharikova, A.A.; Galitsyna, A.A. Studying RNA–DNA interactome by Red-C identifies noncoding RNAs associated with various chromatin types and reveals transcription dynamics. Nucleic Acids Res. 2020, 48, 6699–6714. [Google Scholar] [CrossRef]
- Schmitz, K.M.; Mayer, C.; Postepska, A.; Grummt, I. Interaction of noncoding RNA with the rDNA promoter mediates recruitment of DNMT3b and silencing of rRNA genes. Genes Dev. 2010, 24, 2264–2269. [Google Scholar] [CrossRef]
- Grote, P.; Herrmann, B.G. The long non-coding RNA Fendrr links epigenetic control mechanisms to gene regulatory networks in mammalian embryogenesis. RNA Biol. 2013, 10, 1579–1585. [Google Scholar] [CrossRef]
- O’Leary, V.B.; Ovsepian, S.V.; Carrascosa, L.G.; Buske, F.A.; Radulovic, V.; Niyazi, M.; Moertl, S.; Trau, M.; Atkinson, M.J.; Anastasov, N. PARTICLE, a triplex-forming long ncRNA, regulates locus-specific methylation in response to low-dose irradiation. Cell Rep. 2015, 11, 474–485. [Google Scholar] [CrossRef] [PubMed]
- Postepska Igielska, A.; Giwojna, A.; Gasri-Plotnitsky, L.; Schmitt, N.; Dold, A.; Ginsberg, D.; Grummt, I. LncRNA Khps1 regulates expression of the proto-oncogene SPHK1 via triplex-mediated changes in chromatin structure. Mol. Cell 2015, 60, 626–636. [Google Scholar] [CrossRef] [PubMed]
- Kalwa, M.; Hänzelmann, S.; Otto, S.; Kuo, C.C.; Franzen, J.; Joussen, S.; Fernandez-Rebollo, E.; Rath, B.; Koch, C.; Hofmann, A.; et al. The lncRNA HOTAIR impacts on mesenchymal stem cells via triple helix formation. Nucleic Acids Res. 2016, 44, 10631–10643. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Ke, H.; Zhang, H.; Ma, Y.; Ao, L.; Zou, L.; Yang, Q.; Zhu, H.; Nie, J.; Wu, C.; et al. LncRNA MIR100HG promotes cell proliferation in triple-negative breast cancer through triplex formation with p27 loci. Cell Death Dis. 2018, 9, 805. [Google Scholar] [CrossRef]
- Zhao, Z.; Sentürk, N.; Song, C.; Grummt, I. ncRNA PAPAS tethered to the rDNA enhancer recruits hypophosphorylated CHD4/NuRD to repress rRNA synthesis at elevated temperatures. Genes Dev. 2018, 32, 836–848. [Google Scholar] [CrossRef]
- Simon, M.D.; Wang, C.I.; Kharchenko, P.V.; West, J.A.; Chapman, B.A.; Oleksiak, A.A.; Borowsky, M.L.; Kuroda, M.I.; Kingston, R.E. The genomic binding sites of a noncoding RNA. Proc. Natl. Acad. Sci. USA 2011, 108, 20497–20502. [Google Scholar] [CrossRef]
- Engreitz, J.M.; Pandya-Jones, A.; McDonel, P.; Shishkin, A.; Sirokman, K.; Surka, C.; Kadri, S.; Xing, J.; Goren, A.; Lander, E.S.; et al. The Xist lncRNA exploits three-dimensional genome architecture to spread across the X chromosome. Science 2013, 341, 1237973. [Google Scholar] [CrossRef]
- Buske, F.A.; Bauer, D.C.; Mattick, J.S.; Bailey, T.L. Triplex-Inspector: An analysis tool for triplex-mediated targeting of genomic loci. Bioinformatics 2013, 29, 1895–1897. [Google Scholar] [CrossRef]
- Houlard, M.; Artus, J.; Leguillier, T.; Vandormael-Pournin, S.; Cohen-Tannoudji, M. DNA-RNA hybrids contribute to the replication dependent genomic instability induced by Omcg1 deficiency. Cell Cycle 2011, 10, 108–117. [Google Scholar] [CrossRef]
- Balk, B.; Dees, M.; Bender, K.; Luke, B. The differential processing of telomeres in response to increased telomeric transcription and RNA–DNA hybrid accumulation. RNA Biol. 2014, 11, 95–100. [Google Scholar] [CrossRef]
- Stuckey, R.; García-Rodríguez, N.; Aguilera, A.; Wellinger, R.E. Role for RNA: DNA hybrids in origin-independent replication priming in a eukaryotic system. Proc. Natl. Acad. Sci. USA 2015, 112, 5779–5784. [Google Scholar] [CrossRef] [PubMed]
- Balk, B.; Maicher, A.; Dees, M.; Klermund, J.; Luke-Glaser, S.; Bender, K.; Luke, B. Telomeric RNA-DNA hybrids affect telomere-length dynamics and senescence. Nat. Struct. Mol. Biol. 2013, 20, 1199–1205. [Google Scholar] [CrossRef] [PubMed]
- Li, T.K.; Barbieri, C.M.; Lin, H.-C.; Rabson, A.B.; Yang, G.; Fan, Y.; Gaffney, B.L.; Jones, R.A.; Pilch, D.S. Drug Targeting of HIV-1 RNA DNA Hybrid Structures: Thermodynamics of Recognition and Impact on Reverse Transcriptase-Mediated Ribonuclease H Activity and Viral Replication. Biochemistry 2004, 43, 9732–9742. [Google Scholar] [CrossRef] [PubMed]
- Lombraña, R.; Almeida, R.; Alvarez, A.; Gómez, M. R-loops, and initiation of DNA replication in human cells: A missing link? Front. Genet. 2015, 6, 158. [Google Scholar] [CrossRef] [PubMed]
- Saretzki, G. Telomerase inhibition as cancer therapy. Cancer Lett. 2003, 194, 209–219. [Google Scholar] [CrossRef]
- Mergny, J.L.; Lacroix, L.; Teulade-Fichou, M.P.; Hounsou, C.; Guittat, L.; Hoarau, M.; Arimondo, P.B.; Vigneron, J.-P.; Lehn, J.-M.; Riou, J.-F. Telomerase inhibitors based on quadruplex ligands selected by a fluorescence assay. Proc. Natl. Acad. Sci. USA 2001, 98, 3062–3067. [Google Scholar] [CrossRef]
- Su, H.P.; Yan, Y.; Prasad, G.S.; Smith, R.F.; Daniels, C.L.; Abeywickrema, P.D.; Reid, J.C.; Loughran, H.M.; Kornienko, M.; Sharma, S. Structural basis for the inhibition of RNase H activity of HIV-1 reverse transcriptase by RNase H active site-directed inhibitors. J. Virol. 2010, 84, 7625–7633. [Google Scholar] [CrossRef] [PubMed]
- Tramontano, E.; Di Santo, R. HIV-1 RT-associated RNase H function inhibitors: Recent advances in drug development. Curr. Med. Chem. 2010, 17, 2837–2853. [Google Scholar] [CrossRef]
- Ren, J.; Qu, X.; Dattagupta, N.; Chaires, J. Molecular recognition of a RNA: DNA hybrid structure. J. Am. Chem. Soc. 2001, 123, 6742–6743. [Google Scholar] [CrossRef]
- Shaw, N.N.; Arya, D.P. Recognition of the unique structure of DNA: RNA hybrids. Biochimie 2008, 90, 1026–1039. [Google Scholar] [CrossRef]
- West, C.; Francis, R.; Friedman, S.H. Small molecule/nucleic acid affinity chromatography: Application for the identification of telomerase inhibitors which target its key RNA/DNA heteroduplex. Bioorganic Med. Chem. Lett. 2001, 11, 2727–2730. [Google Scholar] [CrossRef] [PubMed]
- Wheelhouse, R.T.; Chaires, J.B. Drug Binding to DNA⋅RNA Hybrid Structures. In Drug-DNA Interaction Protocols; Humana Press: Totowa, NJ, USA, 2010; pp. 55–70. [Google Scholar]
- Shaw, N.N.; Xi, H.; Arya, D.P. Molecular recognition of a DNA: RNA hybrid: Sub-nanomolar binding by a neomycin–methidium conjugate. Bioorganic Med. Chem. Lett. 2008, 18, 4142–4145. [Google Scholar] [CrossRef]
- Xi, H.; Davis, E.; Ranjan, N.; Xue, L.; Hyde-Volpe, D.; Arya, D.P. Thermodynamics of nucleic acid “shape readout” by an aminosugar. Biochemistry 2011, 50, 9088–9113. [Google Scholar] [CrossRef] [PubMed]
- Wheelhouse, R.T.; Garbett, N.C.; Buurma, N.J.; Chaires, J. Probing the Molecular Recognition of a DNA⋅ RNA Hybrid Duplex. Angew. Chem. 2010, 122, 3275–3278. [Google Scholar] [CrossRef]
- Zeraati, M.; Langley, D.B.; Schofield, P.; Moye, A.L.; Rouet, R.; Hughes, W.E.; Bryan, T.M.; Dinger, M.E.; Christ, D. I-motif DNA structures are formed in the nuclei of human cells. Nat. Chem. 2018, 10, 631–637. [Google Scholar] [CrossRef]
- Brogden, A.L.; Hopcroft, N.H.; Searcey, M.; Cardin, C.J. Ligand Bridging of the DNA Holliday Junction: Molecular Recognition of a Stacked-X Four-Way Junction by a Small Molecule. Angew. Chem. Int. Ed. 2007, 46, 3850–3854. [Google Scholar] [CrossRef]
- Fox, K.R.; Brown, T.; Rusling, D.A. DNA-targeting Molecules as Therapeutic Agents; The Royal Society of Chemistry: London, UK, 2018; pp. 1–32. [Google Scholar]
- Hoogsteen, K. The structure of crystals containing a hydrogen-bonded complex of 1-methylthymine and 9-methyladenine. Acta Crystallogr. 1959, 12, 822–823. [Google Scholar] [CrossRef]
- Carrascosa, L.G.; Gomez-Montes, S.; Avino, A.; Nadal, A.; Pla, M.; Eritja, R.; Lechuga, L.M. Sensitive and label-free biosensing of RNA with predicted secondary structures by a triplex affinity capture method. Nucleic Acids Res. 2012, 40, e56. [Google Scholar] [CrossRef]
- Wang, G.; Seidman, M.M.; Glazer, P.M. Mutagenesis in mammalian cells induced by triple helix formation and transcription-coupled repair. Science 1996, 271, 802–805. [Google Scholar] [CrossRef] [PubMed]
- Okamura, H.; Taniguchi, Y.; Sasaki, S. Aminopyridinyl–pseudodeoxycytidine derivatives selectively stabilize antiparallel triplex DNA with multiple CG inversion sites. Angew. Chem. Int. Ed. 2016, 55, 12445–12449. [Google Scholar] [CrossRef]
- Sun, J.S.; François, J.C.; Montenay-Garestier, T.; Saison-Behmoaras, T.; Roig, V.; Thuong, N.T.; Hélène, C. Sequence-specific intercalating agents: Intercalation at specific sequences on duplex DNA via major groove recognition by oligonucleotide-intercalator conjugates. Proc. Natl. Acad. Sci. USA 1989, 86, 9198–9202. [Google Scholar] [CrossRef] [PubMed]
- Paramasivam, M.; Cogoi, S.; Filichev, V.V.; Bomholt, N.; Pedersen, E.B.; Xodo, L.E. Purine twisted-intercalating nucleic acids: A new class of anti-gene molecules resistant to potassium-induced aggregation. Nucleic Acids Res. 2008, 36, 3494–3507. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, E.B.; Osman, A.M.A.; Globisch, D.; Paramasivam, M.; Cogoi, S.; Bomholt, N.; Jorgensen, P.T.; Xodo, L.E.; Filichev, V.V. Triplex glue by synthesizing conjugated flexible intercalators. In Nucleic Acids Symposium Series; Oxford University Press: Oxford, UK, 2008; pp. 37–38. [Google Scholar]
- Klimkowski, P.; De Ornellas, S.; Singleton, D.; El-Sagheer, A.H.; Brown, T. Design of thiazole orange oligonucleotide probes for detection of DNA and RNA by fluorescence and duplex melting. Org. Biomol. Chem. 2019, 17, 5943–5950. [Google Scholar] [CrossRef]
- Sigman, D.S.; Bruice, T.W.; Mazumder, A.; Sutton, C.L. Targeted chemical nucleases. Acc. Chem. Res. 1993, 26, 98–104. [Google Scholar] [CrossRef]
- Pitié, M.; Sudres, B.; Meunier, B. Dramatic increase of the DNA cleavage activity of Cu(Clip-phen) by fixing the bridging linker on the C3 position of the phenanthroline units. Chem. Commun. 1998, 2597–2598. [Google Scholar] [CrossRef]
- Bales, B.C.; Kodama, T.; Weledji, Y.N.; Pitie, M.; Meunier, B.; Greenberg, M.M. Mechanistic studies on DNA damage by minor groove binding copper–phenanthroline conjugates. Nucleic Acids Res. 2005, 33, 5371–5379. [Google Scholar] [CrossRef]
- Panattoni, A.; El-Sagheer, A.H.; Brown, T.; Kellett, A.; Hocek, M. Oxidative DNA Cleavage with Clip-Phenanthroline Triplex-Forming Oligonucleotide Hybrids. ChemBioChem 2020, 21, 991–1000. [Google Scholar] [CrossRef]
- Gasser, G.; Pinto, A.; Neumann, S.; Sosniak, A.M.; Seitz, M.; Merz, K.; Heumann, R.; Metzler-Nolte, N. Synthesis, characterisation and bioimaging of a fluorescent rhenium-containing PNA bioconjugate. Dalton Trans. 2012, 41, 2304–2313. [Google Scholar] [CrossRef]
- Slator, C.; Molphy, Z.; McKee, V.; Long, C.; Brown, T.; Kellett, A. Di-copper metallodrugs promote NCI-60 chemotherapy via singlet oxygen and superoxide production with tandem TA/TA and AT/AT oligonucleotide discrimination. Nucleic Acids Res. 2018, 46, 2733–2750. [Google Scholar] [CrossRef]
- Kellett, A.; Molphy, Z.; Slator, C.; Mckee, V.; Farrell, N.P. Molecular methods for assessment of non-covalent metallodrug–DNA interactions. Chem. Soc. Rev. 2019, 48, 971–988. [Google Scholar] [CrossRef]
- Levine, M.; Wang, Y.; Padayatty, S.J.; Morrow, J. A new recommended dietary allowance of vitamin C for healthy young women. Proc. Natl. Acad. Sci. USA 2001, 98, 9842–9846. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.N.; Miller, S.C.; Varani, G.; Calabrese, J.M.; Magnuson, T. Multimodal long noncoding RNA interaction networks: Control panels for cell fate specification. Genetics 2019, 213, 1093–1110. [Google Scholar] [CrossRef] [PubMed]
- Yao, R.W.; Wang, Y.; Chen, L.L. Cellular functions of long noncoding RNAs. Nat. Cell Biol. 2019, 21, 542–551. [Google Scholar] [CrossRef]
- Treiber, T.; Treiber, N.; Meister, G. Regulation of microRNA biogenesis and its crosstalk with other cellular pathways. Nat. Rev. Mol. Cell Biol. 2019, 20, 5–20. [Google Scholar] [CrossRef] [PubMed]
- Mercer, T.R.; Mattick, J.S. Structure and function of long noncoding RNAs in epigenetic regulation. Nat. Struct. Mol. Biol. 2013, 20, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Novikova, I.V.; Hennelly, S.P.; Tung, C.S.; Sanbonmatsu, K.Y. Rise of the RNA machines: Exploring the structure of long non-coding RNAs. J. Mol. Biol. 2013, 425, 3731–3746. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.E.; Bentin, T.; Nielsen, P.E. High-affinity triplex targeting of double stranded DNA using chemically modified peptide nucleic acid oligomers. Nucleic Acids Res. 2009, 37, 4498–4507. [Google Scholar] [CrossRef]
- Wittung, P.; Nielsen, P.; Norden, B. Extended DNA-Recognition Repertoire of Peptide Nucleic Acid (PNA): PNA–dsDNA Triplex Formed with Cytosine-Rich Homopyrimidine PNA. Biochemistry 1997, 36, 7973–7979. [Google Scholar] [CrossRef]
- Li, M.; Zengeya, T.; Rozners, E. Short peptide nucleic acids bind strongly to homopurine tract of double helical RNA at pH 5. 5. J. Am. Chem. Soc. 2010, 132, 8676–8681. [Google Scholar] [CrossRef]
- Zengeya, T.; Gupta, P.; Rozners, E. Triple-helical recognition of RNA using 2-aminopyridine-modified PNA at physiologically relevant conditions. Angew. Chem. 2012, 124, 12761–12764. [Google Scholar] [CrossRef]
- Muse, O.; Zengeya, T.; Mwaura, J.; Hnedzko, D.; McGee, D.W.; Grewer, C.T.; Rozners, E. Sequence selective recognition of double-stranded RNA at physiologically relevant conditions using PNA-peptide conjugates. ACS Chem. Biol. 2013, 8, 1683–1686. [Google Scholar] [CrossRef] [PubMed]
- Hnedzko, D.; McGee, D.W.; Karamitas, Y.A.; Rozners, E. Sequence-selective recognition of double-stranded RNA and enhanced cellular uptake of cationic nucleobase and backbone-modified peptide nucleic acids. RNA 2017, 23, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, P.E.; Egholm, M.; Berg, R.H.; Buchardt, O. Sequence-selective recognition of DNA by strand displacement with a thymine-substituted polyamide. Science 1991, 254, 1497–1500. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Kierzek, E.; Loo, Z.P.; Antonio, M.; Yau, Y.H.; Chuah, Y.W.; Geifman-Shochat, S.; Kierzek, R.; Chen, G. Recognition of RNA duplexes by chemically modified triplex-forming oligonucleotides. Nucleic Acids Res. 2013, 41, 6664–6673. [Google Scholar] [CrossRef]
- Devi, G.; Yuan, Z.; Lu, Y.; Zhao, Y.; Chen, G. Incorporation of thio-pseudoisocytosine into triplex-forming peptide nucleic acids for enhanced recognition of RNA duplexes. Nucleic Acids Res. 2014, 42, 4008–4018. [Google Scholar] [CrossRef]
- Sato, T.; Sakamoto, N.; Nishizawa, S. Kinetic and thermodynamic analysis of triplex formation between peptide nucleic acid and double-stranded RNA. Org. Biomol. Chem. 2018, 16, 1178–1187. [Google Scholar] [CrossRef]
- Taehtinen, V.; Granqvist, L.; Murtola, M.; Strömberg, R.; Virta, P. 19F NMR Spectroscopic Analysis of the Binding Modes in Triple-Helical Peptide Nucleic Acid (PNA)/MicroRNA Complexes. Chem.–A Eur. J. 2017, 23, 7113–7124. [Google Scholar] [CrossRef]
- Kim, K.T.; Chang, D.; Winssinger, N. Double-Stranded RNA-Specific Templated Reaction with Triplex Forming PNA. Helv. Chim. Acta 2018, 101, e1700295. [Google Scholar] [CrossRef]
- Hu, J.; Matsui, M.; Gagnon, K.T.; Schwartz, J.C.; Gabillet, S.; Arar, K.; Wu, J.; Bezprozvanny, I.; Corey, D.R. Allele-specific silencing of mutant huntingtin and ataxin-3 genes by targeting expanded CAG repeats in mRNAs. Nat. Biotechnol. 2009, 27, 478–484. [Google Scholar] [CrossRef]
- Hu, J.; Corey, D.R. Inhibiting gene expression with peptide nucleic acid (PNA)—Peptide conjugates that target chromosomal DNA. Biochemistry 2007, 46, 7581–7589. [Google Scholar] [CrossRef]
- Fabani, M.M.; Gait, M.J. MIR-122 targeting with LNA/2′-O-methyl oligonucleotide mixmers, peptide nucleic acids (PNA), and PNA–peptide conjugates. RNA 2008, 14, 336–346. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.J.; Ivanova, G.D.; Verbeure, B.; Williams, D.; Arzumanov, A.A.; Abes, S.; Lebleu, B.; Gait, M.J. Cell-penetrating peptide conjugates of peptide nucleic acids (PNA) as inhibitors of HIV-1 Tat-dependent trans-activation in cells. Nucleic Acids Res. 2005, 33, 6837–6849. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Zengeya, T.; Rozners, E. Triple helical recognition of pyrimidine inversions in polypurine tracts of RNA by nucleobase modified PNA. Chem. Commun. 2011, 47, 11125–11127. [Google Scholar] [CrossRef] [PubMed]
- Hari, Y.; Obika, S.; Imanishi, T. Towards the sequence-selective recognition of double-stranded DNA containing pyrimidine-purine interruptions by triplex-forming oligonucleotides. Eur. J. Org. Chem. 2012, 2012, 2875–2887. [Google Scholar] [CrossRef]
- Prevot Halter, I.; Leumann, C.J. Selective recognition of a CG base-pair in the parallel DNA triple-helical binding motif. Bioorganic Med. Chem. Lett. 1999, 9, 2657–2660. [Google Scholar] [CrossRef] [PubMed]
- Endoh, T.; Hnedzko, D.; Rozners, E.; Sugimoto, N. Nucleobase-Modified PNA Suppresses Translation by Forming a Triple Helix with a Hairpin Structure in mRNA In Vitro and in Cells. Angew. Chem. Int. Ed. 2016, 55, 899–903. [Google Scholar] [CrossRef]
- Ranasinghe, R.T.; Rusling, D.A.; Powers, V.E.C.; Fox, K.R.; Brown, T. Recognition of CG inversions in DNA triple helices by methylated 3H-pyrrolo [2,3-d] pyrimidin-2(7H)-one nucleoside analogues. Chem. Commun. 2005, 2555–2557. [Google Scholar] [CrossRef]
- Anstaett, P.; Zheng, Y.; Thai, T.; Funston, A.M.; Bach, U.; Gasser, G. Synthesis of Stable Peptide Nucleic Acid-Modified Gold Nanoparticles, and their Assembly onto Gold Surfaces. Angew. Chem. 2013, 125, 4311–4314. [Google Scholar] [CrossRef]
- Simon, L.; Lautner, G.; Gyurcsányi, R.E. Reliable microspotting methodology for peptide-nucleic acid layers with high hybridization efficiency on gold SPR imaging chips. Anal. Methods 2015, 7, 6077–6082. [Google Scholar] [CrossRef]
- Cadoni, E.; Rosa Gastaldo, D.; Manicardi, A.; Mancin, F.; Madder, A. Exploiting Double Exchange Diels-Alder Cycloadditions for Immobilization of Peptide Nucleic Acids on Gold Nanoparticles. Front. Chem. 2020, 8, 4. [Google Scholar] [CrossRef]
- Fabbri, E.; Manicardi, A.; Tedeschi, T.; Sforza, S.; Bianchi, N.; Brognara, E.; Finotti, A.; Breveglieri, G.; Borgatti, M.; Corradini, R. Modulation of the biological activity of microRNA-210 with peptide nucleic acids (PNAs). ChemMedChem 2011, 6, 2192–2202. [Google Scholar] [CrossRef] [PubMed]
- Brognara, E.; Fabbri, E.; Aimi, F.; Manicardi, A.; Bianchi, N.; Finotti, A.; Breveglieri, G.; Borgatti, M.; Corradini, R.; Marchelli, R. Peptide nucleic acids targeting miR-221 modulate p27Kip1 expression in breast cancer MDA-MB-231 cells. Int. J. Oncol. 2012, 41, 2119–2127. [Google Scholar] [CrossRef] [PubMed]
- Brandén, L.J.; Mohamed, A.J.; Smith, C.I. A peptide nucleic acid–nuclear localization signal fusion that mediates nuclear transport of DNA. Nat. Biotechnol. 1999, 17, 784–787. [Google Scholar] [CrossRef]
- Gambari, R. Peptide nucleic acids: A review on recent patents and technology transfer. Expert Opin. Ther. Pat. 2014, 24, 267–294. [Google Scholar] [CrossRef] [PubMed]
- Bayramoglu, G.; Ozalp, V.C.; Yilmaz, M.; Guler, U.; Salih, B.; Arica, M.Y. Lysozyme specific aptamer immobilized MCM-41 silicate for single-step purification and quartz crystal microbalance (QCM)-based determination of lysozyme from chicken egg white. Microporous Mesoporous Mater. 2015, 207, 95–104. [Google Scholar] [CrossRef]
- Yamaoki, Y.; Kiyoishi, A.; Miyake, M.; Kano, F.; Murata, M.; Nagata, T.; Katahira, M. The first successful observation of in-cell NMR signals of DNA and RNA in living human cells. Phys. Chem. Chem. Phys. 2018, 20, 2982–2985. [Google Scholar] [CrossRef]
- Dzatko, S.; Krafcikova, M.; Haensel-Hertsch, R.; Fessl, T.; Fiala, R.; Loja, T.; Krafcik, D.; Mergny, J.-L.; Foldynova-Trantirkova, S.; Trantirek, L. Evaluation of the stability of DNA i-Motifs in the nuclei of living mammalian cells. Angew. Chem. Int. Ed. 2018, 57, 2165–2169. [Google Scholar] [CrossRef]
- Bao, H.L.; Liu, H.S.; Xu, Y. Downregulated miRNA-26a-5p induces the apoptosis of endothelial cells in coronary heart disease by inhibiting PI3K/AKT pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 4940–4947. [Google Scholar]
- Bao, H.L.; Masuzawa, T.; Oyoshi, T.; Xu, Y. Oligonucleotides DNA containing 8-trifluoromethyl-2′-deoxyguanosine for observing Z-DNA structure. Nucleic Acids Res. 2020, 48, 7041–7051. [Google Scholar] [CrossRef]
- Yamaoki, Y.; Nagata, T.; Sakamoto, T.; Katahira, M. Recent progress of in-cell NMR of nucleic acids in living human cells. Biophys. Rev. 2020, 12, 411–417. [Google Scholar] [CrossRef]
- Yamaoki, Y.; Nagata, T.; Sakamoto, T.; Katahira, M. Observation of nucleic acids inside living human cells by in-cell NMR spectroscop. Biophys. Phys. 2020, 17, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Barraud, P.; Gato, A.; Heiss, M.; Catala, M.; Kellner, S.; Tisne, C. Time-resolved NMR monitoring of tRNA maturation. Nat. Commun. 2019, 10, 3373. [Google Scholar] [CrossRef] [PubMed]
- Moser, H.E.; Dervan, P.B. Sequence-specific cleavage of double helical DNA by triple helix formation. Science 1987, 238, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Catapano, C.V.; McGuffie, E.M.; Pacheco, D.; Carbone, G.M. Inhibition of gene expression and cell proliferation by triple helix-forming oligonucleotides directed to the c-myc gene. Biochemistry 2000, 39, 5126–5138. [Google Scholar] [CrossRef]
- Mairal, T.; Cengiz Özalp, V.; Lozano Sánchez, P.; Mir, M.; Katakis, I.; O’Sullivan, C.K. Aptamers: Molecular tools for analytical applications. Anal. Bioanal. Chem. 2008, 390, 989–1007. [Google Scholar] [CrossRef]
- Tombelli, S.; Minunni, M.; Mascini, M. Analytical applications of aptamers. Biosens. Bioelectron. 2005, 20, 2424–2434. [Google Scholar] [CrossRef]
- Sharma, R.; Ragavan, K.V.; Thakur, M.S.; Raghavarao, K.S. Recent advances in nanoparticle based aptasensors for food contaminants. Biosens. Bioelectron. 2015, 74, 612–627. [Google Scholar] [CrossRef]
- Kim, Y.S.; Raston, N.H.A.; Gu, M.B. Aptamer-based nanobiosensors. Biosens. Bioelectron. 2016, 76, 2–19. [Google Scholar]
- Mazaafrianto, D.N.; Maeki, M.; Ishida, A.; Tani, H.; Tokeshi, M. Recent microdevice-based aptamer sensors. Micromachines 2018, 9, 202. [Google Scholar] [CrossRef]
- Razmi, N.; Baradaran, B.; Hejazi, M.; Hasanzadeh, M.; Mosafer, J.; Mokhtarzadeh, A.; de la Guardia, M. Recent advances on aptamer-based biosensors to detection of platelet-derived growth factor. Biosens. Bioelectron. 2018, 113, 58–71. [Google Scholar]
- Kaur, N.; Bharti, A.; Batra, S.; Rana, S.; Rana, S.; Bhalla, A.; Prabhakar., N. An electrochemical aptasensor based on graphene doped chitosan nanocomposites for determination of Ochratoxin A. Microchem. J. 2019, 144, 102–109. [Google Scholar] [CrossRef]
- Wang, Y.H.; Ning, G.; Wu, Y.H.; Wu, S.; Zeng, B.Q.; Liu, G.Q.; He, X.X.; Wang, K.M. Facile combination of beta-cyclodextrin host-guest recognition with exonuclease-assistant signal amplification for sensitive electrochemical assay of ochratoxin A. Biosens. Bioelectron. 2019, 124, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Y.; Shan, Y.Q.; Gong, M.; Jin, X.; Lv, L.R.; Jiang, M.; Xu, J. A novel electrochemical sensor for ochratoxin A based on the hairpin aptamer and double report DNA via multiple signal amplification strategy. Sens. Actuators B Chem. 2019, 281, 595–601. [Google Scholar] [CrossRef]
- Wu, K.F.; Ma, C.B.; Zhao, H.; Chen, M.J.; Deng, Z.Y. Sensitive aptamer-based fluorescene assay for ochratoxin A based on RNase H signal amplification. Food Chem. 2019, 277, 273–278. [Google Scholar] [CrossRef]
- Wu, J.; Chu, H.; Mei, Z.; Deng, Y.; Xue, F.; Zheng, L.; Chen, W. Ultrasensitive one-step rapid detection of ochratoxin A by the folding-based electrochemical aptasensor. Anal. Chim. Acta 2012, 753, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.; Jin, Y.; Kang, X.; Zhao, Y.; Cui, C.; Guo, Z. PVP-coated gold nanoparticles for the selective determination of ochratoxin A via quenching fluorescence of the free aptamer. Food Chem. 2018, 249, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Zheng, H.; Sun, M.; Guo, Y.; Luo, F.; Guo, L.; Qiu, B.; Lin, Z.; Chen, G. Highly sensitive colorimetric aptasensor for ochratoxin A detection based on enzyme-encapsulated liposome. Anal. Chim. Acta 2018, 1002, 90–96. [Google Scholar] [CrossRef]
- Bagheri, E.; Abnous, K.; Alibolandi, M.; Ramezani, M.; Taghdisi, S.M. Triple-helix molecular switch-based aptasensors and DNA sensors. Biosens. Bioelectron. 2018, 111, 1–9. [Google Scholar] [CrossRef]
- Zheng, J.; Li, J.S.; Jiang, Y.; Jin, J.Y.; Wang, K.M.; Yang, R.H.; Tan, W.H. Design of aptamer-based sensing platform using triple-helix molecular switch. Anal. Chem. 2011, 83, 6586–6592. [Google Scholar] [CrossRef]
- Xiong, E.H.; Li, Z.Z.; Zhang, X.H.; Zhou, J.W.; Yan, X.X.; Liu, Y.Q.; Chen, J.H. Triple-helix molecular switch electrochemical ratiometric biosensor for ultrasensitive detection of nucleic acids. Anal. Chem. 2017, 89, 8830–8835. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, Y.H.; Ma, H.M.; Ren, X.; Wang, Y.G.; Zhang, Y.; Wei, Q. Electrochemical DNA probe for Hg2+ detection based on a triple-helix DNA and Multistage Signal Amplification Strategy. Biosens. Bioelectron. 2016, 86, 907–912. [Google Scholar] [CrossRef] [PubMed]
- Kohli, R.M.; Zhang, Y. TET enzymes, TDG and the dynamics of DNA demethylation. Nature 2013, 502, 472–479. [Google Scholar] [CrossRef]
- Huang, L.; Tian, S.; Zhao, W.; Liu, K.; Ma, X.; Guo, J. Aptamer-based lateral flow assay on-site biosensors. Biosens. Bioelectron. 2021, 186, 113279. [Google Scholar] [CrossRef] [PubMed]
- Ayela, C.; Roquet, F.; Valera, L.; Granier, C.; Nicu, L.; Pugnière, M. Antibody–antigenic peptide interactions monitored by SPR and QCM-D: A model for SPR detection of IA-2 autoantibodies in human serum. Biosens. Bioelectron. 2007, 22, 3113–3119. [Google Scholar] [CrossRef]
- Sinitsyna, V.V.; Vetcher, A.A. Nucleic Acid Aptamers in Nanotechnology. Biomedicines 2022, 10, 1079. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Midinov, B.; White, R.J. Electrochemical aptamer-based sensor for real-time monitoring of insulin. ACS Sens. 2019, 4, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Angell, C.; Kai, M.; Xie, S.; Dong, X.; Chen, Y. Bioderived DNA nanomachines for potential uses in biosensing, diagnostics, and therapeutic applications. Adv. Healthc. Mater. 2018, 7, 1701189. [Google Scholar] [CrossRef]
- Zhao, J.; Zhai, Q. Recent advances in the development of ligands specifically targeting telomeric multimeric G-quadruplexes. Bioorganic Chem. 2020, 103, 104229. [Google Scholar] [CrossRef]
- Belleperche, M.; DeRosa, M.C. pH-control in aptamer-based diagnostics, therapeutics, and analytical applications. Pharmaceuticals 2018, 11, 80. [Google Scholar] [CrossRef]
- Nguyen, T.Q.N.; Lim, K.W.; Phan, A.T. Folding kinetics of G-quadruplexes: Duplex stem loops drive and accelerate G-quadruplex folding. J. Phys. Chem. B 2020, 124, 5122–5130. [Google Scholar] [CrossRef]
- Cecerska-Heryć, E.; Surowska, O.; Heryć, R.; Serwin, N.; Napiontek-Balińska, S.; Dołęgowska, B. Are antioxidant enzymes essential markers in the diagnosis and monitoring of cancer patients–a review. Clin. Biochem. 2021, 93, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Li, G.; Zou, L.; Lei, S.; Yu, Q.; Ye, B. Highly active DNAzyme-peptide hybrid structure coupled porous palladium for high-performance electrochemical aptasensing platform. Sens. Actuators B Chem. 2018, 259, 372–379. [Google Scholar] [CrossRef]
- Luo, Z.; Sun, D.; Tong, Y.; Zhong, Y.; Chen, Z. DNA nanotetrahedron linked dual-aptamer based voltammetric aptasensor for cardiac troponin I using a magnetic metal-organic framework as a label. Microchim. Acta 2019, 186, 374. [Google Scholar] [CrossRef]
- Sun, D.P.; Lin, X.A.; Lu, J.; Wei, P.; Luo, Z.B.; Lu, X.E.; Chen, Z.G.; Zhang, L.Y. DNA nanotetrahedron-assisted electrochemical aptasensor for cardiac troponin I detection based on the co-catalysis of hybrid nanozyme, natural enzyme and artificial DNAzyme. Biosens. Bioelectron. 2019, 142, 111578. [Google Scholar] [CrossRef] [PubMed]
- Tintoré, M.; Gállego, I.; Manning, B.; Eritja, R.; Fàbrega, C. DNA Origami as a DNA Repair Nanosensor at the Single-Molecule Level. Angew. Chem. 2013, 125, 7901–7904. [Google Scholar] [CrossRef]
- Yuminova, A.V.; Smirnova, I.G.; Arutyunyan, A.M.; Kopylov, A.M.; Golovin, A.V.; Pavlova, G.V. The structure of G-quadruplex thrombine-binding DNA aptamer RA36. Mosc. Univ. Chem. Bull. 2015, 70, 43–46. [Google Scholar] [CrossRef]
- Alieva, R.R.; Zavyalova, E.G.; Tashlitsky, V.N.; Kopylov, A.M. Quantitative characterization of oligomeric state of G-quadruplex antithrombin aptamers by size exclusion HPLC. Mendeleev Commun. 2019, 29, 424–425. [Google Scholar] [CrossRef]
- Zein, S.S.; Vetcher, A.A.; Levene, S. PCR-Based Synthesis of Repetitive Single-Stranded DNA for Applications to Nanobiotechnology. Int. J. Nanosci. 2005, 4, 287–294. [Google Scholar] [CrossRef]
- Kretz, C.K.; Cuddy, K.R.; Stafford, A.R. HD1, a thrombin-and prothrombin-binding DNA aptamer, inhibits thrombin generation by attenuating prothrombin activation and thrombin feedback reactions. Thromb. Haemost. 2010, 103, 83–96. [Google Scholar] [CrossRef]
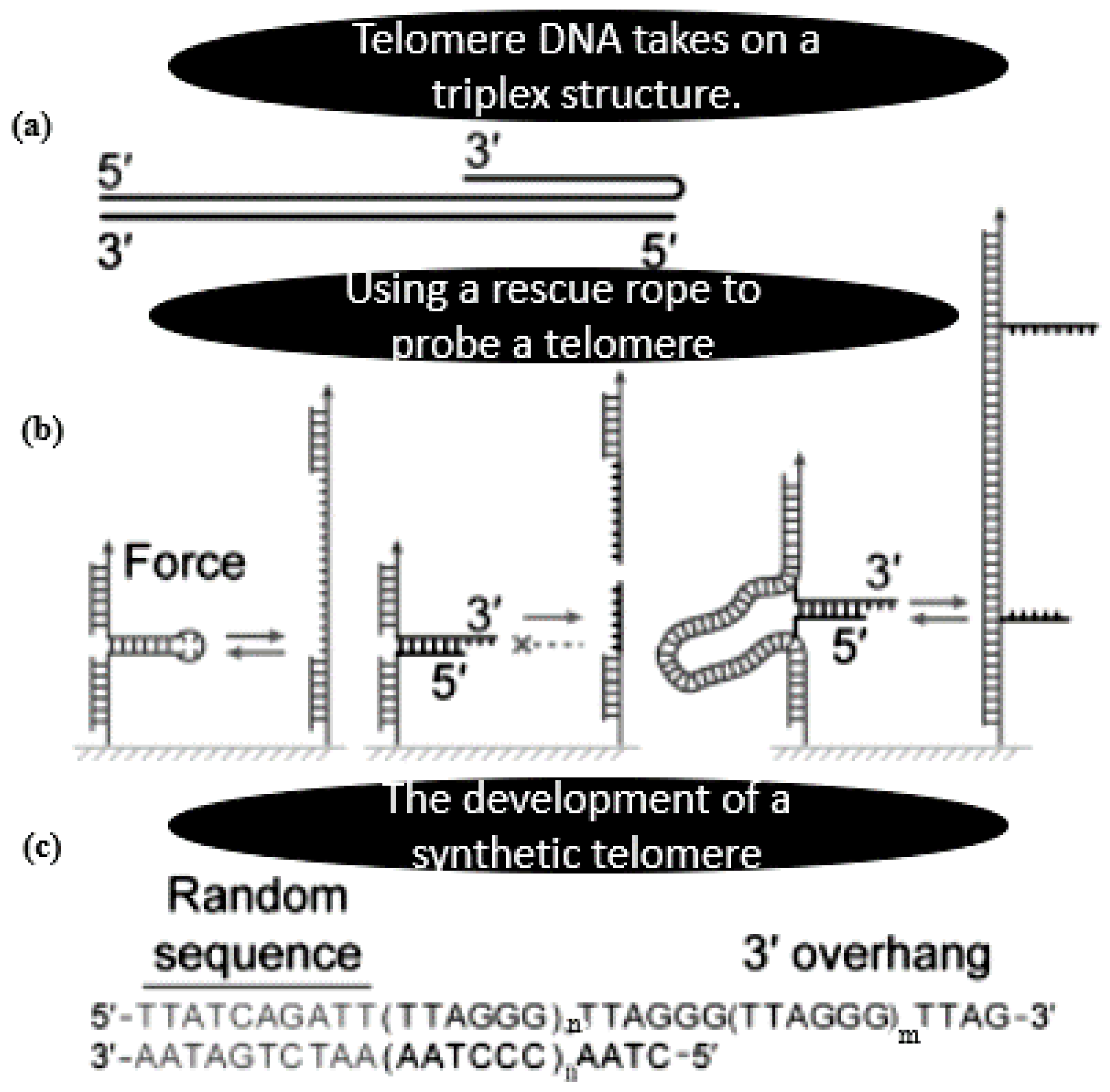
- Li, N.; Wang, J.; Ma, K.; Liang, L.; Mi, L.; Huang, W.; Yu, Z. The dynamics of forming a triplex in an artificial telomere inferred by DNA mechanics. Nucleic Acids Res. 2019, 47, e86. [Google Scholar] [CrossRef]
- Liu, C.P.; Wey, M.T.; Chang, C.C.; Kan, L.S. Direct observation of single molecule conformational change of tight-turn paperclip DNA triplex in solution. Appl. Biochem. Biotechnol. 2009, 159, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.B.; Jia, Q.; Xu, C.H. Asynchrony of base-pair breaking and nucleotide releasing of helicases in DNA unwinding. J. Phys. Chem. B 2018, 122, 5790–5796. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.T.; Ha, T. Direct visualization of helicase dynamics using fluorescence localization and optical trapping. Methods Enzymol. 2017, 582, 121–136. [Google Scholar] [PubMed]
- Lee, J.; Crickard., J.B.; Reese, J.C.; Lee, T.H. Single-molecule Frest method to investigate the dynamics of transcription elongation through the nucleosome by RNA polymerase II. Methods 2019, 159, 51–58. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bekkouche, I.; Shishonin, A.Y.; Vetcher, A.A. Recent Development in Biomedical Applications of Oligonucleotides with Triplex-Forming Ability. Polymers 2023, 15, 858. https://doi.org/10.3390/polym15040858
Bekkouche I, Shishonin AY, Vetcher AA. Recent Development in Biomedical Applications of Oligonucleotides with Triplex-Forming Ability. Polymers. 2023; 15(4):858. https://doi.org/10.3390/polym15040858
Chicago/Turabian StyleBekkouche, Incherah, Alexander Y. Shishonin, and Alexandre A. Vetcher. 2023. "Recent Development in Biomedical Applications of Oligonucleotides with Triplex-Forming Ability" Polymers 15, no. 4: 858. https://doi.org/10.3390/polym15040858
APA StyleBekkouche, I., Shishonin, A. Y., & Vetcher, A. A. (2023). Recent Development in Biomedical Applications of Oligonucleotides with Triplex-Forming Ability. Polymers, 15(4), 858. https://doi.org/10.3390/polym15040858