
Polysaccharides—Naturally Occurring Immune Modulators

 , , ,
, , ,  ,
,  and
and 
Abstract
1. Introduction
2. Polysaccharides
Polysaccharide Classification
3. Extraction of Polysaccharides
3.1. Extraction of β-Glucans
3.2. Extraction of Fucoidan
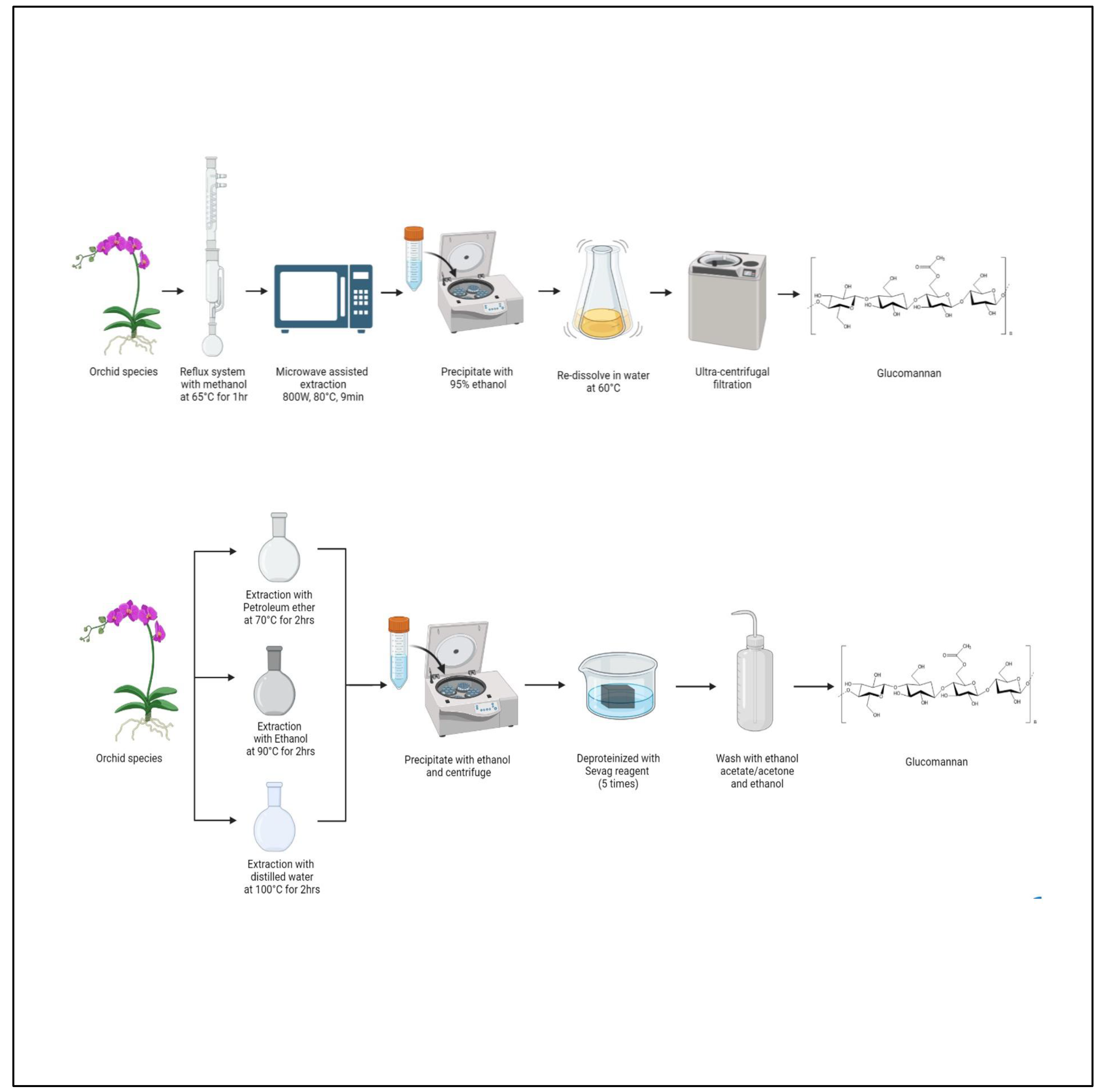
3.3. Extraction of Glucomannan
3.4. Extraction of Chitosan
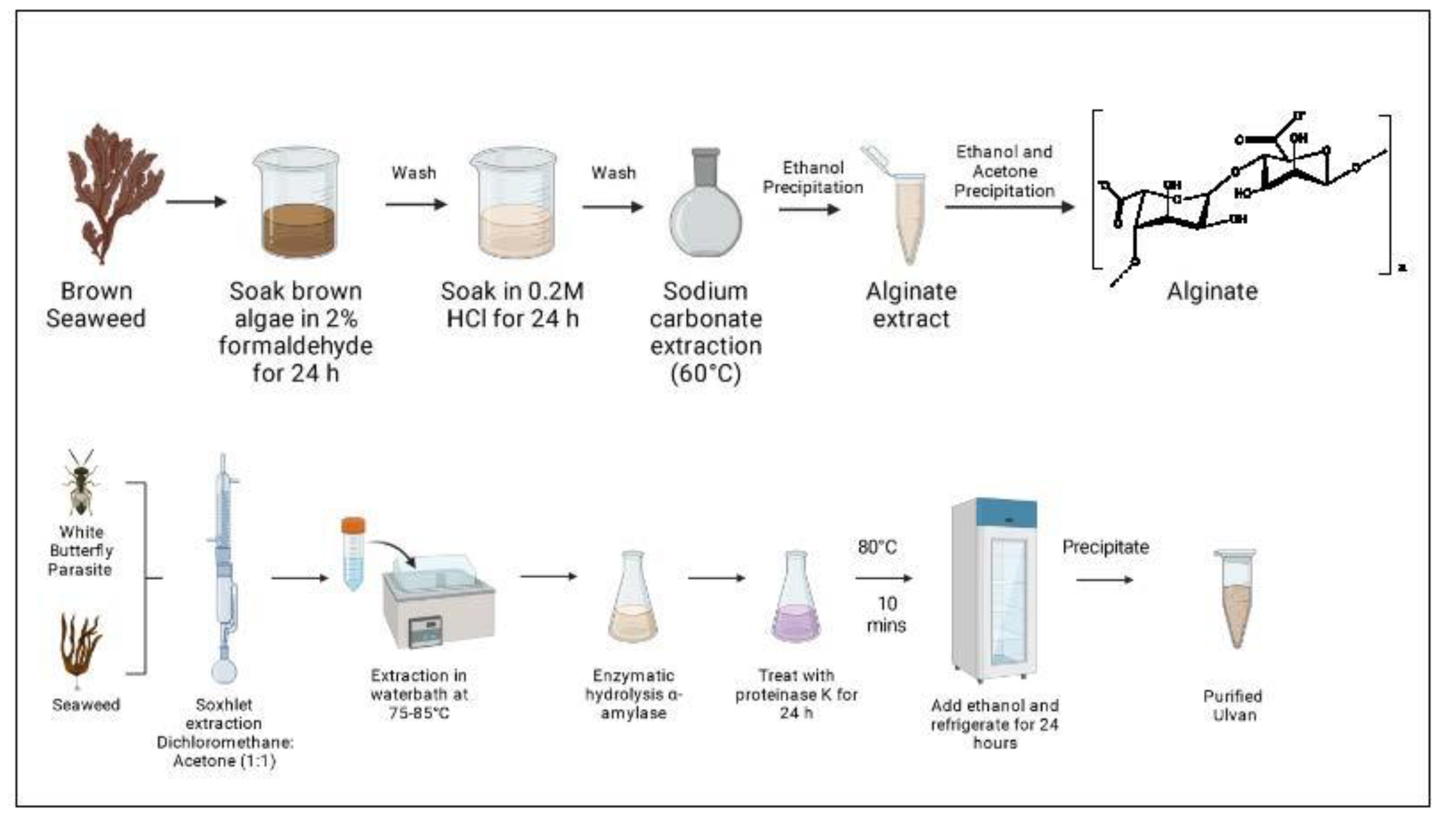
3.5. Extraction of Alginate
3.6. Extraction of Hyaluronic acid
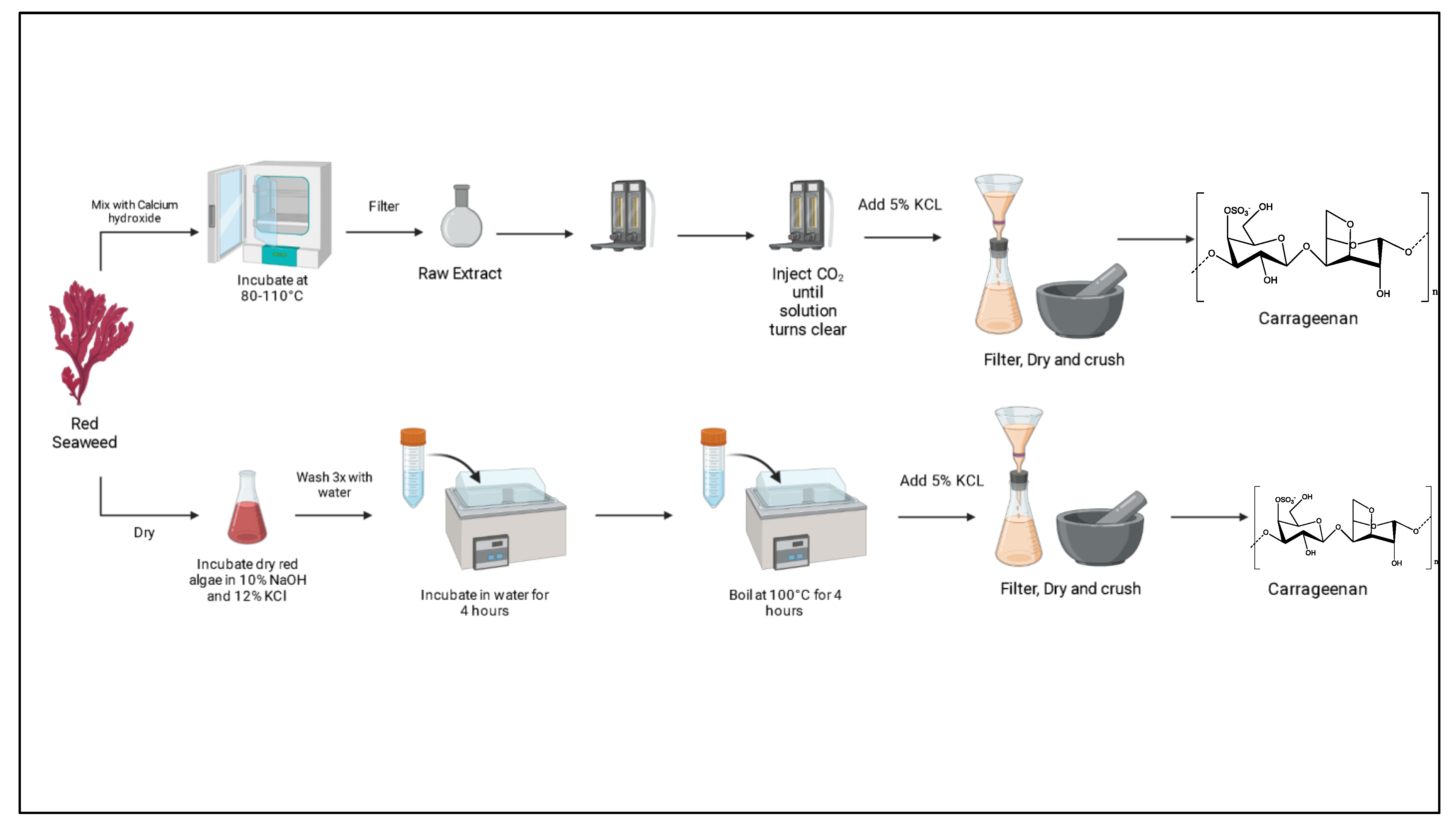
3.7. Extraction of Carrageenan
3.8. Extraction of Ulvan
3.9. Extraction of Xylans
4. Immune Modulation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polysaccharide | Model | Immuno-Modulatory Effect | Reference |
|---|---|---|---|
| Fucoidan | In vitro cell culture, | Activates NF-κB in macrophages (in vitro). Promotes maturation of dendritic cells. Inhibits the polarization of macrophages toward the tumor-promoting M2 phenotype (in vitro). | [142] |
| Fucoidan Undariia pinnatifilda | In vitro cell culture—neutrophils. | Delayed spontaneous apoptosis is associated with increased levels of anti-apoptotic protein Mcl-1 and decreased levels of activated caspase -3. Induced activation of neutrophil–secretion of IL-6, IL-8, and TNF-α–AKT-dependent manner. | [143] |
| Fucoidan Macrocystis pyrifera | In vitro cell culture—human neutrophils, mouse NK cells, spleen dendritic cells, and T-cell in vivo mouse model. | Delayed human neutrophil apoptosis. Promoted NK cells activation and cytotoxic activity. Promoted DC maturation. Enhanced T-cell immune response, antigen-specific antibody production, and memory T-cell generation. | [115] |
| Fucoidan Fucus vesiculosus | In vivo mouse model. | Upregulation of CD40, CD80, and CD86 expression. Production of IL-6, IL-12, and TNF-α in spleen DCs. Influenced INF-γ-producing Th1 and Tc1 cells in an IL-12-dependent manner. | [144] |
| Fucoidan Clad siphon navae-caledoniae | In vitro cell culture. Estrogen-positive and estrogen-negative breast cells. | Co-treatments with chemotherapy drugs inhibited cell growth, apoptosis, and cell-cycle modifications. Decrease in phosphorylation of ERK and Akt. Enhanced intracellular ROS levels. Reduced glutathione levels. | [145] |
| Fucoidan commercial powder 10 mg/mL | Prospective open-label, single-arm clinical study. | Reduction in pro-inflammatory cytokines IL-1β. | [146] |
| Fucoidan Kjellmaniella crassifolia and Undaria pinnatifida | In vitro macrophages RAW264.7. In vivo mouse model. | Enhanced cell proliferation enhanced the secretion of granulocyte-macrophage-colony-stimulating factor (GM-CSF) and tumor necrosis factor-α (TNF-α). Increased the secretion of GM-CSF, TNF-α, interleukin (IL)-4 and IL-10 in vivo. | [147] |
| Fucoidan Sargassumspecies andFucus vesiculosus | Lewis lung carcinoma cells and melanoma B16 cells. In vivo mouse model. | Decreased the viable number of cancerous cells in a dose–response manner (in vitro). Cytolytic activity of natural killer (NK) cells was enhanced (in vivo). | [148] |
| Chitosan | Isolated spleens from oligodendrocyte glycoprotein (MOG) induced experimental autoimmune encephalomyelitis (EAE) mice. | Increases INF-γ and IL-10 levels. | [149] |
| Chitosan | Murine model of autoimmune encephalomyelitis. | Improvement in clinical signs. Reduction in demyelination. INF-γ, IL-17, and TNF-α levels reduced. | [150] |
| Chitosan | In vitro cell culture. | Induces production of TNF-α, IL-6, and INF-γ in macrophages. Promote the expression of the genes, including iNOS and TNF-α. | [151] |
| Chitosan | Leishmania infection in BALB/c mice. | Increases INF-γ secretion. Reduction in lesion formation. Lower parasite load. | [152] |
| Alginate (Commercial) | In vitro cell culture. | Increases TLR-4 expression. Activates NF-ĸB and MAPK pathways. Promote TLR-4-mediated phagocytosis. | [153] |
| Xylans Corn cobs | Dextran sodium sulphate (DSS)-induced UC mouse model. | Reversed the imbalance between pro- and anti-inflammatory cytokines. Rebalanced gut microbiota and reduced Oscillibacter, Ruminococcaceae, Erysipelatoclostridium, and Defluviitaleaceae nuclear factor-κB (NF-κB) inhibition. Reduction of inflammatory intestinal damage. | [154] |
| Commercial λ-Carrageenan | Melanoma B16-F10 and mammary cancer 4T1 mouse models. | Inhibited tumor growth. Enhanced tumor immune response. Increased the number of tumor-infiltrating M1 macrophages and dendritic cells. Enhanced the secretion of IL17A in spleen. Increased the level of TNF-α in tumor. | [155] |
| Ulvan | Labeo rohita | Increase in red blood cells and white blood cells. Increase in superoxide dismutase activity. Increase in respiratory burst activity. Increase in phagocytic activity. Increase in expression IL-1β, TNFα, hepcidin, and IgM cytokines. | [156] |
| Hyaluronic acid | Clinical trial Patients of knee osteoarthritis. | Lymphocyte number increased. Reduction of IL-6 and IL-8. | [157] |
| In vitro cell culture–murine macrophages. | Nitric oxide production by LPS-stimulated macrophages was decreased. Decreased expression of TNF-α, IL-6, CCL2, and IL-1β in LPS-stimulated macrophages. Increased expression of TGF-β1, IL10, IL-11, and Arg1 genes/anti-inflammatory responses. | [158] | |
| Glucomannan Amorphophallus konjac | Cyclophosphamide -induced immunosuppressive mice. | Enhanced the spleen indices. Enhanced the thymus indices. Reduced the proliferation of splenic lymphocytes. Enhanced and regulated humoral immune activity via serum hemolysin. Reduce the capacity of NK cell lethality. Reduction of phagocytic activity of peritoneal macrophages was extremely significantly reduced. Reduction in the production of IL-2, IgG, and TNF-α. | [159] |
| Glucomannan Dendrobium officinale | In vitro cell culture. | Stimulate cytokine production (TNF-α and IL-1β). Induced immune activities involving ERK1/2 and NF-κB pathways. | [160] |
| Glucomannan Dendrobium officinale | In vitro cell culture. | Promoted the degradation of IκB complexes and activated NF-κB phosphorylation. | [161] |
| β-glucans Paramylon E. gracillis | In vivo mouse model (wound healing). | Increased wound contraction. Increased IFN-γ levels. | [162] |
| β-glucans Paramylon E. gracillis | In vivo mouse model (influenza infected). | Increased survival rate. Lower virus titer compared to control group. Increased levels of cytokines-IL-1β, IL-6, IL-12, IL-10, IFN-γ, and TNF-α. | [163] |
| β-glucans Lentinan E. gracillis | In vitro cell culture. | Increased levels of IL-6, TNF-α IL-22, IL-8, and IL-10 in THP-1 macrophages. IL-8 and TNF-α reduction after cytokine insult in A549 and BEAS-2B lung cells. | [128] |
5. Natural Polysaccharides with Immune-Modulatory Activity
5.1. β-Glucans
Immuno-Modulatory Activity of Beta Glucans
5.2. Fucoidan
Immuno-Modulatory Activity of Fucoidan
5.3. Glucomannan
Immuno-Modulatory Activity of Glucomannan
5.4. Chitosan
Immuno-Modulatory Activity of Chitosan
5.5. Alginate
Immuno-Modulatory Activity of Alginate
5.6. Hyaluronic Acid
Immuno-Modulatory Activity of Hyaluronic acid
5.7. Carrageenan
Immuno-Modulatory Activity of Carrageenan
5.8. Xylans
Immuno-Modulatory Activity of Xylans
5.9. Ulvan
Immuno-Modulatory Activity of Ulvan
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Vishwakarma, A.; Bhise, N.S.; Evangelista, M.B.; Rouwkema, J.; Dokmeci, M.R.; Ghaemmaghami, A.M.; Vrana, N.E.; Khademhosseini, A. Engineering Immunomodulatory Biomaterials To Tune the Inflammatory Response. Trends Biotechnol. 2016, 34, 470–482. [Google Scholar] [CrossRef] [PubMed]
- Wiles, M.K.L.; Fishman, J.M.; De Coppi, P.; Birchall, M.A. The Host Immune Response to Tissue-Engineered Organs: Current Problems and Future Directions. Tissue Eng. Part B Rev. 2016, 22, 208–219. [Google Scholar] [CrossRef]
- Roseti, L.; Parisi, V.; Petretta, M.; Cavallo, C.; Desando, G.; Bartolotti, I.; Grigolo, B. Scaffolds for Bone Tissue Engineering: State of the art and new perspectives. Mater. Sci. Eng. C 2017, 78, 1246–1262. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.; Moxon, S.; Morris, G. Biopolymers as wound healing materials. In Wound Healing Biomaterials; Ågren, M.S., Ed.; Woodhead Publishing: Cambridge, UK, 2016; pp. 261–287. [Google Scholar] [CrossRef]
- Vishakha, S.K.; Kishor, D.B.; Sudha, S.R. Natural Polymers-A Comprehensive Review. Int. J. Res. Pharm. Biomed. Sci. 2012, 3, 1597–1613. [Google Scholar]
- He, W.; Benson, R. Polymeric Biomaterials. In Applied Plastics Engineering Handbook; William Andrew Publishing: Norwich, NY, USA, 2017; pp. 145–164. [Google Scholar] [CrossRef]
- Maitz, M. Applications of synthetic polymers in clinical medicine. Biosurface Biotribology 2015, 1, 161–176. [Google Scholar] [CrossRef]
- Hacker, M.C.; Krieghoff, J.; Mikos, A.G. Synthetic Polymers. In Principles of Regenerative Medicine; Academic Press: Cambridge, MA, USA, 2018; pp. 559–590. [Google Scholar] [CrossRef]
- Major, I.; Fuenmayor, E.; McConville, C. The Production of Solid Dosage Forms from Non-Degradable Polymers. Curr. Pharm. Des. 2016, 22, 2738–2760. [Google Scholar] [CrossRef] [PubMed]
- Swetha, M.; Sahithi, K.; Moorthi, A.; Srinivasan, N.; Ramasamy, K.; Selvamurugan, N. Biocomposites containing natural polymers and hydroxyapatite for bone tissue engineering. Int. J. Biol. Macromol. 2010, 47, 1–4. [Google Scholar] [CrossRef]
- Nair, L.S.; Laurencin, C.T. Biodegradable polymers as biomaterials. Prog. Polym. Sci. 2007, 32, 762–798. [Google Scholar] [CrossRef]
- Surin, S.; Surayot, U.; Seesuriyachan, P.; You, S.; Phimolsiripol, Y. Antioxidant and immunomodulatory activities of sulphated polysaccharides from purple glutinous rice bran (Oryza sativa L.). Int. J. Food Sci. Technol. 2017, 53, 994–1004. [Google Scholar] [CrossRef]
- Shi, Y.; Xiong, Q.; Wang, X.; Li, X.; Yu, C.; Wu, J.; Yi, J.; Zhao, X.; Xu, Y.; Cui, H. Characterization of a novel purified polysaccharide from the flesh of Cipangopaludina chinensis. Carbohydr. Polym. 2016, 136, 875–883. [Google Scholar] [CrossRef]
- Navarro, D.M.D.L.; Abelilla, J.J.; Stein, H.H. Structures and characteristics of carbohydrates in diets fed to pigs: A review. J. Anim. Sci. Biotechnol. 2019, 10, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, J.R.; Junior, R.N.D.C. Polysaccharides obtained from natural edible sources and their role in modulating the immune system: Biologically active potential that can be exploited against COVID-19. Trends Food Sci. Technol. 2021, 108, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Rokstad, A.M.A.; Lacík, I.; de Vos, P.; Strand, B.L. Advances in biocompatibility and physico-chemical characterization of microspheres for cell encapsulation. Adv. Drug Deliv. Rev. 2014, 67–68, 111–130. [Google Scholar] [CrossRef]
- De Vos, P.; Lazarjani, H.A.; Poncelet, D.; Faas, M.M. Polymers in cell encapsulation from an enveloped cell perspective. Adv. Drug Deliv. Rev. 2014, 67–68, 15–34. [Google Scholar] [CrossRef]
- Zhang, Y.; Lin, L.; Liu, L.; Liu, F.; Maruyama, A.; Tian, H.; Chen, X. Ionic-crosslinked polysaccharide/PEI/DNA nanoparticles for stabilized gene delivery. Carbohydr. Polym. 2018, 201, 246–256. [Google Scholar] [CrossRef]
- Han, J.; Cai, J.; Borjihan, W.; Ganbold, T.; Rana, T.M.; Baigude, H. Preparation of novel curdlan nanoparticles for intracellular siRNA delivery. Carbohydr. Polym. 2014, 117, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L.D.P.; Mota, M.R.; Brizeno, L.A.; Nogueira, F.C.; Ferreira, E.G.; Pereira, M.G.; Assreuy, A.M. Modulator effect of a polysaccharide-rich extract from Caesalpinia ferrea stem barks in rat cutaneous wound healing: Role of TNF-α, IL-1β, NO, TGF-β. J. Ethnopharmacol. 2016, 187, 213–223. [Google Scholar] [CrossRef]
- Tabandeh, M.R.; Oryan, A.; Mohammadalipour, A. Polysaccharides of Aloe vera induce MMP-3 and TIMP-2 gene expression during the skin wound repair of rat. Int. J. Biol. Macromol. 2014, 65, 424–430. [Google Scholar] [CrossRef]
- Chi, N.-H.; Yang, M.-C.; Chung, T.-W.; Chou, N.-K.; Wang, S.-S. Cardiac repair using chitosan-hyaluronan/silk fibroin patches in a rat heart model with myocardial infarction. Carbohydr. Polym. 2013, 92, 591–597. [Google Scholar] [CrossRef]
- Kumar, A.; Rao, K.M.; Han, S.S. Application of xanthan gum as polysaccharide in tissue engineering: A review. Carbohydr. Polym. 2018, 180, 128–144. [Google Scholar] [CrossRef]
- Zohuri, G.H.; Sandaroos, R.; Ahmadjo, S.; Damavandi, S.; Rabiee, A.; Shamekhi, M.A. Tissue Engineering: Biomaterial Application. In Encyclopedia of Biomedical Polymers and Polymeric Biomaterials; CRC Press: Boca Raton, FL, USA, 2016; pp. 7901–7932. [Google Scholar] [CrossRef]
- Silva, D.; Pinto, L.F.; Bozukova, D.; Santos, L.F.; Serro, A.; Saramago, B. Chitosan/alginate based multilayers to control drug release from ophthalmic lens. Colloids Surf. B Biointerfaces 2016, 147, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Badshah, S.; Riaz, A.; Muhammad, A.; Çayan, G.T.; Çayan, F.; Duru, M.E.; Ahmad, N.; Emwas, A.-H.; Jaremko, M. Isolation, Characterization, and Medicinal Potential of Polysaccharides of Morchella esculenta. Molecules 2021, 26, 1459. [Google Scholar] [CrossRef] [PubMed]
- Sheng, K.; Wang, C.; Chen, B.; Kang, M.; Wang, M.; Chemistry, K.L.-F. Undefined Recent Advances in Polysaccharides from Lentinus edodes (Berk.): Isolation, Structures and Bioactivities; Elsevier: Amsterdam, The Netherlands, 2021. [Google Scholar]
- Ohmes, J.; Mikkelsen, M.D.; Nguyen, T.T.; Tran, V.H.N.; Meier, S.; Nielsen, M.S.; Ding, M.; Seekamp, A.; Meyer, A.S.; Fuchs, S. Depolymerization of fucoidan with endo-fucoidanase changes bioactivity in processes relevant for bone regeneration. Carbohydr. Polym. 2022, 286, 119286. [Google Scholar] [CrossRef] [PubMed]
- Ranjbari, J.; Mokhtarzadeh, A.; Alibakhshi, A.; Tabarzad, M.; Hejazi, M.; Ramezani, M. Anti-Cancer Drug Delivery Using Carbohydrate-Based Polymers. Curr. Pharm. Des. 2018, 23, 6019–6032. [Google Scholar] [CrossRef]
- Hao, W.; Wang, S.-F.; Zhao, J.; Li, S.-P. Effects of extraction methods on immunology activity and chemical profiles of Lycium barbarum polysaccharides. J. Pharm. Biomed. Anal. 2020, 185, 113219. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, G.; Lai, F.; Wu, H. Structural Characterization and Immunomodulatory Activity of a Novel Polysaccharide from Lepidium meyenii. J. Agric. Food Chem. 2016, 64, 1921–1931. [Google Scholar] [CrossRef]
- Thomas, S.; Rezoagli, E.; Abidin, I.Z.; Major, I.; Murray, P.; Murphy, E.J. β-Glucans from Yeast—Immunomodulators from Novel Waste Resources. Appl. Sci. 2022, 12, 5208. [Google Scholar] [CrossRef]
- Murphy, E.J.; Rezoagli, E.; Major, I.; Rowan, N.J.; Laffey, J.G. β-Glucan Metabolic and Immunomodulatory Properties and Potential for Clinical Application. J. Fungi 2020, 6, 356. [Google Scholar] [CrossRef]
- Chaisuwan, W.; Jantanasakulwong, K.; Wangtueai, S.; Phimolsiripol, Y.; Chaiyaso, T.; Techapun, C.; Phongthai, S.; You, S.; Regenstein, J.M.; Seesuriyachan, P. Microbial exopolysaccharides for immune enhancement: Fermentation, modifications and bioactivities. Food Biosci. 2020, 35, 100564. [Google Scholar] [CrossRef]
- Nwodo, U.U.; Green, E.; Okoh, A.I. Bacterial Exopolysaccharides: Functionality and Prospects. Int. J. Mol. Sci. 2012, 13, 14002–14015. [Google Scholar] [CrossRef]
- Wang, J.; Salem, D.R.; Sani, R.K. Extremophilic exopolysaccharides: A review and new perspectives on engineering strategies and applications. Carbohydr. Polym. 2018, 205, 8–26. [Google Scholar] [CrossRef]
- Lordan, S.; Ross, R.P.; Stanton, C. Marine Bioactives as Functional Food Ingredients: Potential to Reduce the Incidence of Chronic Diseases. Mar. Drugs 2011, 9, 1056–1100. [Google Scholar] [CrossRef] [PubMed]
- Ahnen, R.T.; Jonnalagadda, S.S.; Slavin, J.L. Role of plant protein in nutrition, wellness, and health. Nutr. Rev. 2019, 77, 735–747. [Google Scholar] [CrossRef] [PubMed]
- Yockey, L.J.; Lucas, C.; Iwasaki, A. Contributions of maternal and fetal antiviral immunity in congenital disease. Science 2020, 368, 608–612. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Belwal, T.; Cravotto, G.; Luo, Z. Sono-physical and sono-chemical effects of ultrasound: Primary applications in extraction and freezing operations and influence on food components. Ultrason. Sonochemistry 2020, 60, 104726. [Google Scholar] [CrossRef] [PubMed]
- Vaclavik, V.; Christian, E.; Campbell, T. Essentials of Food Science; Springer: Berlin/Heidelberg, Germany, 2008; Volume 45. [Google Scholar]
- Kennedy, J.F.; Quinton, L. Essentials of Carbohydrate Chemistry and Biochemistry: Thisbe K. Lindhorst. Wiley & Sons Ltd, 2000, 217pp, £34.95p, ISBN 3-527-29543-7. Carbohydr. Polym. 2002, 47, 87. [Google Scholar] [CrossRef]
- Li, Q.; Niu, Y.; Xing, P.; Wang, C. Bioactive polysaccharides from natural resources including Chinese medicinal herbs on tissue repair. Chin. Med. 2018, 13, 1–11. [Google Scholar] [CrossRef]
- Dalheim, M.; Arnfinnsdottir, N.B.; Widmalm, G.; Christensen, B.E. The size and shape of three water-soluble, non-ionic polysaccharides produced by lactic acid bacteria: A comparative study. Carbohydr. Polym. 2016, 142, 91–97. [Google Scholar] [CrossRef]
- Sindhu, R.K.; Goyal, A.; Das, J.; Neha; Choden, S.; Kumar, P. Immunomodulatory potential of polysaccharides derived from plants and microbes: A narrative review. Carbohydr. Polym. Technol. Appl. 2021, 2, 100044. [Google Scholar] [CrossRef]
- Mohammed, A.S.A.; Naveed, M.; Jost, N. Polysaccharides; Classification, Chemical Properties, and Future Perspective Applications in Fields of Pharmacology and Biological Medicine (A Review of Current Applications and Upcoming Potentialities). J. Polym. Environ. 2021, 29, 2359–2371. [Google Scholar] [CrossRef]
- Pettolino, F.; Walsh, C.; Fincher, G.B.; Bacic, A. Determining the polysaccharide composition of plant cell walls. Nat. Protoc. 2012, 7, 1590–1607. [Google Scholar] [CrossRef] [PubMed]
- Mudgil, D.; Barak, S. Composition, properties and health benefits of indigestible carbohydrate polymers as dietary fiber: A review. Int. J. Biol. Macromol. 2013, 61, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Sami, N.; Ahmad, R.; Fatma, T. Exploring algae and cyanobacteria as a promising natural source of antiviral drug against SARS-CoV-2. Biomed. J. 2020, 44, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Mourelle, M.L.; Gómez, C.P.; Legido, J.L. The Potential Use of Marine Microalgae and Cyanobacteria in Cosmetics and Thalassotherapy. Cosmetics 2017, 4, 46. [Google Scholar] [CrossRef]
- Huang, L.; Shen, M.; Morris, G.A.; Xie, J. Sulfated polysaccharides: Immunomodulation and signaling mechanisms. Trends Food Sci. Technol. 2019, 92, 1–11. [Google Scholar] [CrossRef]
- Riccio, G.; Lauritano, C. Microalgae with Immunomodulatory Activities. Mar. Drugs 2019, 18, 2. [Google Scholar] [CrossRef]
- Peasura, N.; Laohakunjit, N.; Kerdchoechuen, O.; Vongsawasdi, P.; Chao, L.K. Assessment of biochemical and immunomodulatory activity of sulphated polysaccharides from Ulva intestinalis. Int. J. Biol. Macromol. 2016, 91, 269–277. [Google Scholar] [CrossRef]
- Li, J.; Wang, Y.; Ji, J.; Cao, L.; Bai, Y.; Gao, J. Structural characterization and immunomodulatory activity of a glucan from Radix Codonopsis. J. Funct. Foods 2021, 83, 104537. [Google Scholar] [CrossRef]
- Sanjeewa, K.A.; Lee, J.-S.; Kim, W.-S.; Jeon, Y.-J. The potential of brown-algae polysaccharides for the development of anticancer agents: An update on anticancer effects reported for fucoidan and laminaran. Carbohydr. Polym. 2017, 177, 451–459. [Google Scholar] [CrossRef]
- Jesumani, V.; Du, H.; Pei, P.; Zheng, C.; Cheong, K.L.; Huang, N. Unravelling property of polysaccharides from Sargassum sp. as an anti-wrinkle and skin whitening property. Int. J. Biol. Macromol. 2019, 140, 216–224. [Google Scholar] [CrossRef]
- Venkatesan, J.; Bhatnagar, I.; Manivasagan, P.; Kang, K.-H.; Kim, S.-K. Alginate composites for bone tissue engineering: A review. Int. J. Biol. Macromol. 2015, 72, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Jiang, N.; Li, B.; Wan, M.; Chang, X.; Liu, H.; Zhang, L.; Yin, S.; Qi, H.; Liu, S. Antioxidant activity of purified ulvan in hyperlipidemic mice. Int. J. Biol. Macromol. 2018, 113, 971–975. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, W.; Hao, C.; Yunjia, Y.; Qin, L.; He, M.; Mao, W. Antiviral activity against enterovirus 71 of sulfated rhamnan isolated from the green alga Monostroma latissimum. Carbohydr. Polym. 2018, 200, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Tziveleka, L.-A.; Ioannou, E.; Roussis, V. Ulvan, a bioactive marine sulphated polysaccharide as a key constituent of hybrid biomaterials: A review. Carbohydr. Polym. 2019, 218, 355–370. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, H.; Zhang, K.; Lu, Y.; Wu, Q.; Chen, J.; Li, Y.; Wu, Q.; Chen, Y. Extraction, purification, structural characterization, and gut microbiota relationship of polysaccharides: A review. Int. J. Biol. Macromol. 2022, 213, 967–986. [Google Scholar] [CrossRef]
- Li, J.; Cai, C.; Zheng, M.; Hao, J.; Wang, Y.; Hu, M.; Fan, L.; Yu, G. Alkaline Extraction, Structural Characterization, and Bioactivities of (1→6)-β-d-Glucan from Lentinus edodes. Molecules 2019, 24, 1610. [Google Scholar] [CrossRef]
- Amer, E.M.; Saber, S.H.; Markeb, A.A.; Elkhawaga, A.A.; Mekhemer, I.M.A.; Zohri, A.-N.A.; Abujamel, T.S.; Harakeh, S.; Abd-Allah, E.A. Enhancement of β-Glucan Biological Activity Using a Modified Acid-Base Extraction Method from Saccharomyces cerevisiae. Molecules 2021, 26, 2113. [Google Scholar] [CrossRef]
- Puițel, A.C.; Suditu, G.D.; Danu, M.; Ailiesei, G.-L.; Nechita, M.T. An Experimental Study on the Hot Alkali Extraction of Xylan-Based Hemicelluloses from Wheat Straw and Corn Stalks and Optimization Methods. Polymers 2022, 14, 1662. [Google Scholar] [CrossRef]
- Rodríguez-Sanz, A.; Fuciños, C.; Torrado, A.M.; Rúa, M.L. Extraction of the wheat straw hemicellulose fraction assisted by commercial endo-xylanases. Role of the accessory enzyme activities. Ind. Crop. Prod. 2022, 179, 114655. [Google Scholar] [CrossRef]
- Kang, D.Y.; Kim, W.-S.; Heo, I.S.; Park, Y.H.; Lee, S. Extraction of hyaluronic acid (HA) from rooster comb and characterization using flow field-flow fractionation (FlFFF) coupled with multiangle light scattering (MALS). J. Sep. Sci. 2010, 33, 3530–3536. [Google Scholar] [CrossRef]
- Karami, M.; Shahraky, M.K.; Ranjbar, M.; Tabandeh, F.; Morshedi, D.; Aminzade, S. Preparation, purification, and characterization of low-molecular-weight hyaluronic acid. Biotechnol. Lett. 2020, 43, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Al-Manhel, A.J.; Al-Hilphy, A.R.S.; Niamah, A.K. Extraction of chitosan, characterisation and its use for water purification. J. Saudi Soc. Agric. Sci. 2018, 17, 186–190. [Google Scholar] [CrossRef]
- Triunfo, M.; Tafi, E.; Guarnieri, A.; Salvia, R.; Scieuzo, C.; Hahn, T.; Zibek, S.; Gagliardini, A.; Panariello, L.; Coltelli, M.B.; et al. Characterization of chitin and chitosan derived from Hermetia illucens, a further step in a circular economy process. Sci. Rep. 2022, 12, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Rafiquzzaman, S.M.; Ahmed, R.; Lee, J.M.; Noh, G.; Jo, G.-A.; Kong, I.-S. Improved methods for isolation of carrageenan from Hypnea musciformis and its antioxidant activity. J. Appl. Phycol. 2015, 28, 1265–1274. [Google Scholar] [CrossRef]
- Liu, Y.; An, D.; Xiao, Q.; Chen, F.; Zhang, Y.; Weng, H.; Xiao, A. A novel κ-carrageenan extracting process with calcium hydroxide and carbon dioxide. Food Hydrocoll. 2022, 127, 107507. [Google Scholar] [CrossRef]
- Pankiewicz, R.; Łęska, B.; Messyasz, B.; Fabrowska, J.; Sołoducha, M.; Pikosz, M. First isolation of polysaccharidic ulvans from the cell walls of freshwater algae. Algal Res. 2016, 19, 348–354. [Google Scholar] [CrossRef]
- Hardouin, K.; Bedoux, G.; Burlot, A.S.; Donnay-Moreno, C.; Bergé, J.P.; Nyvall-Collén, P.; Bourgougnon, N. Enzyme-assisted extraction (EAE) for the production of antiviral and antioxidant extracts from the green seaweed Ulva armoricana (Ulvales, Ulvophyceae). Algal Res. 2016, 16, 233–239. [Google Scholar] [CrossRef]
- Torres, M.R.; Sousa, A.P.A.; Silva Filho, E.A.T.; Melo, D.F.; Feitosa, J.P.A.; de Paula, R.C.M.; Lima, M.G.S. Extraction and physicochemical characterization of Sargassum vulgare alginate from Brazil. Carbohydr. Res. 2007, 342, 2067–2074. [Google Scholar] [CrossRef]
- Chee, S.-Y.; Wong, P.-K.; Wong, C.-L. Extraction and characterisation of alginate from brown seaweeds (Fucales, Phaeophyceae) collected from Port Dickson, Peninsular Malaysia. J. Appl. Phycol. 2010, 23, 191–196. [Google Scholar] [CrossRef]
- Hanjabam, M.D.; Kumar, A.; Tejpal, C.S.; Krishnamoorthy, E.; Kishore, P.; Ashok Kumar, K. Isolation of Crude Fucoidan from Sargassum wightii Using Conventional and Ultra-Sonication Extraction Methods. Bioact. Carbohydr. Diet. Fibre 2019, 20, 100200. [Google Scholar] [CrossRef]
- January, G.; Naidoo, R.; Kirby-McCullough, B.; Bauer, R. Assessing methodologies for fucoidan extraction from South African brown algae. Algal Res. 2019, 40, 101517. [Google Scholar] [CrossRef]
- Fan, H.; Meng, Q.; Xiao, T.; Zhang, L. Partial characterization and antioxidant activities of polysaccharides sequentially extracted from Dendrobium officinale. J. Food Meas. Charact. 2018, 12, 1054–1064. [Google Scholar] [CrossRef]
- Deng, Y.; Li, M.; Chen, L.-X.; Chen, X.-Q.; Lu, J.-H.; Zhao, J.; Li, S.-P. Chemical characterization and immunomodulatory activity of acetylated polysaccharides from Dendrobium devonianum. Carbohydr. Polym. 2018, 180, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, J.K.; Mahdi, A.A.; Ahmed, M.I.; Ma, M.; Wang, H. Preparation, deproteinization, characterization, and antioxidant activity of polysaccharide from Medemia argun fruit. Int. J. Biol. Macromol. 2019, 155, 919–926. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, S.; Nie, S.-P.; Cui, S.W.; Wang, Q.; Phillips, A.O.; Phillips, G.O.; Xie, M. Structural Characterization and Chain Conformation of Water-Soluble β-Glucan from Wild Cordyceps sinensis. J. Agric. Food Chem. 2019, 67, 12520–12527. [Google Scholar] [CrossRef]
- Xiao, Z.; Zhang, Q.; Dai, J.; Wang, X.; Yang, Q.; Cai, C.; Mao, J.; Ge, Q. Structural characterization, antioxidant and antimicrobial activity of water-soluble polysaccharides from bamboo (Phyllostachys pubescens Mazel) leaves. Int. J. Biol. Macromol. 2019, 142, 432–442. [Google Scholar] [CrossRef]
- Zeng, X.; Li, P.; Chen, X.; Kang, Y.; Xie, Y.; Li, X.; Xie, T.; Zhang, Y. Effects of deproteinization methods on primary structure and antioxidant activity of Ganoderma lucidum polysaccharides. Int. J. Biol. Macromol. 2018, 126, 867–876. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, G.; Lv, M. Extraction, characterization and antioxidant activities of mannan from yeast cell wall. Int. J. Biol. Macromol. 2018, 118, 952–956. [Google Scholar] [CrossRef]
- Hromádková, Z.; Ebringerová, A.; Sasinková, V.; Šandula, J.; Hříbalová, V.; Omelková, J. Influence of the drying method on the physical properties and immunomodulatory activity of the particulate (1→3)-β-d-glucan from Saccharomyces cerevisiae. Carbohydr. Polym. 2003, 51, 9–15. [Google Scholar] [CrossRef]
- De Jesus Raposo, M.F.; De Morais, A.M.B.; Santos Costa de Morais, R.M. Marine Polysaccharides from Algae with Potential Biomedical Applications. Mar. Drugs 2015, 13, 2967–3028. [Google Scholar] [CrossRef]
- Senthilkumar, K.; Manivasagan, P.; Venkatesan, J.; Kim, S.-K. Brown seaweed fucoidan: Biological activity and apoptosis, growth signaling mechanism in cancer. Int. J. Biol. Macromol. 2013, 60, 366–374. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.-I.; Kim, H.-J. Preparation of low molecular weight fucoidan by gamma-irradiation and its anticancer activity. Carbohydr. Polym. 2013, 97, 358–362. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.; Mikkelsen, M.D.; Tran, V.H.N.; Trang, V.T.D.; Rhein-Knudsen, N.; Holck, J.; Rasin, A.B.; Cao, H.T.T.; Van, T.T.T.; Meyer, A.S. Enzyme-Assisted Fucoidan Extraction from Brown Macroalgae Fucus distichus subsp. evanescens and Saccharina latissima. Mar. Drugs 2020, 18, 296. [Google Scholar] [CrossRef]
- Saravana, P.S.; Tilahun, A.; Gerenew, C.; Tri, V.D.; Kim, N.H.; Kim, G.-D.; Woo, H.-C.; Chun, B.-S. Subcritical water extraction of fucoidan from Saccharina japonica: Optimization, characterization and biological studies. J. Appl. Phycol. 2017, 30, 579–590. [Google Scholar] [CrossRef]
- Yuan, Y.; Macquarrie, D. Microwave assisted extraction of sulfated polysaccharides (fucoidan) from Ascophyllum nodosum and its antioxidant activity. Carbohydr. Polym. 2015, 129, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Flórez-Fernández, N.; López-García, M.; González-Muñoz, M.J.; Vilariño, J.M.L.; Domínguez, H. Ultrasound-assisted extraction of fucoidan from Sargassum muticum. J. Appl. Phycol. 2017, 29, 1553–1561. [Google Scholar] [CrossRef]
- Hammed, A.M.; Jaswir, I.; Simsek, S.; Alam, Z.; Amid, A. Enzyme Aided Extraction of Sulfated Polysaccharides from Turbinaria turbinata Brown Seaweed. Int. Food Res. J. 2017, 24, 1660–1666. [Google Scholar]
- Wijesinghe, W.; Athukorala, Y.; Jeon, Y.-J. Effect of anticoagulative sulfated polysaccharide purified from enzyme-assistant extract of a brown seaweed Ecklonia cava on Wistar rats. Carbohydr. Polym. 2011, 86, 917–921. [Google Scholar] [CrossRef]
- Alboofetileh, M.; Rezaei, M.; Tabarsa, M. Enzyme-assisted extraction of Nizamuddinia zanardinii for the recovery of sulfated polysaccharides with anticancer and immune-enhancing activities. J. Appl. Phycol. 2018, 31, 1391–1402. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, H.; Chen, L.; Mi, Z.; Xu, Y.; Zhao, G.; Liu, S.; Lei, H.; Wang, Z.; Niu, J. Extraction, purification, and determination of the gastroprotective activity of glucomannan from Bletilla striata. Carbohydr. Polym. 2020, 246, 116620. [Google Scholar] [CrossRef]
- Huang, D.; Zhang, M.; Yi, P.; Yan, C. Structural characterization and osteoprotective effects of a novel oligo-glucomannan obtained from the rhizome of Cibotium barometz by alkali extraction. Ind. Crop. Prod. 2018, 113, 202–209. [Google Scholar] [CrossRef]
- Kumoro, A.; Yuganta, T.; Retnowati, D.; Ratnawati, R. Acid Hydrolysis and Ethanol Precipitation for Glucomannan Extraction from Crude Porang (Amorphophallus Oncophyllus) Tuber Flour. Chem. Chem. Technol. 2018, 12, 101–108. [Google Scholar] [CrossRef]
- Kou, S.; Peters, L.M.; Mucalo, M.R. Chitosan: A review of sources and preparation methods. Int. J. Biol. Macromol. 2020, 169, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Machado, D.I.; López-Cervantes, J.; Correa-Murrieta, M.A.; Sánchez-Duarte, R.G.; Cruz-Flores, P.; la Mora-López, G.S. Chitosan. In Nonvitamin and Nonmineral Nutritional Supplements; Elsevier: Amsterdam, The Netherlands, 2019; pp. 485–493. [Google Scholar] [CrossRef]
- Rhein-Knudsen, N.; Ale, M.T.; Meyer, A.S. Seaweed Hydrocolloid Production: An Update on Enzyme Assisted Extraction and Modification Technologies. Mar. Drugs 2015, 13, 3340–3359. [Google Scholar] [CrossRef]
- Raus, R.A.; Nawawi, W.M.F.W.; Nasaruddin, R.R. Alginate and alginate composites for biomedical applications. Asian J. Pharm. Sci. 2020, 16, 280–306. [Google Scholar] [CrossRef]
- Rhein-Knudsen, N.; Ale, M.T.; Ajalloueian, F.; Meyer, A.S. Characterization of alginates from Ghanaian brown seaweeds: Sargassum spp. and Padina spp. Food Hydrocoll. 2017, 71, 236–244. [Google Scholar] [CrossRef]
- Athukorala, Y.; Jung, W.-K.; Vasanthan, T.; Jeon, Y.-J. An anticoagulative polysaccharide from an enzymatic hydrolysate of Ecklonia cava. Carbohydr. Polym. 2006, 66, 184–191. [Google Scholar] [CrossRef]
- Hardouin, K.; Burlot, A.-S.; Umami, A.; Tanniou, A.; Stiger-Pouvreau, V.; Widowati, I.; Bedoux, G.; Bourgougnon, N. Biochemical and antiviral activities of enzymatic hydrolysates from different invasive French seaweeds. J. Appl. Phycol. 2013, 26, 1029–1042. [Google Scholar] [CrossRef]
- Jeeva, P.; Doss, S.S.; Sundaram, V.; Jayaraman, G. Production of controlled molecular weight hyaluronic acid by glucostat strategy using recombinant Lactococcus lactis cultures. Appl. Microbiol. Biotechnol. 2019, 103, 4363–4375. [Google Scholar] [CrossRef]
- Jeeva, P.; Jayaprakash, S.R.; Jayaraman, G. Hyaluronic acid production is enhanced by harnessing the heme-induced respiration in recombinant Lactococcus lactis cultures. Biochem. Eng. J. 2022, 182, 108428. [Google Scholar] [CrossRef]
- Rodriguez-Marquez, C.D.; Arteaga-Marin, S.; Rivas-Sánchez, A.; Autrique-Hernández, R.; Castro-Muñoz, R. A Review on Current Strategies for Extraction and Purification of Hyaluronic Acid. Int. J. Mol. Sci. 2022, 23, 6038. [Google Scholar] [CrossRef] [PubMed]
- He, F. Laemmli-SDS-PAGE. Bio-Protocol 2011, 1, e80. [Google Scholar] [CrossRef]
- BeMiller, J.N. Carrageenans. In Carbohydrate Chemistry for Food Scientists; Elsevier: Amsterdam, The Netherlands, 2019; pp. 279–291. [Google Scholar] [CrossRef]
- Tarman, K.; Sadi, U.; Santoso, J.; Hardjito, L. Carrageenan and its Enzymatic Extraction. Encycl. Mar. Biotechnol. 2020, 1, 147–159. [Google Scholar] [CrossRef]
- Guo, Z.; Wei, Y.; Zhang, Y.; Xu, Y.; Zheng, L.; Zhu, B.; Yao, Z. Carrageenan oligosaccharides: A comprehensive review of preparation, isolation, purification, structure, biological activities and applications. Algal Res. 2021, 61, 102593. [Google Scholar] [CrossRef]
- Liu, D.; Ouyang, Y.; Chen, R.; Wang, M.; Ai, C.; El-Seedi, H.R.; Sarker, M.R.; Chen, X.; Zhao, C. Nutraceutical potentials of algal ulvan for healthy aging. Int. J. Biol. Macromol. 2021, 194, 422–434. [Google Scholar] [CrossRef] [PubMed]
- Kidgell, J.T.; Magnusson, M.; de Nys, R.; Glasson, C.R.K. Ulvan: A systematic review of extraction, composition and function. Algal Res. 2019, 39, 101422. [Google Scholar] [CrossRef]
- Zhang, W.; Oda, T.; Yu, Q.; Jin, J.-O. Fucoidan from Macrocystis pyrifera Has Powerful Immune-Modulatory Effects Compared to Three Other Fucoidans. Mar. Drugs 2015, 13, 1084–1104. [Google Scholar] [CrossRef]
- Zhou, S.; Liu, X.; Guo, Y.; Wang, Q.; Peng, D.; Cao, L. Comparison of the immunological activities of arabinoxylans from wheat bran with alkali and xylanase-aided extraction. Carbohydr. Polym. 2010, 81, 784–789. [Google Scholar] [CrossRef]
- Martin, S.; Burton, D.; Roitt, I.; Delves, P. Roitt’s Essential Immunology; Wiley: Hoboken, NJ, USA, 2016. [Google Scholar]
- van Kasteren, S.I.; Neefjes, J.; Ovaa, H. Creating molecules that modulate immune responses. Nat. Rev. Chem. 2018, 2, 184–193. [Google Scholar] [CrossRef]
- Netea, M.G.; Domínguez-Andrés, J.; Barreiro, L.B.; Chavakis, T.; Divangahi, M.; Fuchs, E.; Joosten, L.A.B.; Van Der Meer, J.W.M.; Mhlanga, M.M.; Mulder, W.J.M.; et al. Defining trained immunity and its role in health and disease. Nat. Rev. Immunol. 2020, 20, 375–388. [Google Scholar] [CrossRef]
- Zindel, J.; Kubes, P. DAMPs, PAMPs, and LAMPs in Immunity and Sterile Inflammation. Annu. Rev. Pathol. Mech. Dis. 2020, 15, 493–518. [Google Scholar] [CrossRef] [PubMed]
- Placha, D.; Jampilek, J. Chronic Inflammatory Diseases, Anti-Inflammatory Agents and Their Delivery Nanosystems. Pharmaceutics 2021, 13, 64. [Google Scholar] [CrossRef]
- Schett, G.; McInnes, I.B.; Neurath, M.F. Reframing Immune-Mediated Inflammatory Diseases through Signature Cytokine Hubs. New Engl. J. Med. 2021, 385, 628–639. [Google Scholar] [CrossRef]
- Rezoagli, E.; Fumagalli, R.; Bellani, G. Definition and epidemiology of acute respiratory distress syndrome. Ann. Transl. Med. 2017, 5, 282. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, A.S.; Alonso, M.J.; de la Fuente, M. Nanoengineering of vaccines using natural polysaccharides. Biotechnol. Adv. 2015, 33, 1279–1293. [Google Scholar] [CrossRef] [PubMed]
- Shariatinia, Z.; Jalali, A.M. Chitosan-based hydrogels: Preparation, properties and applications. Int. J. Biol. Macromol. 2018, 115, 194–220. [Google Scholar] [CrossRef]
- Baraya, Y.S.; Wong, K.K.; Yaacob, N.S. The Immunomodulatory Potential of Selected Bioactive Plant-Based Compounds in Breast Cancer: A Review. Anti-Cancer Agents Med. Chem. 2017, 17, 770–783. [Google Scholar] [CrossRef]
- Stickney, D.R.; Foon, K.A. Biologic response modifiers. Curr. Opin. Oncol. 1992, 4, 847–855. [Google Scholar] [CrossRef]
- Murphy, E.J.; Rezoagli, E.; Pogue, R.; Simonassi-Paiva, B.; Abidin, I.I.Z.; Fehrenbach, G.W.; O’Neil, E.; Major, I.; Laffey, J.G.; Rowan, N. Immunomodulatory activity of β-glucan polysaccharides isolated from different species of mushroom—A potential treatment for inflammatory lung conditions. Sci. Total. Environ. 2021, 809, 152177. [Google Scholar] [CrossRef]
- Cao, S.; He, J.; Zhang, F.; Tian, H.; Liu, C.; Wang, H.; Zhou, Q. Baeomyces lotiformis sp. nov. from China. Mycotaxon 2018, 132, 831–837. [Google Scholar] [CrossRef]
- Teng, L.; Fu, H.; Deng, C.; Chen, J.; Chen, J. Modulating the SDF-1/CXCL12-induced cancer cell growth and adhesion by sulfated K5 polysaccharides in vitro. Biomed. Pharmacother. 2015, 73, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Feng, Z.; Yuan, F.; Guo, J.; Suo, S.; Wang, B. Identification and functional analysis of the autofluorescent substance in Limonium bicolor salt glands. Plant Physiol. Biochem. 2015, 97, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ma, L.; Zhang, L. Molecular basis for Poria cocos mushroom polysaccharide used as an antitumor drug in China. Prog. Mol. Biol. Transl. Sci. 2019, 163, 263–296. [Google Scholar] [CrossRef]
- Cordero, R.J.B.; Frases, S.; Guimaräes, A.J.; Rivera, J.; Casadevall, A. Evidence for branching in cryptococcal capsular polysaccharides and consequences on its biological activity. Mol. Microbiol. 2011, 79, 1101–1117. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Lorenzo, C.; Blanco-Fernandez, B.; Puga, A.M.; Concheiro, A. Crosslinked ionic polysaccharides for stimuli-sensitive drug delivery. Adv. Drug Deliv. Rev. 2013, 65, 1148–1171. [Google Scholar] [CrossRef] [PubMed]
- Arca, H.Ç.; Günbeyaz, M.; Şenel, S. Chitosan-based systems for the delivery of vaccine antigens. Expert Rev. Vaccines 2009, 8, 937–953. [Google Scholar] [CrossRef] [PubMed]
- Suginta, W.; Khunkaewla, P.; Schulte, A. Electrochemical Biosensor Applications of Polysaccharides Chitin and Chitosan. Chem. Rev. 2013, 113, 5458–5479. [Google Scholar] [CrossRef]
- Mizrahy, S.; Peer, D. Polysaccharides as building blocks for nanotherapeutics. Chem. Soc. Rev. 2011, 41, 2623–2640. [Google Scholar] [CrossRef]
- Akhtar, M.; Ding, R. Covalently cross-linked proteins & polysaccharides: Formation, characterisation and potential applications. Curr. Opin. Colloid Interface Sci. 2017, 28, 31–36. [Google Scholar] [CrossRef]
- Lo, T.C.T.; Kang, M.W.; Wang, B.C.; Chang, C.A. Glycosyl linkage characteristics and classifications of exo-polysaccharides of some regionally different strains of Lentinula edodes by amplified fragment length polymorphism assay and cluster analysis. Anal. Chim. Acta 2007, 592, 146–153. [Google Scholar] [CrossRef]
- Kralovec, J.; Metera, K.; Kumar, J.; Watson, L.; Girouard, G.; Guan, Y.; Carr, R.; Barrow, C.; Ewart, H. Immunostimulatory principles from Chlorella pyrenoidosa—Part 1: Isolation and biological assessment in vitro. Phytomedicine 2007, 14, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Li, J.; Liu, Y.; Wu, D.; Cai, P.; Pan, Y. Structural characterization and immunomodulatory activity of a water soluble polysaccharide isolated from Botrychium ternatum. Carbohydr. Polym. 2017, 171, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Florean, C.; Dicato, M.; Diederich, M. Immune-modulating and anti-inflammatory marine compounds against cancer. Semin. Cancer Biol. 2022, 80, 58–72. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.-O.; Yu, Q. Fucoidan delays apoptosis and induces pro-inflammatory cytokine production in human neutrophils. Int. J. Biol. Macromol. 2015, 73, 65–71. [Google Scholar] [CrossRef]
- Jin, J.-O.; Zhang, W.; Du, J.-Y.; Wong, K.-W.; Oda, T.; Yu, Q. Fucoidan Can Function as an Adjuvant In Vivo to Enhance Dendritic Cell Maturation and Function and Promote Antigen-Specific T Cell Immune Responses. PLoS ONE 2014, 9, e99396. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Teruya, K.; Yoshida, T.; Eto, H.; Shirahata, S. Fucoidan Extract Enhances the Anti-Cancer Activity of Chemotherapeutic Agents in MDA-MB-231 and MCF-7 Breast Cancer Cells. Mar. Drugs 2013, 11, 81–98. [Google Scholar] [CrossRef]
- Takahashi, H.; Kawaguchi, M.; Kitamura, K.; Narumiya, S.; Kawamura, M.; Tengan, I.; Nishimoto, S.; Hanamure, Y.; Majima, Y.; Tsubura, S.; et al. An Exploratory Study on the Anti-inflammatory Effects of Fucoidan in Relation to Quality of Life in Advanced Cancer Patients. Integr. Cancer Ther. 2017, 17, 282–291. [Google Scholar] [CrossRef]
- Peng, Y.; Song, Y.; Wang, Q.; Hu, Y.; He, Y.; Ren, D.; Wu, L.; Liu, S.; Cong, H.; Zhou, H. In vitro and in vivo immunomodulatory effects of fucoidan compound agents. Int. J. Biol. Macromol. 2018, 127, 48–56. [Google Scholar] [CrossRef]
- Ale, M.T.; Maruyama, H.; Tamauchi, H.; Mikkelsen, J.D.; Meyer, A.S. Fucoidan from Sargassum sp. and Fucus vesiculosus reduces cell viability of lung carcinoma and melanoma cells in vitro and activates natural killer cells in mice in vivo. Int. J. Biol. Macromol. 2011, 49, 331–336. [Google Scholar] [CrossRef]
- Mami, S.; Yeganeh, F.; Farahani, E.; Anissian, A.; Hoseini, M.H.M. Chitin Micro Particles Regulate Splenocytes Immune Response in Experimental Autoimmune Encephalomyelitis. Iran. J. Allergy, Asthma Immunol. 2019, 18, 190–199. [Google Scholar] [CrossRef]
- Mami, S.; Yeganeh, F.; Salari, A.-A.; Anissian, A.; Azizi, M.; Hajimollahoseini, M. Oral chitin treatment improved demyelination in murine autoimmune encephalomyelitis model by inhibition of inflammatory responses. Int. Immunopharmacol. 2020, 84, 106536. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Wen, Z.-S.; Xiang, X.-W.; Huang, Y.-N.; Gao, Y.; Qu, Y.-L. Immunostimulative Activity of Low Molecular Weight Chitosans in RAW264.7 Macrophages. Mar. Drugs 2015, 13, 6210–6225. [Google Scholar] [CrossRef] [PubMed]
- Azizi, M.; Yousefi, R.; Yeganeh, F.; Mami, S.; Hoseini, M.H.M. Co-administration of chitin micro-particle and Leishmania antigen proposed a new immune adjuvant against experimental leishmaniasis. Parasite Immunol. 2019, 41, e12676. [Google Scholar] [CrossRef] [PubMed]
- Bi, D.; Zhou, R.; Cai, N.; Lai, Q.; Han, Q.; Peng, Y.; Jiang, Z.; Tang, Z.; Lu, J.; Bao, W.; et al. Alginate enhances Toll-like receptor 4-mediated phagocytosis by murine RAW264.7 macrophages. Int. J. Biol. Macromol. 2017, 105, 1446–1454. [Google Scholar] [CrossRef]
- Zha, Z.; Lv, Y.; Tang, H.; Li, T.; Miao, Y.; Cheng, J.; Wang, G.; Tan, Y.; Zhu, Y.; Xing, X.; et al. An orally administered butyrate-releasing xylan derivative reduces inflammation in dextran sulphate sodium-induced murine colitis. Int. J. Biol. Macromol. 2020, 156, 1217–1233. [Google Scholar] [CrossRef]
- Luo, M.; Shao, B.; Nie, W.; Wei, X.-W.; Li, Y.-L.; Wang, B.-L.; He, Z.-Y.; Liang, X.; Ye, T.-H.; Wei, Y.-Q. Antitumor and Adjuvant Activity of λ-carrageenan by Stimulating Immune Response in Cancer Immunotherapy. Sci. Rep. 2015, 5, srep11062. [Google Scholar] [CrossRef]
- Harikrishnan, R.; Devi, G.; Van Doan, H.; Balasundaram, C.; Arockiaraj, J.; Jagruthi, C. Efficacy of ulvan on immune response and immuno-antioxidant gene modulation in Labeo rohita against columnaris disease. Fish Shellfish. Immunol. 2021, 117, 262–273. [Google Scholar] [CrossRef]
- Jin, L.; Xu, K.; Liang, Y.; Du, P.; Wan, S.; Jiang, C. Effect of hyaluronic acid on cytokines and immune cells change in patients of knee osteoarthritis. BMC Musculoskelet. Disord. 2022, 23, 1–9. [Google Scholar] [CrossRef]
- Lee, B.M.; Park, S.J.; Noh, I.; Kim, C.-H. The effects of the molecular weights of hyaluronic acid on the immune responses. Biomater. Res. 2021, 25, 1–13. [Google Scholar] [CrossRef]
- Dai, J.; Chen, J.; Qi, J.; Ding, M.; Liu, W.; Shao, T.; Han, J.; Wang, G. Konjac Glucomannan from Amorphophallus konjac enhances immunocompetence of the cyclophosphamide-induced immunosuppressed mice. Food Sci. Nutr. 2020, 9, 728–735. [Google Scholar] [CrossRef]
- He, T.-B.; Huang, Y.-P.; Yang, L.; Liu, T.-T.; Gong, W.-Y.; Wang, X.-J.; Sheng, J.; Hu, J.-M. Structural characterization and immunomodulating activity of polysaccharide from Dendrobium officinale. Int. J. Biol. Macromol. 2016, 83, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-P.; He, T.-B.; Cuan, X.-D.; Wang, X.-J.; Hu, J.-M.; Sheng, J. 1,4-β-d-Glucomannan from Dendrobium officinale Activates NF-κB via TLR4 to Regulate the Immune Response. Molecules 2018, 23, 2658. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, K.; Ogushi, M.; Nakashima, A.; Nakano, Y.; Suzuki, K. Accelerated Wound Healing on the Skin Using a Film Dressing with β-Glucan Paramylon. In Vivo 2018, 32, 799–805. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, A.; Suzuki, K.; Asayama, Y.; Konno, M.; Saito, K.; Yamazaki, N.; Takimoto, H. Oral administration of Euglena gracilis Z and its carbohydrate storage substance provides survival protection against influenza virus infection in mice. Biochem. Biophys. Res. Commun. 2017, 494, 379–383. [Google Scholar] [CrossRef]
- Wang, Q.; Sheng, X.; Shi, A.; Hu, H.; Yang, Y.; Liu, L.; Fei, L.; Liu, H. β-Glucans: Relationships between Modification, Conformation and Functional Activities. Molecules 2017, 22, 257. [Google Scholar] [CrossRef]
- Han, B.; Baruah, K.; Cox, E.; Vanrompay, D.; Bossier, P. Structure-Functional Activity Relationship of β-Glucans From the Perspective of Immunomodulation: A Mini-Review. Front. Immunol. 2020, 11, 658. [Google Scholar] [CrossRef]
- Kaur, R.; Sharma, M.; Ji, D.; Xu, M.; Agyei, D. Structural Features, Modification, and Functionalities of Beta-Glucan. Fibers 2019, 8, 1. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, N.; Xiong, Z.-Q.; Wang, G.; Xia, Y.; Lai, P.; Ai, L. Structural characterization and rheological properties of β-D-glucan from hull-less barley (Hordeum vulgare L. var. nudum Hook. f.). Phytochemistry 2018, 155, 155–163. [Google Scholar] [CrossRef]
- Friedman, M. Mushroom Polysaccharides: Chemistry and Antiobesity, Antidiabetes, Anticancer, and Antibiotic Properties in Cells, Rodents, and Humans. Foods 2016, 5, 80. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Cai, L.; Yang, Q.; Luo, Z.; Liang, L.; Liang, Y.; Wu, B.; Ding, L.; Zhang, D.; Xu, X.; et al. Anti-leukemia activities of selenium nanoparticles embedded in nanotube consisted of triple-helix β-d-glucan. Carbohydr. Polym. 2020, 240, 116329. [Google Scholar] [CrossRef]
- Lyu, F.; Xu, X.; Zhang, L. Natural polysaccharides with different conformations: Extraction, structure and anti-tumor activity. J. Mater. Chem. B 2020, 8, 9652–9667. [Google Scholar] [CrossRef]
- Zheng, X.; Lu, F.; Xu, X.; Zhang, L. Extended chain conformation of β-glucan and its effect on antitumor activity. J. Mater. Chem. B 2017, 5, 5623–5631. [Google Scholar] [CrossRef]
- Rieder, A.; Grimmer, S.; Kolset, S.O.; Michaelsen, T.E.; Knutsen, S.H. Cereal β-glucan preparations of different weight average molecular weights induce variable cytokine secretion in human intestinal epithelial cell lines. Food Chem. 2011, 128, 1037–1043. [Google Scholar] [CrossRef]
- Talbott, S.M.; Talbott, J.A. Baker’s Yeast Beta-Glucan Supplement Reduces Upper Respiratory Symptoms and Improves Mood State in Stressed Women. J. Am. Coll. Nutr. 2012, 31, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Bekkering, S.; Blok, B.A.; Joosten, L.A.B.; Riksen, N.P.; van Crevel, R.; Netea, M.G. In Vitro Experimental Model of Trained Innate Immunity in Human Primary Monocytes. Clin. Vaccine Immunol. 2016, 23, 926–933. [Google Scholar] [CrossRef] [PubMed]
- Saeed, S.; Quintin, J.; Kerstens, H.H.D.; Rao, N.A.; Aghajanirefah, A.; Matarese, F.; Cheng, S.-C.; Ratter, J.; Berentsen, K.; van der Ent, M.A.; et al. Epigenetic programming of monocyte-to-macrophage differentiation and trained innate immunity. Science 2014, 345, 1251086. [Google Scholar] [CrossRef] [PubMed]
- Tanioka, A.; Tanabe, K.; Hosono, A.; Kawakami, H.; Kaminogawa, S.; Tsubaki, K.; Hachimura, S. Enhancement of Intestinal Immune Function in Mice by β-D-Glucan from Aureobasidium Pullulans ADK-34. Scand. J. Immunol. 2013, 78, 61–68. [Google Scholar] [CrossRef]
- Stothers, C.L.; Burelbach, K.R.; Owen, A.M.; Patil, N.K.; McBride, M.A.; Bohannon, J.K.; Luan, L.; Hernandez, A.; Patil, T.K.; Williams, D.L.; et al. β-Glucan Induces Distinct and Protective Innate Immune Memory in Differentiated Macrophages. J. Immunol. 2021, 207, 2785–2798. [Google Scholar] [CrossRef]
- Evans, M.; Falcone, P.H.; Crowley, D.C.; Sulley, A.M.; Campbell, M.; Zakaria, N.; Lasrado, J.A.; Fritz, E.P.; Herrlinger, K.A. Effect of a Euglena gracilis Fermentate on Immune Function in Healthy, Active Adults: A Randomized, Double-Blind, Placebo-Controlled Trial. Nutrients 2019, 11, 2926. [Google Scholar] [CrossRef]
- Sugiyama, A.; Hata, S.; Suzuki, K.; Yoshida, E.; Nakano, R.; Mitra, S.; Arashida, R.; Asayama, Y.; Yabuta, Y.; Takeuchi, T. Oral Administration of Paramylon, a.BETA.-1,3-D-Glucan Isolated from Euglena gracilis Z Inhibits Development of Atopic Dermatitis-Like Skin Lesions in NC/Nga Mice. J. Veter- Med Sci. 2010, 72, 755–763. [Google Scholar] [CrossRef]
- Sonck, E.; Stuyven, E.; Goddeeris, B.; Cox, E. The effect of β-glucans on porcine leukocytes. Veter- Immunol. Immunopathol. 2010, 135, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Russo, R.; Barsanti, L.; Evangelista, V.; Frassanito, A.M.; Longo, V.; Pucci, L.; Penno, G.; Gualtieri, P. Euglena gracilis paramylon activates human lymphocytes by upregulating pro-inflammatory factors. Food Sci. Nutr. 2016, 5, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Murphy, E.J.; Masterson, C.; Rezoagli, E.; O’Toole, D.; Laffey, J.G.; Major, I.; Stack, G.; Rowan, N. Immunomodulation Properties of a Novel β-Glucan Extract from the Mushroom Lentinus edodes in an In-Vitro Lung Injury Model. In A60. LUNG INJURY, SEPSIS, AND ARDS; American Thoracic Society: New York, NY, USA, 2019; p. A2114. [Google Scholar] [CrossRef]
- Murphy, E.J.; Masterson, C.; Rezoagli, E.; O’Toole, D.; Major, I.; Stack, G.D.; Lynch, M.; Laffey, J.G.; Rowan, N.J. β-Glucan extracts from the same edible shiitake mushroom Lentinus edodes produce differential in-vitro immunomodulatory and pulmonary cytoprotective effects—Implications for coronavirus disease (COVID-19) immunotherapies. Sci. Total. Environ. 2020, 732, 139330. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Mendieta, S.; Guillén, D.; Hernández-Pando, R.; Sánchez, S.; Rodríguez-Sanoja, R. Potential of glucans as vaccine adjuvants: A review of the α-glucans case. Carbohydr. Polym. 2017, 165, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Kylin, H. Zur Biochemie Der Meeresalgen. Hoppe Seylers Z Physiol. Chem. 1913, 83, 171–197. [Google Scholar] [CrossRef]
- I Bilan, M.; A Grachev, A.; E Ustuzhanina, N.; Shashkov, A.S.; E Nifantiev, N.; I Usov, A. Structure of a fucoidan from the brown seaweed Fucus evanescens C.Ag. Carbohydr. Res. 2002, 337, 719–730. [Google Scholar] [CrossRef]
- Deniaud-Bouët, E.; Kervarec, N.; Michel, G.; Tonon, T.; Kloareg, B.; Hervé, C. Chemical and enzymatic fractionation of cell walls from Fucales: Insights into the structure of the extracellular matrix of brown algae. Ann. Bot. 2014, 114, 1203–1216. [Google Scholar] [CrossRef]
- Shang, F.; Mou, R.; Zhang, Z.; Gao, N.; Lin, L.; Li, Z.; Wu, M.; Zhao, J. Structural analysis and anticoagulant activities of three highly regular fucan sulfates as novel intrinsic factor Xase inhibitors. Carbohydr. Polym. 2018, 195, 257–266. [Google Scholar] [CrossRef]
- Li, C.; Niu, Q.; Li, S.; Zhang, X.; Liu, C.; Cai, C.; Li, G.; Yu, G. Fucoidan from sea cucumber Holothuria polii: Structural elucidation and stimulation of hematopoietic activity. Int. J. Biol. Macromol. 2019, 154, 1123–1131. [Google Scholar] [CrossRef]
- El Rashed, Z.; Lupidi, G.; Kanaan, H.; Grasselli, E.; Canesi, L.; Khalifeh, H.; Demori, I. Antioxidant and Antisteatotic Activities of a New Fucoidan Extracted from Ferula hermonis Roots Harvested on Lebanese Mountains. Molecules 2021, 26, 1161. [Google Scholar] [CrossRef]
- Haddad, M.; Zein, S.; Shahrour, H.; Hamadeh, K.; Karaki, N.; Kanaan, H. Antioxidant activity of water-soluble polysaccharide extracted from Eucalyptus cultivated in Lebanon. Asian Pac. J. Trop. Biomed. 2017, 7, 157–160. [Google Scholar] [CrossRef]
- Wu, S.-Y.; Yan, M.-D.; Wu, A.T.; Yuan, K.S.-P.; Liu, S.H. Brown Seaweed Fucoidan Inhibits Cancer Progression by Dual Regulation of mir-29c/ADAM12 and miR-17-5p/PTEN Axes in Human Breast Cancer Cells. J. Cancer 2016, 7, 2408–2419. [Google Scholar] [CrossRef] [PubMed]
- Shannon, E.; Abu-Ghannam, N. Antibacterial Derivatives of Marine Algae: An Overview of Pharmacological Mechanisms and Applications. Mar. Drugs 2016, 14, 81. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Q.; Zhang, Z.; Hou, Y.; Zhang, H. In-vitro anticoagulant activity of fucoidan derivatives from brown seaweed Laminaria japonica. Chin. J. Oceanol. Limnol. 2011, 29, 679–685. [Google Scholar] [CrossRef]
- Ahmadi, A.; Moghadamtousi, S.Z.; Abubakar, S.; Zandi, K. Antiviral potential of algae polysaccharides isolated from marine sources: A Review. BioMed Res. Int. 2015, 2015, 825203. [Google Scholar] [CrossRef]
- Ale, M.T.; Mikkelsen, J.D.; Meyer, A.S. Important Determinants for Fucoidan Bioactivity: A Critical Review of Structure-Function Relations and Extraction Methods for Fucose-Containing Sulfated Polysaccharides from Brown Seaweeds. Mar. Drugs 2011, 9, 2106–2130. [Google Scholar] [CrossRef] [PubMed]
- Apostolova, E.; Lukova, P.; Baldzhieva, A.; Katsarov, P.; Nikolova, M.; Iliev, I.; Peychev, L.; Trica, B.; Oancea, F.; Delattre, C.; et al. Immunomodulatory and Anti-Inflammatory Effects of Fucoidan: A Review. Polymers 2020, 12, 2338. [Google Scholar] [CrossRef]
- Kasai, A.; Arafuka, S.; Koshiba, N.; Takahashi, D.; Toshima, K. Systematic synthesis of low-molecular weight fucoidan derivatives and their effect on cancer cells. Org. Biomol. Chem. 2015, 13, 10556–10568. [Google Scholar] [CrossRef]
- Zhang, W.; Du, J.-Y.; Jiang, Z.; Okimura, T.; Oda, T.; Yu, Q.; Jin, J.-O. Ascophyllan Purified from Ascophyllum nodosum Induces Th1 and Tc1 Immune Responses by Promoting Dendritic Cell Maturation. Mar. Drugs 2014, 12, 4148–4164. [Google Scholar] [CrossRef]
- Teruya, T.; Tatemoto, H.; Konishi, T.; Tako, M. Structural characteristics and in vitro macrophage activation of acetyl fucoidan from Cladosiphon okamuranus. Glycoconj. J. 2009, 26, 1019–1028. [Google Scholar] [CrossRef]
- Xue, M.; Liang, H.; Tang, Q.; Xue, C.; He, X.; Zhang, L.; Zhang, Z.; Liang, Z.; Bian, K.; Zhang, L.; et al. The Protective and Immunomodulatory Effects of Fucoidan Against 7,12-Dimethyl benz[a]anthracene-Induced Experimental Mammary Carcinogenesis Through the PD1/PDL1 Signaling Pathway in Rats. Nutr. Cancer 2017, 69, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.-Y.; Lin, T.-Y.; Wu, Y.-C.; Tsao, S.-M.; Hwang, P.-A.; Shih, Y.-W.; Hsu, J. Fucoidan inhibition of lung cancer in vivo and in vitro: Role of the Smurf2-dependent ubiquitin proteasome pathway in TGFβ receptor degradation. Oncotarget 2014, 5, 7870–7885. [Google Scholar] [CrossRef] [PubMed]
- Coussens, L.M.; Zitvogel, L.; Palucka, A.K. Neutralizing Tumor-Promoting Chronic Inflammation: A Magic Bullet? Science 2013, 339, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, J.V.; Cobucci, R.; Jatobá, C.A.N.; Fernandes, T.; De Azevedo, J.W.V.; De Araújo, J.M.G. The Role of the Mediators of Inflammation in Cancer Development. Pathol. Oncol. Res. 2015, 21, 527–534. [Google Scholar] [CrossRef]
- Tsai, H.-L.; Tai, C.-J.; Huang, C.-W.; Chang, F.-R.; Wang, J.-Y. Efficacy of Low-Molecular-Weight Fucoidan as a Supplemental Therapy in Metastatic Colorectal Cancer Patients: A Double-Blind Randomized Controlled Trial. Mar. Drugs 2017, 15, 122. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, J.; Li, Q.; Wang, Y.; Wang, C. Two natural glucomannan polymers, from Konjac and Bletilla, as bioactive materials for pharmaceutical applications. Biotechnol. Lett. 2014, 37, 1–8. [Google Scholar] [CrossRef]
- Wu, W.-T.; Cheng, H.-C.; Chen, H.-L. Ameliorative effects of konjac glucomannan on human faecal β-glucuronidase activity, secondary bile acid levels and faecal water toxicity towards Caco-2 cells. Br. J. Nutr. 2010, 105, 593–600. [Google Scholar] [CrossRef]
- Harmayani, E.; Aprilia, V.; Marsono, Y. Characterization of glucomannan from Amorphophallus oncophyllus and its prebiotic activity in vivo. Carbohydr. Polym. 2014, 112, 475–479. [Google Scholar] [CrossRef]
- Vassilaros, S.; Tsibanis, A.; Tsikkinis, A.; A Pietersz, G.; McKenzie, I.F.; Apostolopoulos, V. Up to 15-year clinical follow-up of a pilot Phase III immunotherapy study in stage II breast cancer patients using oxidized mannan–MUC1. Immunotherapy 2013, 5, 1177–1182. [Google Scholar] [CrossRef]
- Shariatinia, Z. Pharmaceutical applications of chitosan. Adv. Colloid Interface Sci. 2018, 263, 131–194. [Google Scholar] [CrossRef]
- Nishimura, K.; Nishi, N.; Saiki, I.; Tokura, S.; Azuma, I. Immunological activity of chitin and its derivatives. Vaccine 1984, 2, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Shibata, Y.; Metzger, W.J.; Myrvik, Q.N. Chitin particle-induced cell-mediated immunity is inhibited by soluble mannan: Mannose receptor-mediated phagocytosis initiates IL-12 production. J. Immunol. 1997, 159, 2462–2467. [Google Scholar] [CrossRef] [PubMed]
- Carroll, E.C.; Jin, L.; Mori, A.; Muñoz-Wolf, N.; Oleszycka, E.; Moran, H.B.; Mansouri, S.; McEntee, C.; Lambe, E.; Agger, E.M.; et al. The Vaccine Adjuvant Chitosan Promotes Cellular Immunity via DNA Sensor cGAS-STING-Dependent Induction of Type I Interferons. Immunity 2016, 44, 597–608. [Google Scholar] [CrossRef]
- Bueter, C.L.; Lee, C.K.; Wang, J.P.; Ostroff, G.R.; Specht, C.A.; Levitz, S.M. Spectrum and Mechanisms of Inflammasome Activation by Chitosan. J. Immunol. 2014, 192, 5943–5951. [Google Scholar] [CrossRef] [PubMed]
- Komi, D.E.A.; Sharma, L.; Cruz, C.S.D. Chitin and Its Effects on Inflammatory and Immune Responses. Clin. Rev. Allergy Immunol. 2017, 54, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Bennacef, C.; Desobry-Banon, S.; Probst, L.; Desobry, S. Advances on alginate use for spherification to encapsulate biomolecules. Food Hydrocoll. 2021, 118, 106782. [Google Scholar] [CrossRef]
- Zhang, J.; Qi, Y.; Shen, Y.; Li, H. Research Progress on Chemical Modification and Application of Cellulose: A Review. Mater. Sci. 2022, 28, 60–67. [Google Scholar] [CrossRef]
- Hecht, H.; Srebnik, S. Structural Characterization of Sodium Alginate and Calcium Alginate. Biomacromolecules 2016, 17, 2160–2167. [Google Scholar] [CrossRef]
- Yang, J.-S.; Xie, Y.-J.; He, W. Research progress on chemical modification of alginate: A review. Carbohydr. Polym. 2011, 84, 33–39. [Google Scholar] [CrossRef]
- Tønnesen, H.H.; Karlsen, J. Alginate in Drug Delivery Systems. Drug Dev. Ind. Pharm. 2002, 28, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Yudiati, E.; Isnansetyo, A.; Murwantoko; Triyanto; Handayani, C.R. Alginate from Sargassum siliquosum Simultaneously Stimulates Innate Immunity, Upregulates Immune Genes, and Enhances Resistance of Pacific White Shrimp (Litopenaeus vannamei) Against White Spot Syndrome Virus (WSSV). Mar. Biotechnol. 2019, 21, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Afraei, S.; Azizi, G.; Zargar, S.J.; Sedaghat, R.; Mirshafiey, A. New therapeutic approach by G2013 in experimental model of multiple sclerosis. Acta Neurol. Belg. 2014, 115, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Mirshafiey, A.; Mortazavi-Jahromi, S.S.; Taeb, M.; Cuzzocrea, S.; Esposito, E. Evaluation of the Effect of α-L-Guluronic Acid (G2013) on COX-1, COX-2 Activity and Gene Expression for Introducing this Drug as a Novel NSAID with Immunomodulatory Property. Recent Patents Inflamm. Allergy Drug Discov. 2018, 12, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Mortazavi-Jahromi, S.S.; Jamshidi, M.M.; Yousefi, M.; Navabi, S.S.; Motamed, N.; Zavareh, F.T.; Mirshafiey, A. Inhibitory effect of G2013 molecule as a novel immunomodulatory agent, on miR-155 gene expression in HEK-Blue hTLR4 cell line. Eur. J. Inflamm. 2016, 14, 86–92. [Google Scholar] [CrossRef]
- Nazeri, S.; Azarian, S.K.; Fattahi, M.J.; Sedaghat, R.; Zavareh, F.T.; Aghazadeh, Z.; Abdollahi, M.; Mirshafiey, A. Preclinical and pharmacotoxicology evaluation of α-l-guluronic acid (G2013) as a non-steroidal anti-inflammatory drug with immunomodulatory property. Immunopharmacol. Immunotoxicol. 2017, 39, 59–65. [Google Scholar] [CrossRef]
- Ibragimova, D.S.; Davletova, X.S.; Khayitbayev, A.K.; Qizi, S.M.A. Extracting Of Hyaluronic Acid from Different Sources. Eurasian J. Phys. Chem. Math. 2022, 3, 50–54. [Google Scholar]
- Mohammed, M.; Devnarain, N.; Elhassan, E.; Govender, T. Exploring the applications of hyaluronic acid-based nanoparticles for diagnosis and treatment of bacterial infections. WIREs Nanomed. Nanobiotechnology 2022, 14, e1799. [Google Scholar] [CrossRef]
- Samadi, M.; Shahraky, M.K.; Tabandeh, F.; Aminzadeh, S.; Dina, M. Enhanced hyaluronic acid production in Streptococcus zooepidemicus by an optimized culture medium containing hyaluronidase inhibitor. Prep. Biochem. Biotechnol. 2021, 52, 413–423. [Google Scholar] [CrossRef]
- Thanh, T.N.; Laowattanatham, N.; Ratanavaraporn, J.; Sereemaspun, A.; Yodmuang, S. Hyaluronic acid crosslinked with alginate hydrogel: A versatile and biocompatible bioink platform for tissue engineering. Eur. Polym. J. 2022, 166, 111027. [Google Scholar] [CrossRef]
- Zhang, D.; Ren, Y.; He, Y.; Chang, R.; Guo, S.; Ma, S.; Guan, F.; Yao, M. In situ forming and biocompatible hyaluronic acid hydrogel with reactive oxygen species-scavenging activity to improve traumatic brain injury repair by suppressing oxidative stress and neuroinflammation. Mater. Today Bio 2022, 15, 100278. [Google Scholar] [CrossRef]
- Bowman, S.; Awad, M.E.; Hamrick, M.W.; Hunter, M.; Fulzele, S. Recent advances in hyaluronic acid based therapy for osteoarthritis. Clin. Transl. Med. 2018, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- Huynh, A.; Priefer, R. Hyaluronic acid applications in ophthalmology, rheumatology, and dermatology. Carbohydr. Res. 2020, 489, 107950. [Google Scholar] [CrossRef] [PubMed]
- Snetkov, P.; Zakharova, K.; Morozkina, S.; Olekhnovich, R.; Uspenskaya, M. Hyaluronic Acid: The Influence of Molecular Weight on Structural, Physical, Physico-Chemical, and Degradable Properties of Biopolymer. Polymers 2020, 12, 1800. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.; Bahadur, P. Modified hyaluronic acid based materials for biomedical applications. Int. J. Biol. Macromol. 2018, 121, 556–571. [Google Scholar] [CrossRef] [PubMed]
- Trombino, S.; Servidio, C.; Curcio, F.; Cassano, R. Strategies for Hyaluronic Acid-Based Hydrogel Design in Drug Delivery. Pharmaceutics 2019, 11, 407. [Google Scholar] [CrossRef]
- Smith, P.; Ziolek, R.M.; Gazzarrini, E.; Owen, D.M.; Lorenz, C.D. On the interaction of hyaluronic acid with synovial fluid lipid membranes. Phys. Chem. Chem. Phys. 2019, 21, 9845–9857. [Google Scholar] [CrossRef]
- Fang, R.; Jiang, Q.; Guan, Y.; Gao, P.; Zhang, R.; Zhao, Z.; Jiang, Z. Golgi apparatus-synthesized sulfated glycosaminoglycans mediate polymerization and activation of the cGAMP sensor STING. Immunity 2021, 54, 962–975.e8. [Google Scholar] [CrossRef]
- Mohan, N.; Tadi, S.R.R.; Pavan, S.S.; Sivaprakasam, S. Deciphering the role of dissolved oxygen and N-acetyl glucosamine in governing higher molecular weight hyaluronic acid synthesis in Streptococcus zooepidemicus cell factory. Appl. Microbiol. Biotechnol. 2020, 104, 3349–3365. [Google Scholar] [CrossRef]
- Kisukeda, T.; Onaya, J.; Yoshioka, K. Effect of diclofenac etalhyaluronate (SI-613) on the production of high molecular weight sodium hyaluronate in human synoviocytes. BMC Musculoskelet. Disord. 2019, 20, 1–9. [Google Scholar] [CrossRef]
- Zamboni, F.; Vieira, S.; Reis, R.L.; Oliveira, J.M.; Collins, M.N. The potential of hyaluronic acid in immunoprotection and immunomodulation: Chemistry, processing and function. Prog. Mater. Sci. 2018, 97, 97–122. [Google Scholar] [CrossRef]
- Unfer, V.; Tilotta, M.; Kaya, C.; Noventa, M.; Török, P.; Alkatout, I.; Gitas, G.; Bilotta, G.; Laganà, A.S. Absorption, distribution, metabolism and excretion of hyaluronic acid during pregnancy: A matter of molecular weight. Expert Opin. Drug Metab. Toxicol. 2021, 17, 823–840. [Google Scholar] [CrossRef] [PubMed]
- Shebl, R.I.; Amer, M.E.; Abuamara, T.M.M.; Matar, E.R.; Ahmed, H.F.; Gomah, T.A.; El Moselhy, L.E.; Abu-Elghait, M.; Mohamed, A.F. Staphylococcus aureus derived hyaluronic acid and bacillus Calmette-Guérin purified proteins as immune enhancers to rabies vaccine and related immuno-histopathological alterations. Clin. Exp. Vaccine Res. 2021, 10, 229–239. [Google Scholar] [CrossRef]
- Galdi, F.; Pedone, C.; McGee, C.A.; George, M.; Rice, A.B.; Hussain, S.S.; Vijaykumar, K.; Boitet, E.R.; Tearney, G.J.; McGrath, J.A.; et al. Inhaled high molecular weight hyaluronan ameliorates respiratory failure in acute COPD exacerbation: A pilot study. Respir. Res. 2021, 22, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Pacheco-Quito, E.-M.; Ruiz-Caro, R.; Veiga, M.-D. Carrageenan: Drug Delivery Systems and Other Biomedical Applications. Mar. Drugs 2020, 18, 583. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.-I.; Zhang, G.; Li, X.; Leung, C.-H.; Ma, D.-L.; Dong, L.; Wang, C. Carrageenan activates monocytes via type-specific binding with interleukin-8: An implication for design of immuno-active biomaterials. Biomater. Sci. 2017, 5, 403–407. [Google Scholar] [CrossRef]
- Yermak, I.M.; Volod’ko, A.V.; Khasina, E.I.; Davydova, V.N.; Chusovitin, E.A.; Goroshko, D.L.; Kravchenko, A.O.; Solov’eva, T.F.; Maleev, V.V. Inhibitory Effects of Carrageenans on Endotoxin-Induced Inflammation. Mar. Drugs 2020, 18, 248. [Google Scholar] [CrossRef]
- Kalitnik, A.A.; Karetin, Y.A.; Kravchenko, A.O.; Khasina, E.I.; Yermak, I.M. Influence of carrageenan on cytokine production and cellular activity of mouse peritoneal macrophages and its effect on experimental endotoxemia. J. Biomed. Mater. Res. Part A 2017, 105, 1549–1557. [Google Scholar] [CrossRef]
- Kuroyama, H.; Tsumuraya, Y. A xylosyltransferase that synthesizes β-(1→4)-xylans in wheat (Triticum aestivum L.) seedlings. Planta 2001, 213, 231–240. [Google Scholar] [CrossRef]
- Kabel, M.A.; Borne, H.V.D.; Vincken, J.-P.; Voragen, A.G.; Schols, H.A. Structural differences of xylans affect their interaction with cellulose. Carbohydr. Polym. 2007, 69, 94–105. [Google Scholar] [CrossRef]
- Hsieh, Y.S.; Harris, P.J. Xylans of Red and Green Algae: What Is Known about Their Structures and How They Are Synthesised? Polymers 2019, 11, 354. [Google Scholar] [CrossRef]
- Bush, J.R.; Liang, H.; Dickinson, M.; Botchwey, E.A. Xylan hemicellulose improves chitosan hydrogel for bone tissue regeneration. Polym. Adv. Technol. 2016, 27, 1050–1055. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.T.; Duarte, M.E.; Holanda, D.M.; Kim, S.W. Friend or Foe? Impacts of Dietary Xylans, Xylooligosaccharides, and Xylanases on Intestinal Health and Growth Performance of Monogastric Animals. Animals 2021, 11, 609. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhan, A.; Ye, Y.; Liu, C.; Hang, F.; Li, K.; Li, J. Molecular modification, structural characterization, and biological activity of xylans. Carbohydr. Polym. 2021, 269, 118248. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, S.S.; Passos, C.P.; Madureira, P.; Vilanova, M.; Coimbra, M.A. Structure–function relationships of immunostimulatory polysaccharides: A review. Carbohydr. Polym. 2015, 132, 378–396. [Google Scholar] [CrossRef]
- Ebringerová, A.; Kardošová, A.; Hromádková, Z.; Malovíková, A.; Hříbalová, V. Immunomodulatory activity of acidic xylans in relation to their structural and molecular properties. Int. J. Biol. Macromol. 2002, 30, 1–6. [Google Scholar] [CrossRef]
- Akhtar, M.; Tariq, A.F.; Awais, M.M.; Iqbal, Z.; Muhammad, F.; Shahid, M.; Hiszczynska-Sawicka, E. Studies on wheat bran Arabinoxylan for its immunostimulatory and protective effects against avian coccidiosis. Carbohydr. Polym. 2012, 90, 333–339. [Google Scholar] [CrossRef]
- Kardošová, A.; Malovíková, A.; Pätoprstý, V.; Nosál’Ová, G.; Matáková, T. Structural characterization and antitussive activity of a glucuronoxylan from Mahonia aquifolium (Pursh) Nutt. Carbohydr. Polym. 2002, 47, 27–33. [Google Scholar] [CrossRef]
- Tabarsa, M.; You, S.; Dabaghian, E.H.; Surayot, U. Water-soluble polysaccharides from Ulva intestinalis: Molecular properties, structural elucidation and immunomodulatory activities. J. Food Drug Anal. 2018, 26, 599–608. [Google Scholar] [CrossRef]
- Thanh, T.T.T.; Quach, T.M.T.; Nguyen, T.N.; Luong, D.V.; Bui, M.L.; Van Tran, T.T. Structure and cytotoxic activity of ulvan extracted from green seaweed Ulva lactuca. Int. J. Biol. Macromol. 2016, 93, 695–702. [Google Scholar] [CrossRef]
- Berri, M.; Olivier, M.; Holbert, S.; Dupont, J.; Demais, H.; Le Goff, M.; Collen, P.N. Ulvan from Ulva armoricana (Chlorophyta) activates the PI3K/Akt signalling pathway via TLR4 to induce intestinal cytokine production. Algal Res. 2017, 28, 39–47. [Google Scholar] [CrossRef]
- Fernández-Díaz, C.; Coste, O.; Malta, E.-J. Polymer chitosan nanoparticles functionalized with Ulva ohnoi extracts boost in vitro ulvan immunostimulant effect in Solea senegalensis macrophages. Algal Res. 2017, 26, 135–142. [Google Scholar] [CrossRef]







| Polysaccharide | Species | Extraction Method (EM) and Purification (PFN) | Ref. |
| Beta-glucan (C6H10O5)n Up to 3000 kDa | Lentinus edodes Mushroom | EM A water-extracted residue was used to extract the alkaline-soluble crude PSR. The supernatant was then neutralized, dialyzed, precipitated (1:4 v/v) in EtOH, and freeze-dried. PFN Extract diluted in 15 mL of DW and loaded onto a Q-Sepharose Fast Flow strong anion-exchange column (QFF, 4.6 15 cm). At a flow rate of 4 mL/min, the PSR was eluted with 600 mL of DTW and 0.2 and 0.4 mol/L NaCl solutions, sequentially. | [62] |
| Saccharomyces cerevisiae | EM Yeast cultured in extract–glucose broth for 48 h, heated for 2 h at 80 °C, centrifuged, and then rinsed with DTW. Samples centrifuged and a five-fold CH3COOH extraction was used to extract β-glucan from lysed yeast pellets. | [63] | |
| Xylans (C5H8O4)n Up to 200 kDa | Wheat straw and corn stalks | EM Hot alkali extraction. Raw materials were extracted in sealed with NaOH, and optimal conditions were 100 °C, 72 min, and 7% NaOH (wheat straw); 120 °C, 118 min, and 9% NaOH (corn stalks). | [64] |
| Wheat straw | EM Cold alkaline extraction and enzymatic hydrolysis. Wheat straw was incubated with NaOH for 90 min at 40 °C. Solid fraction washed with DTW and dried. Commercial enzyme Ultraflo L, Shearzyme 500 L, and Pentopan mono conc. were tested. Straw was sterilized, and 1 g was incubated at 230 rpm for 54 h, with 200 U of endo-xylanase and 42 mL of buffer. | [65] | |
| Hyaluronic acid (C14H21NO11)n Up to 20,000 kDa | Gallus gallus | EM Frozen rooster combs were ground and incubated in acetone for 2 h at 4 °C. Acetone was drained, and dried combs were incubated in 5% CH3COON for 2 h and 10 times. EtOH was added, and the solution purified in chloroform 4 times and then chloroform–amyl alcohol. The solution was dialyzed and precipitated with EtOH. PFN Chloroform and chloroform–amyl alcohol (1:2) were used in the extraction to remove impurities, such as proteins. | [66] |
| Recombinant Corynebacterium glutamicum | EM Recombinant C. glutamicum was cultured in fermenter containing 5 g/L of (NH4)2SO4, 5 g/L of urea, 1 g/L of K2HPO4, 1 g/L of KH2PO4, 250 mg/L of MgSO4, and 10 mg/L of CaCl2. The culture was harvested and purified to isolate the hyaluronic acid. PFN First purification with EtOH:C3H8O (1:1, 1:2, and 1:3) at 4 °C or −20 °C and then centrifuged (4000 rpm, 30 min). Pellets dissolved in 3% SDA, and concentration of HA was calculated. (1) Impurities were removed in three steps consisting of SDA/charcoal/centrifugation; (2) TCA 100% was added to the SDA and incubated in an ice bath for 30 min and centrifuged (16,000 rpm, 30 min); (3) solution was dissolved in chloroform–butanol and stirred on a shaker for 30 min. The aqueous phase was collected. | [67] | |
| Chitosan (C6H11NO4)n Up to 1000 kDa | Penaeus semisulcatus | EM Dry shells demineralized in 1N HCl at 30 °C for 6 h and deproteinized in 3.5% NaOH at 65 °C for 2 h. Then, shells were decolorized in 0.315% NaCl and obtained chitin deacetylated in 50% NaOH at 100 °C for 5 h to obtain chitosan. | [68] |
| Hermetia illucens larvae, pupal exuviae and dead adults | EM Powdered samples were demineralized in 0.5 M formic acid at RT for 1 h, filtered, washed, dried, and then incubated in 2 M NaOH at 80 °C for 2 h for deproteinization. Unbleached and bleached chitin fractions were resuspended in 12 M NaOH and stirred at 100 °C for 4 h, followed by incubation in 1% (v/v) CH3COOH at RT for 48 h. Chitosan was precipitated in pH 8. PFN Realized after deproteinization. Bleached chitin was obtained after treating the samples with 5% H2O2 at 90 °C for 30–60 min. High-purity chitin was obtained (84–86.8%), resulting in higher yields of chitosan. | [69] | |
| Carrageenan (C24H36O25S2)n Up to 2000 kDa | Hypnea musciformis | EM Conventional and ultrasonic-assisted extraction (UAE). Sun-dried algae was hydrated with water and kept at RT for 12 h, followed by depigmentation using methanol:acetone (1:1). The depigmented extract was treated with 3% KOH at 80 °C for 4 h and filtered/washed. Retained was redissolved in water, incubated at 90 °C for 4 h, and then filtered. Extract was precipitated with 95% ethanol, dried, and milled. UAE was performed as conventional until depigmentation then sonicated at 400 and 500 W for 10 and 20 min in an ice bath. | [70] |
| Eucheuma cottonii | EM Two extraction methods using Ca(OH)2 and a second with NaOH were performed. Dry algae were mixed with Ca(OH)2 and incubated at 80–110 °C for 2–12 h and filtered; CO2 was then injected into extract until pH reached 7–8, twice, and injected again until solution became clear. Finally, 5% KCl was added, and the solution was filtered, dried, and crushed. In the NaOH method, dry algae were incubated in 10% NaOH/12% KCl at 65 °C for 4 h and washed 3 times. Then, algae were incubated in water for 4 h and boiled at 100 °C for 4 h. After that, 5% KCl solution was used to precipitate carrageenan. | [71] | |
| Ulvan RMO (C5H9O4)n XYL (C6H10O7) n UA (C5H8O6) n Up to 200 kDa | Cotesia glomerata and Ulva flexuosa | EM Dry algae extracted by Soxhlet for 7 h with DCM/acetone (1:1). The residual was subjected to a hot extraction with water in a water bath for 7 h at 75–85 °C. Extract was filtrated and liquid concentrated for enzymatic hydrolysis with α-amylase at 20 °C for 30 min. Then, extract was treated with proteinase K at 37 °C for 24 h, and the reaction stopped by heating at 80 °C for 10 min. Ethanol was added and incubated for 24 h in a fridge to precipitate ulvan. PFN Washed consecutively with ethanol, acetone, and diethyl ether. | [72] |
| Ulva armoricana | EM Enzyme-assisted extraction. Crushed algae were incubated with water (1:1) and a neutral endo-protease, a mix of neutral and alkaline endo-proteases, a multiple mix of carbohydrates, a mix of endo-1,4-β-xylanase/endo-1,3(4)-β-glucanase, cellulase, and exo-β1,3(4)-glucanase enzymes (6% weight/dw, w/dw) at 50 °C for 3 h, followed by denaturation at 90 °C for 15 min. Samples were filtered and then freeze-dried. | [73] | |
| Alginate MA (C6H8O6)n GA (C6H8O7)n | Sargassum vulgare | EM Dry algae were soaked in 2% formaldehyde for 24 h, washed with water, and left for 24 h in 0.2M HCl. Second wash with water and extraction with 2% Na2CO3 at 60 °C for 5 h and precipitation of alginate with ethanol. PFN Re-precipitation with EtOH and acetone. | [74] |
| Sargassum baccularia, Sargassum binderi, Sargassum siliquosum and Turbinaria conoides | EM Both a cold and hot extraction were performed. In the cold extraction, algae were left in 1% CaCl2 at RT for 18 h, washed with DTW, and incubated with 3% Na2CO3 for 1 h and left for 18 h. Mixture was separated by centrifugation and alginate extracted by adding EtOH/water (1:1). Precipitate was collected and dried. In the hot extraction, algae were treated similarly as in cold method, except the incubation time and temperature were 3 h and 50 °C. PFN Washed with EtOH and dried in air, followed by drying in a vacuum oven. | [75] | |
| Fucoidan (C6H10O5S)n Up to 1000 kDa | Sargassum wightii | EM Hot-water extraction and ultrasonication-assisted extraction. | [76] |
| Ecklonia maxima, Laminaria pallida and Splachnidium rugosum | EM 3 methods using a hot-water extraction (24 h at 70 °C), acid extraction (0.15 M HCl at 65 °C for 2 h), and salt extraction (incubated with methanol–chloroform overnight, RT) were performed. | [77] | |
| Glucomannan GLU (C6H10O5)n MAN (C6H12O6)n | Dendrobium officinale | EM Solid–liquid extraction. Petroleum ether (70 °C/2 h), 80% EtOH (90 °C/2 h), and DTW (100 °C/2 h). PFN Precipitated with EtOH, centrifuged, resuspended in distilled water and deproteinized 5 times (Sevag reagent), washed with ethyl acetate/acetone and EtOH. | [78] |
| Dendrobium devonianum | EM Reflux system with methanol for 1 h at 65 °C and microwave-assisted extraction (800 W/80 °C/9 min), precipitation with 95% EtOH, and redissolved in water (60 °C). PFN Ultra-centrifugal filtration. | [79] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murphy, E.J.; Fehrenbach, G.W.; Abidin, I.Z.; Buckley, C.; Montgomery, T.; Pogue, R.; Murray, P.; Major, I.; Rezoagli, E. Polysaccharides—Naturally Occurring Immune Modulators. Polymers 2023, 15, 2373. https://doi.org/10.3390/polym15102373
Murphy EJ, Fehrenbach GW, Abidin IZ, Buckley C, Montgomery T, Pogue R, Murray P, Major I, Rezoagli E. Polysaccharides—Naturally Occurring Immune Modulators. Polymers. 2023; 15(10):2373. https://doi.org/10.3390/polym15102373
Chicago/Turabian StyleMurphy, Emma J., Gustavo Waltzer Fehrenbach, Ismin Zainol Abidin, Ciara Buckley, Therese Montgomery, Robert Pogue, Patrick Murray, Ian Major, and Emanuele Rezoagli. 2023. "Polysaccharides—Naturally Occurring Immune Modulators" Polymers 15, no. 10: 2373. https://doi.org/10.3390/polym15102373
APA StyleMurphy, E. J., Fehrenbach, G. W., Abidin, I. Z., Buckley, C., Montgomery, T., Pogue, R., Murray, P., Major, I., & Rezoagli, E. (2023). Polysaccharides—Naturally Occurring Immune Modulators. Polymers, 15(10), 2373. https://doi.org/10.3390/polym15102373