
Bone Marrow Mesenchymal Stromal Cells on Silk Fibroin Scaffolds to Attenuate Polymicrobial Sepsis Induced by Cecal Ligation and Puncture




Abstract

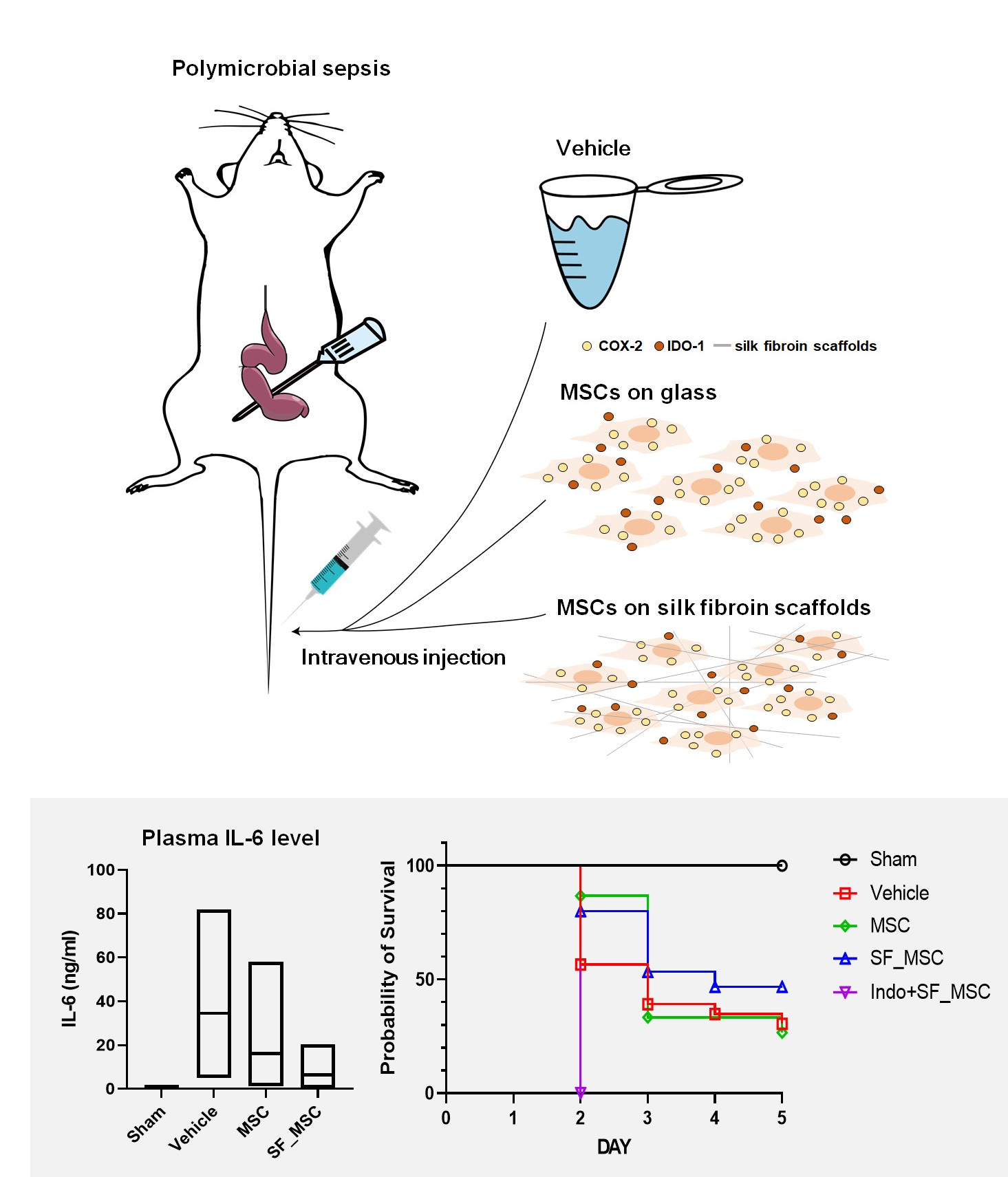{kind=link}
{kind=link}
{kind=link}
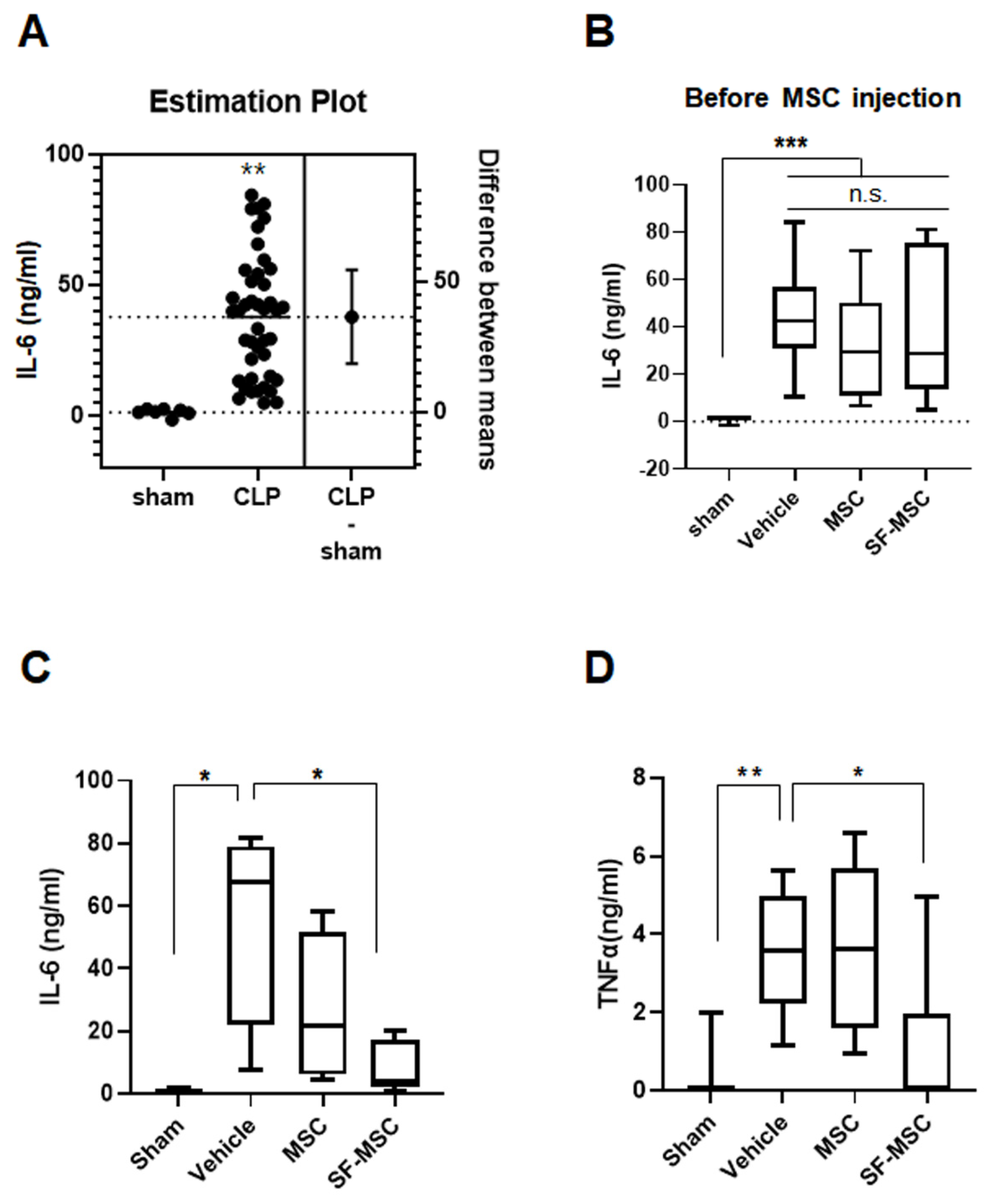{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Preparation of Silk Solution
2.2. Fabrication and Characterization of SF Nanofibers
2.3. MSC Preparation
2.4. Animal Surgery
2.5. Measurement of Prostaglandin E2 (PGE2) Production
2.6. Determination of Serum Cytokine Levels
2.7. Hematoxylin–Eosin (H&E) Staining Assay
2.8. RNA Extraction and Quantitative Real-Time PCR
2.9. Western Blotting
2.10. Survival Studies
2.11. Statistical Analyses
3. Results
3.1. Characterization of the SF Nanofiber Scaffolds
3.2. MSCs on the SF Nanofibers Reduce the IL-6 Level Efficiently in Septic Mice
3.3. Abrogation of Cecal Ligation and Puncture Induced Inflammation by MSCs on SF Nanofibers
3.4. MSCs on SF Nanofibers Produce More PGE2
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.D.; Coopersmith, C.M.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 2016, 315, 801–810. [Google Scholar] [CrossRef]
- Ayala, A.; Venet, M.P.F.; Lomas-Neira, J.; Swan, R.; Chung, C.S. Apoptosis in Sepsis: Mechanisms, Clinical Impact and Potential Therapeutic Targets. Curr. Pharm. Des. 2008, 14, 1852–1859. [Google Scholar] [CrossRef]
- Dombrovskiy, V.Y.; Martin, A.A.; Sunderram, J.; Paz, H.L. Rapid Increase in Hospitalization and Mortality Rates for Severe Sepsis in the United States: A Trend Analysis from 1993 to 2003. Crit. Care Med. 2007, 35, 1244–1250. [Google Scholar] [CrossRef]
- Melamed, A.; Sorvillo, F.J. The Burden of Sepsis-Associated Mortality in the United States from 1999 to 2005: An Analysis of Multiple-Cause-of-Death Data. Crit. Care 2009, 13, R28. [Google Scholar] [CrossRef]
- Ho, M.S.; Mei, S.H.; Stewart, D.J. The Immunomodulatory and Therapeutic Effects of Mesenchymal Stromal Cells for Acute Lung Injury and Sepsis. J. Cell Physiol. 2015, 230, 2606–2617. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Kim, W.Y. Mesenchymal Stromal Cell Application as an Emerging Translational Medicine for Acute Respiratory Distress Syndrome. Ann. Tranl. Med. 2020, 8, 267. [Google Scholar] [CrossRef]
- Wu, J.; Wang, Y.; Li, L. Functional Significance of Exosomes Applied in Sepsis: A Novel Approach to Therapy. Biochim. Biophys. Acta 2017, 1863, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Chen, W.; Huang, S.; Yao, G.; Tang, X.; Sun, L. Mesenchymal Stem Cells Prevent Overwhelming Inflammation and Reduce Infection Severity Via Recruiting Cxcr3+ Regulatory T Cells. Clin. Transl. Immunol. 2020, 9, e1181. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, K.; Mayer, B.; Mezey, E. Modulation of Bone Marrow Stromal Cell Functions in Infectious Disease by Toll-Like Receptor Ligands. J. Mol. Med. 2010, 88, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal Criteria for Defining Multipotent Mesenchymal Stromal Cells. The International Society for Cellular Therapy Position Statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Kobabu, S.; Lowery, J.W.; Jimi, E. Cell Fate and Differentiation of Bone Marrow Mesenchymal Stem Cells. Stem. Cell Int. 2016, 2016, 3753581. [Google Scholar]
- Diaz, M.F.; Vaidya, A.B.; Evans, S.M.; Lee, H.J.; Aertker, B.M.; Alexander, A.J.; Price, K.M.; Ozuna, J.A.; Liao, G.P.; Aroom, K.R.; et al. Biomechanical Forces Promote Immune Regulatory Function of Bone Marrow Mesenchymal Stromal Cells. Stem. Cells 2017, 35, 1259–1272. [Google Scholar] [CrossRef]
- Kim, O.H.; Yoon, O.J.; Lee, H.J. Silk Fibroin Scaffolds Potentiate Immunomodulatory Function of Human Mesenchymal Stromal Cells. Biochem. Biophys. Res. Commun. 2019, 519, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, S.; Kundu, S.C. Silk Protein-Based Hydrogels: Promising Advanced Materials for Biomedical Applications. Acta Biomater. 2016, 31, 17–32. [Google Scholar] [CrossRef]
- Sapru, S.; Das, S.; Mandal, M.; Ghosh, A.K.; Kundu, S.C. Prospects of Nonmulberry Silk Protein Sericin-Based Nanofibrous Matrices for Wound Healing—In Vitro and in Vivo Investigations. Acta Biomater. 2018, 78, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Aronoff, D.M. Cyclooxygenase Inhibition in Sepsis: Is There Life after Death? Mediat. Inflamm. 2012, 2012, 696897. [Google Scholar] [CrossRef] [PubMed]
- Khatri, M.; Hussain, N.; Ghazali, S.E.; Yamamoto, T.; Kobayashi, S.; Khatri, Z.; Ahmed, F.; Kim, I.S. Ultrasonic Assisted Dyeing of Silk Fbroin Nanofbers: An Energy Efcient Coloration at Room Temperature. Appl. Nanosci. 2020, 10, 917–930. [Google Scholar] [CrossRef]
- Shao, J.; Liu, J.; Zheng, J.; Carr, C.M. X-Ray Photoelectron Spectroscopic Study of Silk Fibroin Surface. Polym. Int. 2002, 51, 1479–1483. [Google Scholar] [CrossRef]
- Nasim, A.; Mahdi, N.; Mohammad, H.K. Structural Characterization and Mechanical Properties of Electrospun Silk Fibroin Nanofiber Mats. Polym. Sci. 2010, 51, 407–412. [Google Scholar]
- Leila, S.; Mahdi, N. Electrospun Silk Fibroin Nanofibers with Improved Surface Texture. J. Text. Polym. 2016, 4, 75–82. [Google Scholar]
- Meisel, R.; Zibert, A.; Laryea, M.; Göbel, U.; Däubener, W.; Dilloo, D. Human Bone Marrow Stromal Cells Inhibit Allogeneic T-Cell Responses by Indoleamine 2,3-Dioxygenase-Mediated Tryptophan Degradation. Blood 2004, 103, 4619–4621. [Google Scholar] [CrossRef] [PubMed]
- Spaggiari, G.M.; Capobianco, A.; Abdelrazik, H.; Becchetti, F.; Mingari, M.C.; Moretta, L. Mesenchymal Stem Cells Inhibit Natural Killer-Cell Proliferation, Cytotoxicity, and Cytokine Production: Role of Indoleamine 2,3-Dioxygenase and Prostaglandin E2. Blood 2008, 111, 1327–1333. [Google Scholar] [CrossRef]
- Wang, C.; Chen, J.; Sun, L.; Liu, Y. Tgf-Beta Signaling-Dependent Alleviation of Dextran Sulfate Sodium-Induced Colitis by Mesenchymal Stem Cell Transplantation. Mol. Biol. Rep. 2014, 41, 4977–4983. [Google Scholar] [CrossRef]
- Liu, Y.; Ren, H.; Wang, J.; Yang, F.; Li, J.; Zhou, Y.; Yuan, X.; Zhu, W.; Shi, X. Prostaglandin E2 Secreted by Mesenchymal Stem Cells Protects against Acute Liver Failure Via Enhancing Hepatocyte Proliferation. FASEB J. 2018, 33, 2514–2525. [Google Scholar] [CrossRef] [PubMed]
- Lever, A.; Mackenzie, I. Sepsis: Definition, Epidemiology, and Diagnosis. BMJ 2007, 335, 879–883. [Google Scholar] [CrossRef]
- McIntyre, L.A.; Stewart, D.J.; Mei, S.H.J.; Courtman, D.; Watpool, I.; Granton, J.; Marshall, J.; dos Santos, C.; Walley, K.R.; Winston, B.W.; et al. Cellular Immunotherapy for Septic Shock. A Phase I Clinical Trial. Am. J. Respir. Crit. Care Med. 2018, 197, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.Y.; Ding, X.F.; Liang, H.Y.; Zhang, X.Y.; Liu, S.H.; Han, B.; Duan, X.G.; Sun, T.W. Efficacy of Mesenchymal Stem Cell Therapy for Sepsis: A Meta-Analysis of Preclinical Studies. Stem. Cell Res. Ther. 2020, 11, 214. [Google Scholar] [CrossRef]
- Oberholzer, A.; Oberholzer, C.; Moldawer, L.L. Sepsis Syndromes: Understanding the Role of Innate and Acquired Immunity. Shock 2001, 16, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Ebong, S.J.; Call, D.R.; Bolgos, G.; Newcomb, D.E.; Granger, J.I.; O’Reilly, M.; Remick, D.G. Immunopathologic Alterations in Murine Models of Sepsis of Increasing Severity. Infect. Immun. 1999, 7, 6603–6610. [Google Scholar] [CrossRef]
- Remick, D.G.; Bolgos, G.R.; Siddiqui, J.; Shin, J.; Nemzek, J.A. Six at Six: Interleulin-6 Measured 6 H after the Initiation of Sepsis Predicts Mortality over 3 Days. Shock 2002, 17, 463–467. [Google Scholar] [CrossRef]
- Park, K.S.; Svennerholm, K.; Shelke, G.V.; Bandeira, E.; Lässer, C.; Jang, S.C.; Chandode, R.; Gribonika, I.; Lötvall, J. Mesenchymal Stromal Cell-Derived Nanovesicles Ameliorate Bacterial Outer Membrane Vesicle-Induced Sepsis Via Il-10. Stem Cell Res. Therapy 2019, 10, 231. [Google Scholar] [CrossRef]
- Hack, C.E.; De Groot, E.R.; Felt-Bersma, R.J.; Nuijens, J.H.; Strack Van Schijndel, R.J.; Eerenberg-Belmer, A.J.; Thijs, L.G.; Aarden, L.A. Increased Plasma Levels of Interleukin-6 in Sepsis. Blood 1989, 74, 1704–1710. [Google Scholar] [CrossRef] [PubMed]
- Riedemann, N.C.; Neff, T.A.; Guo, R.F.; Bernacki, K.D.; Laudes, I.J.; Sarma, J.V.; Lambris, J.D.; Ward, P.A. Rotective Effects of Il-6 Blockade in Sepsis Are Linked to Reduced C5a Receptor Expression. J. Immunol. 2003, 170, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Tipnis, S.; Viswanathan, C.; Majumdar, A.S. Immunosuppressive Properties of Human Umbilical Cord-Derived Mesenchymal Stem Cells: Role of B7-H1 and Ido. Immunol. Cell Biol. 2010, 88, 795–806. [Google Scholar] [CrossRef] [PubMed]
- Saparov, A.; Ogay, V.; Nurgozhin, T.; Jumabay, M.; Chen, W.C.W. Preconditioning of Human Mesenchymal Stem Cells to Enhance Their Regulation of the Immune Response. Stem. Cell Int. 2016, 2016, 3924858. [Google Scholar] [CrossRef]
- Eggenhofer, E.; Benseler, V.; Kroemer, A.; Popp, F.C.; Geissler, E.K.; Schlitt, H.J.; Baan, C.C.; Dahlke, M.H.; Hoogduijn, M.J. Mesenchymal Stem Cells Are Short-Lived and Do Not Migrate Beyond the Lungs after Intravenous Infusions. Front. Immunol. 2012, 3, 297. [Google Scholar] [CrossRef] [PubMed]
- Cao, B.; Peng, Y.; Liu, X.; Ding, J. Effects of Functional Groups of Materials on Nonspecific Adhesion and Chondrogenic Induction of Mesenchymal Stem Cells on Free and Micropatterned Surfaces. ACS Appl. Mater. Interfaces 2017, 9, 23574–23585. [Google Scholar] [CrossRef]
- Noronha, N.C.; Mizukami, A.; Caliári-Oliveira, C.; Cominal, J.G.; Rocha, J.L.M.; Covas, D.T.; Swiech, K.; Malmegrim, K.C.R. Priming Approaches to Improve the Efficacy of Mesenchymal Stromal Cell-Based Therapies. Stem Cell Res. Ther. 2019, 10, 131. [Google Scholar] [CrossRef]
- Caliari, S.R.; Vega, S.L.; Kwon, M.; Soulas, E.M.; Burdick, J.A. Dimensionality and Spreading Influence Msc Yap/Taz Signaling in Hydrogel Environments. Biomaterials 2016, 103, 314–323. [Google Scholar] [CrossRef]
- Černochová, P.; Blahová, L.; Medalová, J.; Nečas, N.; Michlíček, M.; Kaushik, P.; Přibyl, J.; Bartošíková, J.; Manakhov, A.; Bačáková, L.; et al. Cell Type Specific Adhesion to Surfaces Functionalised by Amine Plasma Polymers. Sci. Rep. 2020, 10, 9357. [Google Scholar] [CrossRef]
- Bacakova, L.; Filova, E.; Parizek, M.; Ruml, T.; Svorcik, V. Modulation of Cell Adhesion, Proliferation and Differentiation on Materials Designed for Body Implants. Biotechnol. Adv. 2011, 29, 739–767. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, O.-H.; Park, J.-H.; Son, J.-I.; Yoon, O.-J.; Lee, H.-J. Bone Marrow Mesenchymal Stromal Cells on Silk Fibroin Scaffolds to Attenuate Polymicrobial Sepsis Induced by Cecal Ligation and Puncture. Polymers 2021, 13, 1433. https://doi.org/10.3390/polym13091433
Kim O-H, Park J-H, Son J-I, Yoon O-J, Lee H-J. Bone Marrow Mesenchymal Stromal Cells on Silk Fibroin Scaffolds to Attenuate Polymicrobial Sepsis Induced by Cecal Ligation and Puncture. Polymers. 2021; 13(9):1433. https://doi.org/10.3390/polym13091433
Chicago/Turabian StyleKim, Ok-Hyeon, Jun-Hyung Park, Jong-In Son, Ok-Ja Yoon, and Hyun-Jung Lee. 2021. "Bone Marrow Mesenchymal Stromal Cells on Silk Fibroin Scaffolds to Attenuate Polymicrobial Sepsis Induced by Cecal Ligation and Puncture" Polymers 13, no. 9: 1433. https://doi.org/10.3390/polym13091433
APA StyleKim, O.-H., Park, J.-H., Son, J.-I., Yoon, O.-J., & Lee, H.-J. (2021). Bone Marrow Mesenchymal Stromal Cells on Silk Fibroin Scaffolds to Attenuate Polymicrobial Sepsis Induced by Cecal Ligation and Puncture. Polymers, 13(9), 1433. https://doi.org/10.3390/polym13091433