Immunomodulatory and Anti-Inflammatory Effects of Fucoidan: A Review

 ,
,  ,
,  ,
,  , ,
, ,  ,
,  ,
,  and
and 
Abstract
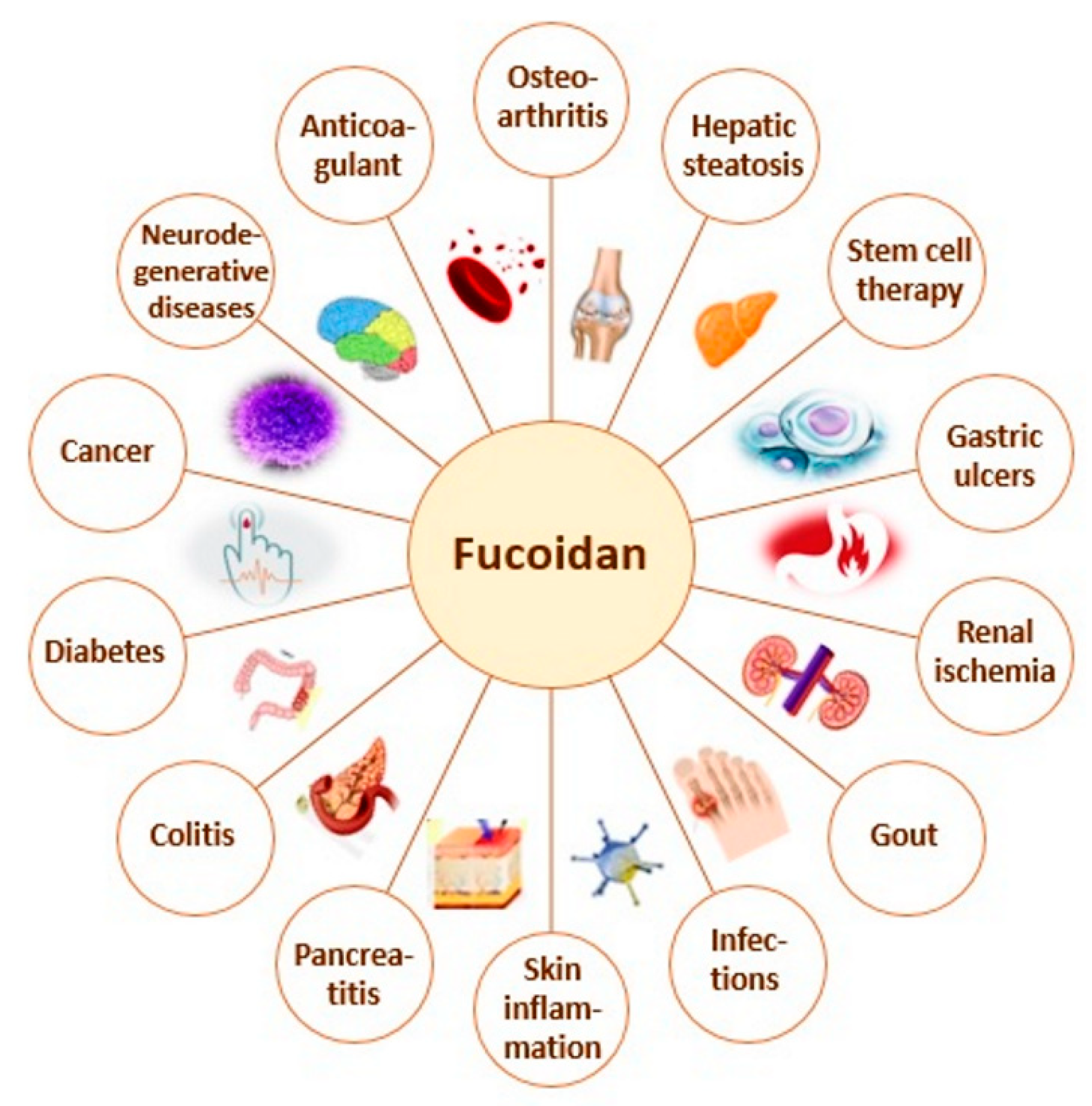
1. Introduction
2. Materials and Methods
3. An Overview of Human Immune System and Inflammation
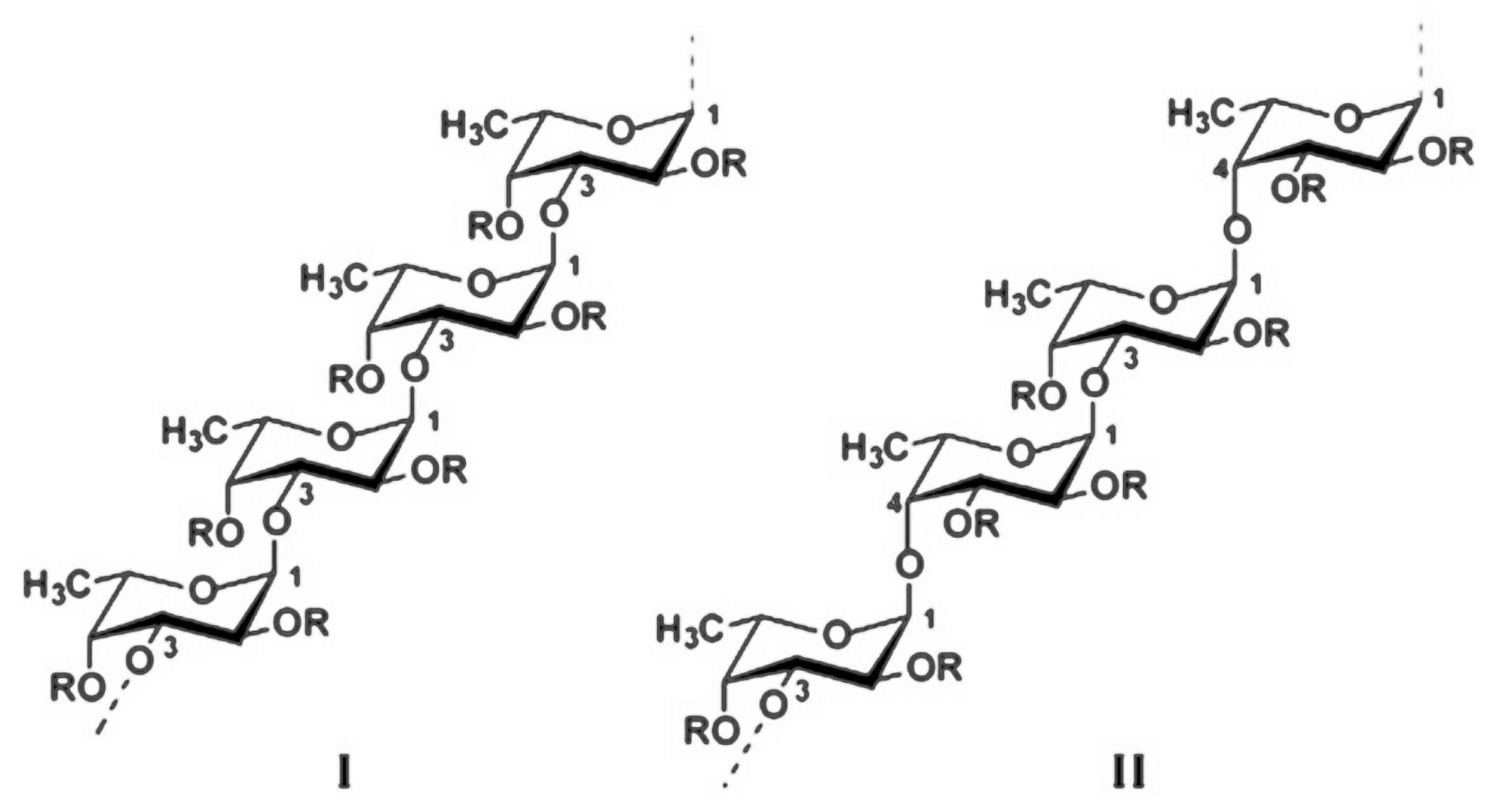
4. Structure and Structure—Activity Relationship of Fucoidan
5. Pharmacokinetic of Fucoidan
6. Anti-Inflammatory Activity of Fucoidan: Evidence from In Vitro and In Vivo Models
6.1. In Vitro Studies of the Anti-Inflammatory Effect of Fucoidan on Cell Lines
6.1.1. RAW 264.7 Macrophages
6.1.2. Human Keratinocyte Cell Line (HaCaT)
6.1.3. Rat Primary Microglia
6.1.4. Caco-2 Cell Line and Caco-2/RAW 264.7 Coculture
6.1.5. Other Models
6.2. In Vivo Studies of the Effects of Fucoidan on Inflammation and Cytokine Levels in Animal Models
6.2.1. Diabetes Mellitus in Rodents
6.2.2. Chronic Colitis in Mice
6.2.3. Inflammation in Zebrafish Embryos
6.2.4. Rheumatoid Arthritis in Mice and Rats
6.2.5. Murine Paw Edema
6.2.6. Myocardial Infarction in Rats
6.2.7. Liver Damage in Rodents
6.2.8. Other Models
6.3. Clinical Trials
7. Immunomodulatory Effect of Fucoidan
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hou, C.; Chen, L.; Yang, L.; Ji, X. An insight into anti-inflammatory effects of natural polysaccharides. Int. J. Biol. Macromol. 2020, 153, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.W.; Hwang, S.J.; Han, M.H.; Lee, D.S.; Yoo, J.S.; Choi, I.W.; Cha, H.J.; Kim, S.; Kim, H.S.; Kim, G.Y.; et al. Fucoidan inhibits lipopolysaccharide-induced inflammatory responses in RAW 264.7 macrophages and zebrafish larvae. Mol. Cell. Toxicol. 2017, 13, 405–417. [Google Scholar] [CrossRef]
- Muralidharan, S.; Mandrekar, P. Cellular stress response and innate immune signaling: Integrating pathways in host defense and inflammation. J. Leukoc. Biol. 2013, 94, 1167–1184. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xing, M.; Cao, Q.; Ji, A.; Liang, H.; Song, S. Biological activities of fucoidan and the factors mediating its therapeutic effects: A review of recent studies. Mar. Drugs 2019, 17, 183. [Google Scholar] [CrossRef] [PubMed]
- Ale, M.T.; Mikkelsen, J.D.; Meyer, A.S. Important determinants for fucoidan bioactivity: A critical review of structure-function relations and extraction methods for fucose-containing sulfated polysaccharides from brown seaweeds. Mar. Drugs. 2011, 9, 2106–2130. [Google Scholar] [CrossRef] [PubMed]
- Hentati, F.; Delattre, C.; Ursu, A.V.; Desbrières, J.; Le Cerf, D.; Gardarin, C.; Abdelkafi, S.; Michaud, P.; Pierre, G. Structural characterization and antioxidant activity of water-soluble polysaccharides from the Tunisian brown seaweed Cystoseira compressa. Carbohydr. Polym. 2018, 198, 589–600. [Google Scholar] [CrossRef]
- Balboa, E.M.; Conde, E.; Moure, A.; Falqué, E.; Domínguez, H. In vitro antioxidant properties of crude extracts and compounds from brown algae. Food Chem. 2013, 138, 1764–1785. [Google Scholar] [CrossRef]
- Ale, M.T.; Meyer, A.S. Fucoidans from brown seaweeds: An update on structures, extraction techniques and use of enzymes as tools for structural elucidation. RSC Adv. 2013, 3, 8131–8141. [Google Scholar] [CrossRef]
- Aleissa, M.S.; Alkahtani, S.; Abd Eldaim, M.A.; Ahmed, A.M.; Bungău, S.G.; Almutairi, B.; Bin-Jumah, M.; AlKahtane, A.A.; Alyousif, M.S.; Abdel-Daim, M.M. Fucoidan ameliorates oxidative stress, inflammation, DNA damage, and hepatorenal injuries in diabetic rats intoxicated with aflatoxin B1. Oxidative Med. Cell. Longev. 2020, 2020, 9316751. [Google Scholar] [CrossRef]
- Myers, S.P.; Mulder, A.M.; Baker, D.G.; Robinson, S.R.; Rolfe, M.I.; Brooks, L.; Fitton, J.H. Effects of fucoidan from Fucus vesiculosus in reducing symptoms of osteoarthritis: A randomized placebo-controlled trial. Biol. Targets Ther. 2016, 10, 81–88. [Google Scholar] [CrossRef]
- Fitton, J.H. Therapies from fucoidan; multifunctional marine polymers. Mar. Drugs 2011, 9, 1731–1760. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Lu, F.; Wei, X.; Zhao, R. Fucoidan: Structure and bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, M.A. A brief review of the basics of immunology: The innate and adaptive response. Vet. Clin. Small Anim. Pract. 2010, 40, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Kyung, J.; Kim, D.; Park, D.; Yang, Y.H.; Choi, E.K.; Lee, S.P.; Kim, T.S.; Lee, Y.B.; Kim, Y.B. Synergistic anti-inflammatory effects of Laminaria japonica fucoidan and Cistanche tubulosa extract. Lab. Anim. Res. 2012, 28, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Fernando, I.S.; Sanjeewa, K.A.; Samarakoon, K.W.; Lee, W.W.; Kim, H.S.; Kang, N.; Ranasinghe, P.; Lee, H.S.; Jeon, Y.J. A fucoidan fraction purified from Chnoospora minima; a potential inhibitor of LPS-induced inflammatory responses. Int. J. Biol. Macromol. 2017, 104, 1185–1193. [Google Scholar] [CrossRef] [PubMed]
- Ni, L.; Wang, L.; Fu, X.; Duan, D.; Jeon, Y.J.; Xu, J.; Gao, X. In vitro and in vivo anti-inflammatory activities of a fucose-rich fucoidan isolated from Saccharina japonica. Int. J. Biol. Macromol. 2020, 156, 717–729. [Google Scholar] [CrossRef]
- Parkin, J.; Cohen, B. An overview of the immune system. Lancet 2001, 357, 1777–1789. [Google Scholar] [CrossRef]
- Cui, Y.Q.; Zhang, L.J.; Zhang, T.; Luo, D.Z.; Jia, Y.J.; Guo, Z.X.; Zhang, Q.B.; Wang, X.; Wang, X.M. Inhibitory effect of fucoidan on nitric oxide production in lipopolysaccharide-activated primary microglia. Clin. Exp. Pharmacol. Physiol. 2010, 37, 422–428. [Google Scholar] [CrossRef]
- Citkowska, A.; Szekalska, M.; Winnicka, K. Possibilities of fucoidan utilization in the development of pharmaceutical dosage forms. Mar. Drugs 2019, 17, 458. [Google Scholar] [CrossRef]
- Cumashi, A.; Ushakova, N.A.; Preobrazhenskaya, M.E.; D’Incecco, A.; Piccoli, A.; Totani, L.; Tinari, N.; Morozevich, G.E.; Berman, A.E.; Bilan, M.I.; et al. A comparative study of the anti-inflammatory, anticoagulant, antiangiogenic, and antiadhesive activities of nine different fucoidans from brown seaweeds. Glycobiology 2007, 17, 541–552. [Google Scholar] [CrossRef]
- Vo, T.S.; Kim, S.K. Fucoidans as a natural bioactive ingredient for functional foods. J. Funct. Foods 2013, 5, 16–27. [Google Scholar] [CrossRef]
- Lin, Y.; Qi, X.; Liu, H.; Xue, K.; Xu, S.; Tian, Z. The anti-cancer effects of fucoidan: A review of both in vivo and in vitro investigations. Cancer Cell Int. 2020, 20, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Flórez-Fernández, N.; Torres, M.D.; González-Muñoz, M.J.; Domínguez, H. Potential of intensification techniques for the extraction and depolymerization of fucoidan. Algal Res. 2018, 30, 128–148. [Google Scholar] [CrossRef]
- Ponce, N.M.A.; Pujol, C.A.; Damonte, E.B. Fucoidans from the brown seaweed Adenocystis utricularis: Extraction methods, antiviral activity and structural studies. Carbohydr. Res. 2003, 338, 153–165. [Google Scholar] [CrossRef]
- Maruyama, H.; Tamauchi, H.; Kawakami, F.; Yoshinaga, K.; Nakano, T. Suppressive effect of dietary fucoidan on proinflammatory immune response and MMP-1 expression in UVB-irradiated mouse skin. Planta Med. 2015, 81, 1370–1374. [Google Scholar] [CrossRef]
- Matsumoto, S.; Nagaoka, M.; Hara, T.; Kimura-Takagi, I.; Mistuyama, K.; Ueyama, S. Fucoidan derived from Cladosiphon okamuranus Tokida ameliorates murine chronic colitis through the down-regulation of interleukin 6 production on colonic epithelial cells. Clin. Exp. Immunol. 2004, 136, 432–439. [Google Scholar] [CrossRef]
- Zhang, W.; Oda, T.; Yu, Q.; Jin, J.O. Fucoidan from Macrocystis pyrifera has powerful immune-modulatory effects compared to three other fucoidans. Mar. Drugs 2015, 13, 1084–1104. [Google Scholar] [CrossRef]
- Manikandan, R.; Parimalanandhini, D.; Mahalakshmi, K.; Beulaja, M.; Arumugam, M.; Janarthanan, S.; Palanisamy, S.; You, S.; Prabhu, N.M. Studies on isolation, characterization of fucoidan from brown algae Turbinaria decurrens and evaluation of it’s in vivo and in vitro anti-inflammatory activities. Int. J. Biol. Macromol. 2020, 160, 1263–1276. [Google Scholar] [CrossRef]
- Menshova, R.V.; Shevchenko, N.M.; Imbs, T.I.; Zvyagintseva, T.N.; Malyarenko, O.S.; Zaporoshets, T.S.; Besednova, N.N.; Ermakova, S.P. Fucoidans from brown alga Fucus evanescens: Structure and biological activity. Front. Mar. Sci. 2016, 3, 129. [Google Scholar] [CrossRef]
- Haroun-Bouhedja, F.; Ellouali, M.; Sinquin, C.; Boisson-Vidal, C. Relationship between sulfate groups and biological activities of fucans. Thromb. Res. 2000, 100, 453–459. [Google Scholar] [CrossRef]
- Park, S.B.; Chun, K.R.; Kim, J.K.; Suk, K.; Jung, Y.M.; Lee, W.H. The differential effect of high and low molecular weight fucoidans on the severity of collagen-induced arthritis in mice. Phytother. Res. 2010, 24, 1384–1391. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.Y.; Moon, S.Y.; Joo, H.G. Differential effects of fucoidans with low and high molecular weight on the viability and function of spleen cells. Food Chem. Toxicol. 2014, 68, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Rioux, L.E.; Moulin, V.; Beaulieu, M.; Turgeon, S.L. Human skin fibroblast response is differentially regulated by galactofucan and low molecular weight galactofucan. Bioact. Carbohydr. Diet. Fibre 2013, 1, 105–110. [Google Scholar] [CrossRef]
- Hwang, P.A.; Phan, N.N.; Lu, W.J.; Ngoc Hieu, B.T.; Lin, Y.C. Low-molecular-weight fucoidan and high-stability fucoxanthin from brown seaweed exert prebiotics and anti-inflammatory activities in Caco-2 cells. Food Nutr. Res. 2016, 60, 32033. [Google Scholar] [CrossRef] [PubMed]
- Sanjeewa, K.A.; Fernando, I.P.S.; Kim, E.A.; Ahn, G.; Jee, Y.; Jeon, Y.J.; Noh, H.J.; Koh, H.B.; Kim, H.K.; Cho, H.H.; et al. Anti-inflammatory activity of a sulfated polysaccharide isolated from an enzymatic digest of brown seaweed Sargassum horneri in RAW 264.7 cells. Nutr. Res. Pr. 2017, 11, 3–10. [Google Scholar] [CrossRef]
- Irhimeh, M.R.; Fitton, J.H.; Lowenthal, R.M.; Kongtawelert, P. A quantitative method to detect fucoidan in human plasma using a novel antibody. Methods Find. Exp. Clin. Pharmacol. 2005, 27, 705–710. [Google Scholar] [CrossRef]
- Tokita, Y.; Nakajima, K.; Mochida, H.; Iha, M.; Nagamine, T. Development of a fucoidan-specific antibody and measurement of fucoidan in serum and urine by sandwich ELISA. Biosci. Biotechnol. Biochem. 2010, 74, 350–357. [Google Scholar] [CrossRef]
- Nagamine, T.; Nakazato, K.; Tomioka, S.; Iha, M.; Nakajima, K. Intestinal absorption of fucoidan extracted from the brown seaweed, cladosiphon okamuranus. Mar. Drugs 2015, 13, 48–64. [Google Scholar] [CrossRef]
- Fitton, J.H.; Stringer, D.N.; Karpiniec, S.S. Therapies from fucoidan: An update. Mar. Drugs 2015, 13, 5920–5946. [Google Scholar] [CrossRef]
- Kimura, R.; Rokkaku, T.; Takeda, S.; Senba, M.; Mori, N. Cytotoxic effects of fucoidan nanoparticles against osteosarcoma. Mar. Drugs 2013, 11, 4267–4278. [Google Scholar] [CrossRef]
- Pozharitskaya, O.N.; Shikov, A.N.; Obluchinskaya, E.D.; Vuorela, H. The pharmacokinetics of fucoidan after topical application to rats. Mar. Drugs 2019, 17, 687. [Google Scholar] [CrossRef] [PubMed]
- Pozharitskaya, O.N.; Shikov, A.N.; Faustova, N.M.; Obluchinskaya, E.D.; Kosman, V.M.; Vuorela, H.; Makarov, V.G. Pharmacokinetic and tissue distribution of fucoidan from Fucus vesiculosus after oral administration to rats. Mar. Drugs 2018, 16, 132. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Sun, D.; Zhao, X.; Jin, W.; Wang, J.; Zhang, Q. Microanalysis and preliminary pharmacokinetic studies of a sulfated polysaccharide from Laminaria japonica. Chin. J. Oceanol. Limnol. 2016, 34, 177–185. [Google Scholar] [CrossRef]
- Zhang, X.W.; Liu, Q.; Thorlacius, H. Inhibition of selectin function and leukocyte rolling protects against dextran sodium sulfate-induced murine colitis. Scand. J. Gastroenterol. 2001, 36, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.C.; Sousa, R.B.; Franco, A.X.; Costa, J.V.; Neves, L.M.; Ribeiro, R.A.; Sutton, R.; Criddle, D.N.; Soares, P.M.; de Souza, M.H. Protective effects of fucoidan, a p- and l-selectin inhibitor, in murine acute pancreatitis. Pancreas 2014, 43, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Amin, M.L.; Mawad, D.; Dokos, S.; Koshy, P.; Martens, P.J.; Sorrell, C.C. Immunomodulatory properties of photopolymerizable fucoidan and carrageenans. Carbohydr. Polym. 2020, 230, 115691. [Google Scholar] [CrossRef]
- Lee, S.H.; Ko, C.I.; Ahn, G.; You, S.; Kim, J.S.; Heu, M.S.; Kim, J.; Jee, Y.; Jeon, Y.J. Molecular characteristics and anti-inflammatory activity of the fucoidan extracted from Ecklonia cava. Carbohydr. Polym. 2012, 89, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Sanjeewa, K.A.; Fernando, I.P.S.; Kim, S.Y.; Kim, H.S.; Ahn, G.; Jee, Y.; Jeon, Y.J. In vitro and in vivo anti-inflammatory activities of high molecular weight sulfated polysaccharide; containing fucose separated from Sargassum horneri. Int. J. Biol. Macromol. 2018, 107, 803–807. [Google Scholar] [CrossRef]
- Mizuno, M.; Nishitani, Y.; Hashimoto, T.; Kanazawa, K. Different suppressive effects of fucoidan and lentinan on IL-8 mRNA expression in in vitro gut inflammation. Biosci. Biotechnol. Biochem. 2009, 73, 2324–2325. [Google Scholar] [CrossRef]
- Ryu, M.J.; Chung, H.S. Anti-inflammatory activity of fucoidan with blocking NF-kappaB and STAT1 in human keratinocytes cells. Nat. Prod. Sci. 2015, 21, 205–209. [Google Scholar] [CrossRef]
- Lee, K.S.; Cho, E.; Weon, J.B.; Park, D.; Fréchet, M.; Chajra, H.; Jung, E. Inhibition of UVB-induced inflammation by Laminaria japonica extract via regulation of nc886-PKR pathway. Nutrients 2020, 12, 1958. [Google Scholar] [CrossRef] [PubMed]
- Park, H.Y.; Han, M.H.; Park, C.; Jin, C.Y.; Kim, G.Y.; Choi, I.W.; Kim, N.D.; Nam, T.J.; Kwon, T.K.; Choi, Y.H. Anti-inflammatory effects of fucoidan through inhibition of NF-κB, MAPK and Akt activation in lipopolysaccharide-induced BV2 microglia cells. Food Chem. Toxicol. 2011, 49, 1745–1752. [Google Scholar] [CrossRef] [PubMed]
- Bahar, B.; O’Doherty, J.V.; Smyth, T.J.; Ahmed, A.M.; Sweeney, T. A cold water extract of Fucus vesiculosus inhibits lipopolysaccharide (LPS) induced pro-inflammatory responses in the porcine colon ex-vivo model. Innov. Food Sci Emerg. Technol. 2016, 37, 229–236. [Google Scholar] [CrossRef]
- Yang, H.S.; Haj, F.G.; Lee, M.; Kang, I.; Zhang, G.; Lee, Y. Laminaria japonica extract enhances intestinal barrier function by altering inflammatory response and tight junction-related protein in lipopolysaccharide-stimulated caco-2 cells. Nutrients 2019, 11, 1001. [Google Scholar] [CrossRef] [PubMed]
- Pozharitskaya, O.N.; Obluchinskaya, E.D.; Shikov, A.N. Mechanisms of bioactivities of fucoidan from the brown seaweed Fucus vesiculosus L. of the Barents Sea. Mar. Drugs 2020, 18, 275. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, Q.; Luo, D.; Wang, J.; Duan, D. Low molecular weight fucoidan ameliorates the inflammation and glomerular filtration function of diabetic nephropathy. J. Appl. Phycol. 2017, 29, 531–542. [Google Scholar] [CrossRef]
- Lin, H.V.; Tsou, Y.C.; Chen, Y.T.; Lu, W.J.; Hwang, P.A. Effects of low-molecular-weight fucoidan and high stability fucoxanthin on glucose homeostasis, lipid metabolism, and liver function in a mouse model of type II diabetes. Mar. Drugs 2017, 15, 113. [Google Scholar] [CrossRef]
- Lean, Q.Y.; Eri, R.D.; Fitton, J.H.; Patel, R.P.; Gueven, N. Fucoidan extracts ameliorate acute colitis. PLoS ONE 2015, 10, e0128453. [Google Scholar] [CrossRef]
- Lee, S.H.; Ko, C.I.; Jee, Y.; Jeong, Y.; Kim, M.; Kim, J.S.; Jeon, Y.J. Anti-inflammatory effect of fucoidan extracted from Ecklonia cava in zebrafish model. Carbohydr. Polym. 2013, 92, 84–89. [Google Scholar] [CrossRef]
- Jeon, H.; Yoon, W.J.; Ham, Y.M.; Yoon, S.A.; Kang, S.C. Anti-arthritis effect through the anti-inflammatory effect of Sargassum muticum extract in collagen-induced arthritic (CIA) Mice. Molecules 2019, 24, 276. [Google Scholar] [CrossRef]
- Phull, A.R.; Kim, S.J. Fucoidan as bio-functional molecule: Insights into the anti-inflammatory potential and associated molecular mechanisms. J. Funct. Foods. 2017, 38, 415–426. [Google Scholar] [CrossRef]
- Ananthi, S.; Gayathri, V.; Malarvizhi, R.; Bhardwaj, M.; Vasanthi, H.R. Anti-arthritic potential of marine macroalgae Turbinaria ornata in complete freund’s adjuvant induced rats. Exp. Toxicol. Pathol. 2017, 69, 672–680. [Google Scholar] [CrossRef] [PubMed]
- Ammar, H.; Lajili, S.; Ben Said, R.; Le Cerf, D.; Bouraoui, A.; Majdoub, H. Physico-chemical characterization and pharmacological evaluation of sulfated polysaccharides from three species of Mediterranean brown algae of the genus Cystoseira. DARU J. Pharm. Sci. 2015, 23, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ananthi, S.; Raghavendran, H.R.; Sunil, A.G.; Gayathri, V.; Ramakrishnan, G.; Vasanthi, H.R. In vitro antioxidant and in vivo anti-inflammatory potential of crude polysaccharide from Turbinaria ornata (Marine Brown Alga). Food Chem. Toxicol. 2010, 48, 187–192. [Google Scholar] [CrossRef]
- Li, C.; Gao, Y.; Xing, Y.; Zhu, H.; Shen, J.; Tian, J. Fucoidan, a sulfated polysaccharide from brown algae, against myocardial ischemia-reperfusion injury in rats via regulating the inflammation response. Food Chem. Toxicol. 2011, 49, 2090–2095. [Google Scholar] [CrossRef]
- Lu, X.; Wang, Q.; Liu, Y.; Yang, S.; Tan, B.; Feng, Y.; Cao, C. Effect of fucoidan on NF-κB-mediated inflammatory response in rats with acute myocardial infarction. J. Biomater. Tissue Eng. 2019, 9, 1376–1380. [Google Scholar] [CrossRef]
- Lim, J.D.; Lee, S.R.; Kim, T.; Jang, S.A.; Kang, S.C.; Koo, H.J.; Sohn, E.; Bak, J.P.; Namkoong, S.; Kim, H.K.; et al. Fucoidan from Fucus vesiculosus protects against alcohol-induced liver damage by modulating inflammatory mediators in mice and HepG2 cells. Mar. Drugs 2015, 13, 1051–1067. [Google Scholar] [CrossRef]
- Meenakshi, S.; Umayaparvathi, S.; Saravanan, R.; Manivasagam, T.; Balasubramanian, T. Hepatoprotective effect of fucoidan isolated from the seaweed Turbinaria decurrens in ethanol intoxicated rats. Int. J. Biol. Macromol. 2014, 67, 367–372. [Google Scholar] [CrossRef]
- AlKahtane, A.A.; Abushouk, A.I.; Mohammed, E.T.; ALNasser, M.; Alarifi, S.; Ali, D.; Alessia, M.S.; Almeer, R.S.; AlBasher, G.; Alkahtani, S.; et al. Fucoidan alleviates microcystin-LR-induced hepatic, renal, and cardiac oxidative stress and inflammatory injuries in mice. Environ. Sci Pollut. Res. 2020, 27, 2935–2944. [Google Scholar] [CrossRef]
- Chale-Dzul, J.; de Vaca, R.P.C.; Quintal-Novelo, C.; Olivera-Castillo, L.; Moo-Puc, R. Hepatoprotective effect of a fucoidan extract from Sargassum fluitans Borgesen against CCl4-induced toxicity in rats. Int. J. Biol. Macromol. 2020, 145, 500–509. [Google Scholar] [CrossRef]
- Phull, A.R.; Majid, M.; Haq, I.U.; Khan, M.R.; Kim, S.J. In vitro and in vivo evaluation of anti-arthritic, antioxidant efficacy of fucoidan from Undaria pinnatifida (Harvey) Suringar. Int. J. Biol. Macromol. 2017, 97, 468–480. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Ren, D.; Song, Y.; Wu, L.; He, Y.; Peng, Y.; Zhou, H.; Liu, S.; Cong, H.; Zhang, Z.; et al. Gastric protective activities of fucoidan from brown alga Kjellmaniella crassifolia through the NF-κB signaling pathway. Int. J. Biol. Macromol. 2020, 149, 893–900. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Cha, J.D.; Choi, K.M.; Lee, K.Y.; Han, K.M.; Jang, Y.S. Fucoidan inhibits LPS-induced inflammation in vitro and during the acute response in vivo. Int. Immunopharmacol. 2017, 43, 91–98. [Google Scholar] [CrossRef]
- Subash, A.; Veeraraghavan, G.; Sali, V.K.; Bhardwaj, M.; Vasanthi, H.R. Attenuation of inflammation by marine algae Turbinaria ornata in cotton pellet induced granuloma mediated by fucoidan like sulfated polysaccharide. Carbohydr. Polym. 2016, 151, 1261–1268. [Google Scholar] [CrossRef] [PubMed]
- Myers, S.P.; O’Connor, J.; Fitton, J.H.; Brooks, L.; Rolfe, M.; Connellan, P.; Wohlmuth, H.; Cheras, P.A.; Morris, C. A combined phase I and II open label study on the effects of a seaweed extract nutrient complex on osteoarthritis. Biologics 2010, 4, 33–44. [Google Scholar] [CrossRef]
- Jin, J.O.; Zhang, W.; Du, J.Y.; Wong, K.W.; Oda, T.; Yu, Q. Fucoidan can function as an adjuvant in vivo to enhance dendritic cell maturation and function and promote antigen-specific t cell immune responses. PLoS ONE 2014, 9, e99396. [Google Scholar] [CrossRef]
- Makarenkova, I.D.; Logunov, D.Y.; Tukhvatulin, A.I.; Semenova, I.B.; Besednova, N.N.; Zvyagintseva, T.N. Interactions between sulfated polysaccharides from sea brown algae and toll-like receptors on hek293 eukaryotic cells in vitro. Bull. Exp. Biol. Med. 2012, 154, 241–244. [Google Scholar] [CrossRef]
- Hayashi, K.; Nakano, T.; Hashimoto, M.; Kanekiyo, K.; Hayashi, T. Defensive effects of a fucoidan from brown alga Undaria pinnatifida against herpes simplex virus infection. Int. Immunopharmacol. 2008, 8, 109–116. [Google Scholar] [CrossRef]
- Tabarsa, M.; Dabaghian, E.H.; You, S.; Yelithao, K.; Cao, R.; Rezaei, M.; Alboofetileh, M.; Bita, S. The activation of NF-κB and MAPKs signaling pathways of RAW264.7 murine macrophages and natural killer cells by fucoidan from Nizamuddinia zanardinii. Int. J. Biol. Macromol. 2020, 148, 56–67. [Google Scholar] [CrossRef]
- Jiang, Z.; Okimura, T.; Yamaguchi, K.; Oda, T. The potent activity of sulfated polysaccharide, ascophyllan, isolated from Ascophyllum nodosum to induce nitric oxide and cytokine production from mouse macrophage RAW264.7 cells: Comparison between ascophyllan and fucoidan. Nitric Oxide 2011, 25, 407–415. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Source | Compound | Cell Line | Concentration | Activity | Reference |
|---|---|---|---|---|---|
| Fucus vesiculosus | Commercially available fucoidan | Murine RAW 264.7 macrophages | 50–100 µg/mL | Decreased secretion of NO, PGE2; diminished secretion of TNF-α and IL-1β | [2] |
| Commercially available fucoidan | BV2 microglial cells | 100 µg/mL | Decreased production of NO, PGE2, IL-1β, and TNF-α; inhibition of NF-κB, Akt, ERK, p38 MAPK, and JNK pathways | [52] | |
| Commercially available fucoidan | Human keratinocyte (HaCaT) | 1–50 µg/mL | Inhibition of NF-κB, STAT1 and ERK ½ pathways; decreased production of IL-1β and IL-6 after TNF- α/ IFN-γ stimulation | [50] | |
| Methacrylate-functionalized fucoidan | RAW 264.7 macrophages; THP-1 macrophages | 100 µg/mL | Decreased production of NO, decreased production of CD86 after LPS/IFN-γ stimulation | [46] | |
| Cold water extract | Caco-2 | 1 mg/mL | Decreased production of IL-8 after TNF-α stimulation | [53] | |
| Fuc:Glc:Gal:Xyl:Man = 1.0:0.16:0.05:0.09: 0.03:0.03 | Human recombinant COX1 and COX2 assay kit | 0.1–10 µg/mL | Inhibition of COX1 and COX2, predominantly of COX2 | [55] | |
| Laminaria japonica | LMWF (MW = 7 kDa); Fuc:Gal fraction = 1.0:0.24 | Rat primary microglia | 125 µg/mL | Reduced synthesis of NO and expression of iNOS; decreased phosphorylation of p38 and ERK MAPK | [18] |
| HMWF containing Fuc 79.49%, Gal 16.76%, Rha 0.82%, Xyl 1.08%, Man 1.84%; sulfate content 30.72%; MW = 104.3 kDa | RAW 264.7 macrophages | 25 µg/mL | Decreased production of TNF-α, IL-1β, IL-6, NO, iNOS, and COX-2 expression; downregulation of MAPK and NF-κB signaling pathways | [16] | |
| Lyophilized water extract | Human keratinocyte (HaCaT) | 100 µg/mL | Decreased gene expression of COX2; diminished secretion of PGE2, TNF-α and IL-8 stimulated after UVB irradiation | [51] | |
| No information added | Caco-2/RAW 264.7 coculture | 100 µg/mL | Decreased production of TNF-α and IL-8 mRNA expression | [49] | |
| Four types of water extracts (with or without fermentation) | Caco-2 | 100 µg/mL | Decreased production of NO and IL-6 by all four extracts | [54] | |
| Sargassum hemiphyllum | LMWF (MW = 0.8 kDa), sulfate content 38.9% | Caco-2 | 100 µg/mL | Decreased TNF-α and IL-1β; increased IL-10 and IFN-γ | [34] |
| Sargassum horneri | HMWF; polysaccharide fraction (MW > 30 kDa) | RAW 264.7 macrophages | 25–100 µg/mL | Decreased production of TNF-α, IL-6, NO and PGE2 | [35] |
| Chnoospora minima | Fucose-rich polysaccharide fraction (79.32% Fuc) | RAW 264.7 macrophages | 25–100 µg/mL | Decreased production of PGE2, TNF-α, IL-1β, IL-6, NO, iNOS and COX-2 expression | [15] |
| Cladosiphon okamuranus Tokida | Fucoidan containing Fuc:GlcA:sulfate groups = 6.1:1.0:2.9 | Murine colon carcinoma cell line CMT-93 | 2.5 µg/mL | Decreased levels of IL-6; attenuated activation of the NF-κB signaling pathway | [26] |
| Ecklonia cava | LMWF rich in fucose (77.9% Fuc), sulfate content 39.1% | RAW 264.7 macrophages | 50–100 µg/mL | Reduced NO production and levels of TNF-α, IL-1β, and IL-6 | [47] |
| Source | Composition | Model | DosesTested | Effective Doses | Positive Control | Treatment | Activity | Reference |
|---|---|---|---|---|---|---|---|---|
| Fucus vesiculosus | Extract 1: Fucoidan polyphenol complex (MW = 203 kDa, sulfate content 21.8%) Extract 2: high purity fucoidan (MW = 62 kDa, sulfate content 26.6%) | Acute colitis in C57BL/6 mice induced by oral intake of 3% w/v of dextran sulphate sodium for 7 consecutive days | Extract 1 in dose 400 mg/kg daily p.o.; Extract 2 in dose 10 mg/kg daily p.o. or i.p. | Extract 1 in dose 400 mg/kg daily p.o.; Extract 2 in dose 10 mg/kg daily p.o. | No | Oral or i.p. application for 7 days | Lowered levels of IL-1α, IL-1β, and IL-10 | [58] |
| Commercially available fucoidan | Acute pancreatitis in mice induced by cerulein or taurolithocholic acid sulfate | 25 mg/kg | 25 mg/kg | No | Intravenously; single dose, 30 min before application of cerulein or taurolithocholic acid sulfate | Decreased levels of IL-1β, TNF-α, and myeloperoxidase; inhibited neutrophil migration | [45] | |
| Commercially available fucoidan | Alcohol-induced liver damage in mice | 30 and 60 mg/kg | 30 and 60 mg/kg | No | Orally; 7 days of application | Decreased production of TGF-β1 and COX-2 in mice livers | [67] | |
| Laminaria japonica | Commercially available fucoidan | Microcystin-LR-induced liver damage in mice | 50 and 100 mg/kg | 50 and 100 mg/kg | No | Orally; 21 days of application | Reduced levels of TNF-α, IL-1β, and IL-6 | [69] |
| No information | Myocardial infarction in rats | 50, 100, and 200 mg/kg | 100 and 200 mg/kg | No | Orally; 7 days of pretreatment before the surgery | Decreased levels of TNF- α and IL-6; increased levels of IL-10 | [65] | |
| Commercially available fucoidan | Diabetes mellitus in rats, induced by 50 mg/kg STZ (i.p.) | 100 mg/kg | 100 mg/kg | No | Orally; daily application between fifth to eighth week after STZ injection | Reduced blood glucose level and serum levels of IL-1β, IL-6, and TNF-α | [9] | |
| Sargassum muticum | Ethyl alcohol (70%) extract | Collagen-induced rheumatoid arthritis in mice | 50, 100, and 200 mg/kg | 100 and 200 mg/kg reduce TNF-α and IL-6 levels; 50, 100, and 200 mg/kg reduce TNF-α, IFN-γ, and IL-6 | Joins® 10 mg/kg | Orally; 77-day treatment (21–98 day of the experiment) | Decreased levels of TNF-α, IFN-γ, and IL-6 | [60] |
| Sargassum fluitans Borgesen | Water extract, 7.56% sulfate content | Carbon tetrachloride-induced liver injury in rats | 50 mg/kg | 50 mg/kg | Silymarin 100 mg/kg p.o. | Orally; 1 week of pretreatment + 12 weeks treatment | Decreased levels of TNF-α and IL-1β | [70] |
| Cladosiphon okamuranus Tokida | Fucoidan containing Fuc:GlcA:sulfate groups = 6.1:1.0:2.9 | Chronic colitis in mice induced by 4% dextran sodium sulphate (DSS) (p.o.) | No information | No information | No | Orally, as fucoidan containing chow; treatment during the whole 2-week experiment | Decreased levels of IL-6 and increased levels of IL-10 | [26] |
| Chnoospora minima | Fucose-rich polysaccharide fraction (79.32% Fuc) | LPS-stimulated zebrafish embryos | 12.5, 25, and 50 μg/mL added to the embryo media | 12.5, 25, and 50 μg/mL | No | 1-h long treatment on the 8th hour of postfertilization | Decreased production of NO, ROS, COX-2, and iNOS expression | [15] |
| Ecklonia cava | Extract (sulfate content 20.1%, Fuc 61.1%, Rha 3.9%, Gal 27.2%, Glc 0.8%, and Xyl 7%) | LPS-stimulated zebrafish embryos | 100 μg/mL added to the embryo media | 100 μg/mL added to the embryo media | Commercially available fucoidan | 1-h long treatment on the 8th hour of postfertilization | Reduced production of NO | [59] |
| Undaria pinnatifida | LMWF, medium molecular weight fucoidan (MMWF), and HMWF | Collagen-induced rheumatoid arthritis in mice | 300 mg/kg | 300 mg/kg | No | Orally; 49 days treatment | LMWF reduced cartilage and bone destruction, and the tissue infiltration with inflammatory cells | [31] |
| Turbinaria ornata | Water extract and sulfated polysaccharide | Complete Freund’s adjuvant (CFA)-induced arthritis in rats | Water extract: 30, 100, and 300 mg/kg; sulfated polysaccharide 2.5, 5, and 10 mg/kg | Water extract 100 mg/kg and sulfated polysaccharide 5 and 10 mg/kg decreased TNF-α, IL-6, and PGE2 levels | Dexamethasone 100 μg/kg p.o. | Orally; pretreatment: two times a day for 7 days before CFA injection | Decreased levels of TNF-α, IL-6, and PGE2 | [62] |
| Turbinaria decurrens | Extract; sulfate content 23.51%, Fuc 59.3%, Gal 12.6%, Man 9.6% Rha 6.4%, Xyl 11.4% | Formalin-induced paw edema in mice | 50 mg/kg | 50 mg/kg | Dexamethasone 2.5 mg/kg i.p. | Orally; 5 days of pretreatment | Reduced the expression of genes of COX-2, IL-1β, the NF-κB signaling pathway | [28] |
| Source | Compound | Cell Line/Lines | Tested Concentrations | Activity | Reference |
|---|---|---|---|---|---|
| Laminaria japonica | Fuc:Gal:Man:Xyl:Glc = 65:20:8:4:3 MW = 10–30 kDa | Human embryonic kidney cells (HEK293-null, HEK293-TLR2/CD14, HEK293-hTLR4/CD14-MD2, and HEK293-hTLR5) | 1 mg/mL; 100 µg/mL; 10 µg/mL; 1 µg/mL; 100 ng/mL; 10 ng/mL of each extract | Specific activation of Toll-like receptors (TLR) 2 and following activation of NF-κB pathways is observed for L. japonica fucoidan (1 mg/mL), L. cichorioides fucoidan (100 μg/mL and 1 mg/mL), and F. evanescens fucoidan (10 μg/mL−1 mg/mL); activation of TLR-4 and following activation of NF-κB pathways is registered for L. japonica fucoidan (100 μg/mL and 1 mg/mL), L. cichorioides fucoidan (10 μg/mL−1 mg/mL), and F. evanescens fucoidan (1 μg/mL−1 mg/mL). | [77] |
| Laminaria cichorioides | Completely sulfated fucoidan MW = 40–80 kDa | ||||
| Fucus evanescens | Gal:Xyl:Man = 70:9:10.8 MW = 40–60 kDa | ||||
| Ascophyllum nodosum | Fuc:Xyl:Glc:Man:Gal = 39.80:3.68:0.88:0.72:3.37 | Human neutrophils | 50 μg/mL | All fucoidans significantly increased the production of IL-6, IL-8, and TNF-α from neutrophils | [27] |
| Macrocystis pyrifera | Fuc:Xyl:Glc:Man:Gal = 25.77:0.84:1.14:1.12:3.93 | ||||
| Undaria pinnatifida | Fuc:Xyl:Glc:Man:Gal = 28.27:0.45:0.49:0.30:24.94 | ||||
| Fucus vesiculosus | Fuc:Xyl:Glc:Man:Gal = 38.02:2.73:0.49:1.27:3.38 | ||||
| Nizamuddinia zanardinii | Fuc:Xyl:Man:Gal = 38.1:15.2:33.2:13.4 | RAW 264.7 murine macrophage | 10, 25, and 50 μg/mL | Increased secretion of NO, TNF-α, IL-1β, and IL-6; activation of NK cells, NF-κB, and MAPKs signaling pathways with subsequent release of TNF-α and INF-γ | [79] |
| Source | Composition | Model | Dose Tested | Effective Dose | Positive Control | Treatment | Activity | Reference |
|---|---|---|---|---|---|---|---|---|
| Fucus vesiculosus | Commercially available fucoidan | C57BL/6 mice | 10 mg/kg | 10 mg/kg | No | Single dose injected i.p. | Increased levels of TNF-α and IL-6 in spleens and blood serum | [76] |
| Undaria pinnatifida | Fuc:Gal = 1.0:1.1; MW = 9000 Da | 5-fluorouracil induces immunosuppression in BALB/c mice followed by herpes simplex virus-1 inoculation | 10 mg/mouse per day | 10 mg/mouse per day | No | 7 days of oral treatment | Increased activity of NK cells | [78] |
| Ascophyllum nodosum | Fuc:Xyl:Glc:Man:Gal = 39.80:3.68: 0.88:0.72:3.37 | C57BL/6 mice | 50 mg/kg | 50 mg/kg | No | 4 days of intraperitoneal treatment | M. pyrifera fucoidan increased the maturation and activation of NK cells | [27] |
| Macrocystis pyrifera | Fuc:Xyl:Glc:Man:Gal = 25.77:0.84: 1.14:1.12:3.93 | |||||||
| Undaria pinnatifida | Fuc:Xyl:Glc:Man:Gal = 28.27:0.45: 0.49:0.30:24.94 | |||||||
| Fucus vesiculosus | Fuc:Xyl:Glc:Man:Gal = 38.02:2.73: 0.49:1.27:3.38 | |||||||
| Undaria pinnatifida | Gal 20.7%, Fuc 24.4%, uronic acid 8.2% and ester sulfate 31.3% | Skin of BALB/c mice after UVB irradiation | 500 mg/kg | 500 mg/kg | Vit. C 600 mg/kg weight | 12 days of oral treatment | Increased lowered levels of IFN-γ after irradiation; reduced skin edema and leukocyte migration; no significant changes in IL-4, IL-6, TNF-α, and NF-κB expression | [25] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Apostolova, E.; Lukova, P.; Baldzhieva, A.; Katsarov, P.; Nikolova, M.; Iliev, I.; Peychev, L.; Trica, B.; Oancea, F.; Delattre, C.; et al. Immunomodulatory and Anti-Inflammatory Effects of Fucoidan: A Review. Polymers 2020, 12, 2338. https://doi.org/10.3390/polym12102338
Apostolova E, Lukova P, Baldzhieva A, Katsarov P, Nikolova M, Iliev I, Peychev L, Trica B, Oancea F, Delattre C, et al. Immunomodulatory and Anti-Inflammatory Effects of Fucoidan: A Review. Polymers. 2020; 12(10):2338. https://doi.org/10.3390/polym12102338
Chicago/Turabian StyleApostolova, Elisaveta, Paolina Lukova, Alexandra Baldzhieva, Plamen Katsarov, Mariana Nikolova, Ilia Iliev, Lyudmil Peychev, Bogdan Trica, Florin Oancea, Cédric Delattre, and et al. 2020. "Immunomodulatory and Anti-Inflammatory Effects of Fucoidan: A Review" Polymers 12, no. 10: 2338. https://doi.org/10.3390/polym12102338
APA StyleApostolova, E., Lukova, P., Baldzhieva, A., Katsarov, P., Nikolova, M., Iliev, I., Peychev, L., Trica, B., Oancea, F., Delattre, C., & Kokova, V. (2020). Immunomodulatory and Anti-Inflammatory Effects of Fucoidan: A Review. Polymers, 12(10), 2338. https://doi.org/10.3390/polym12102338