DNA Phase Transition in Charge Neutralization and Comformation Induced by Trivalent-Hydrolysed Metal Ions

Abstract
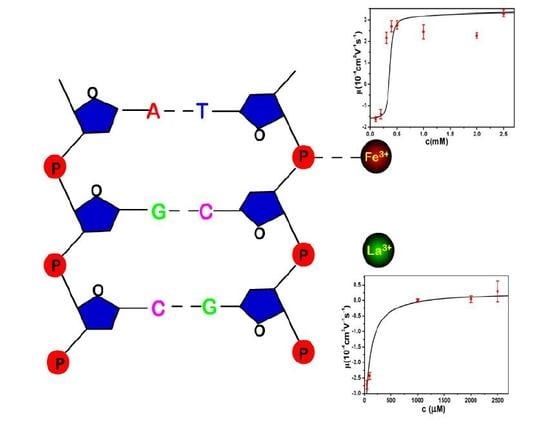
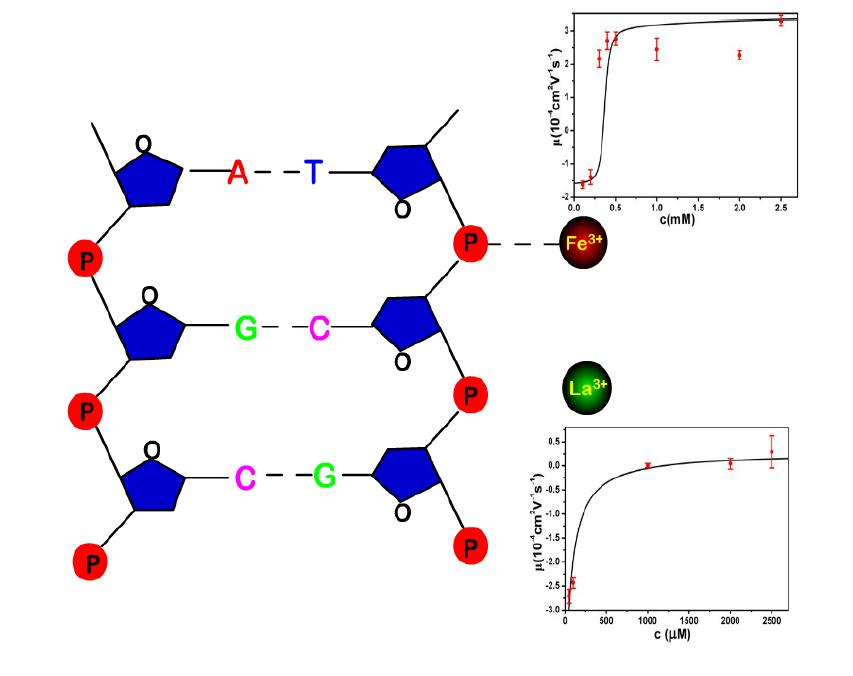
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Experimental Procedures
2.1. Materials
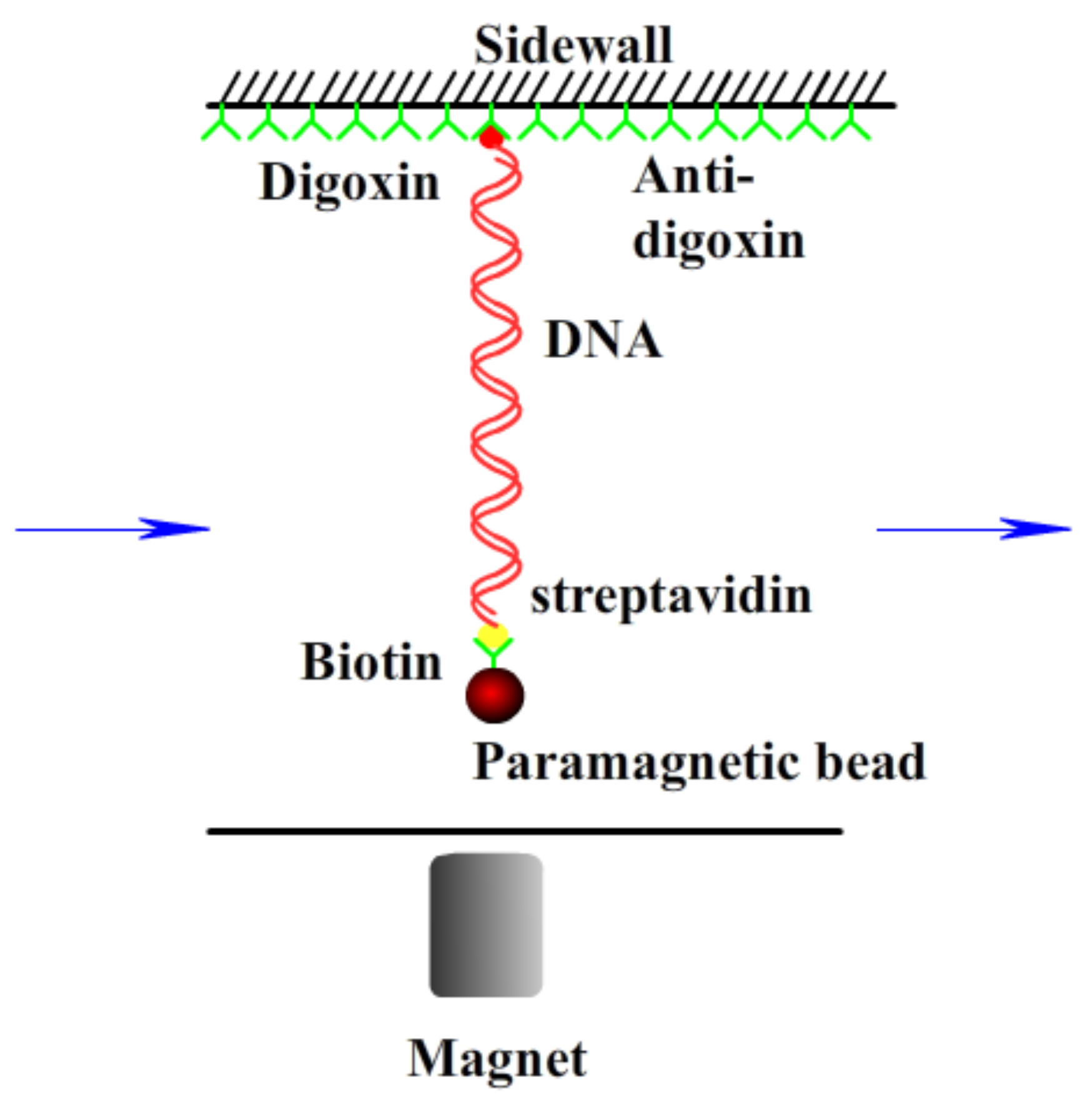
2.2. Electrophoretic Mobility and Size Measurement by Dynamic Light Scattering (DLS)
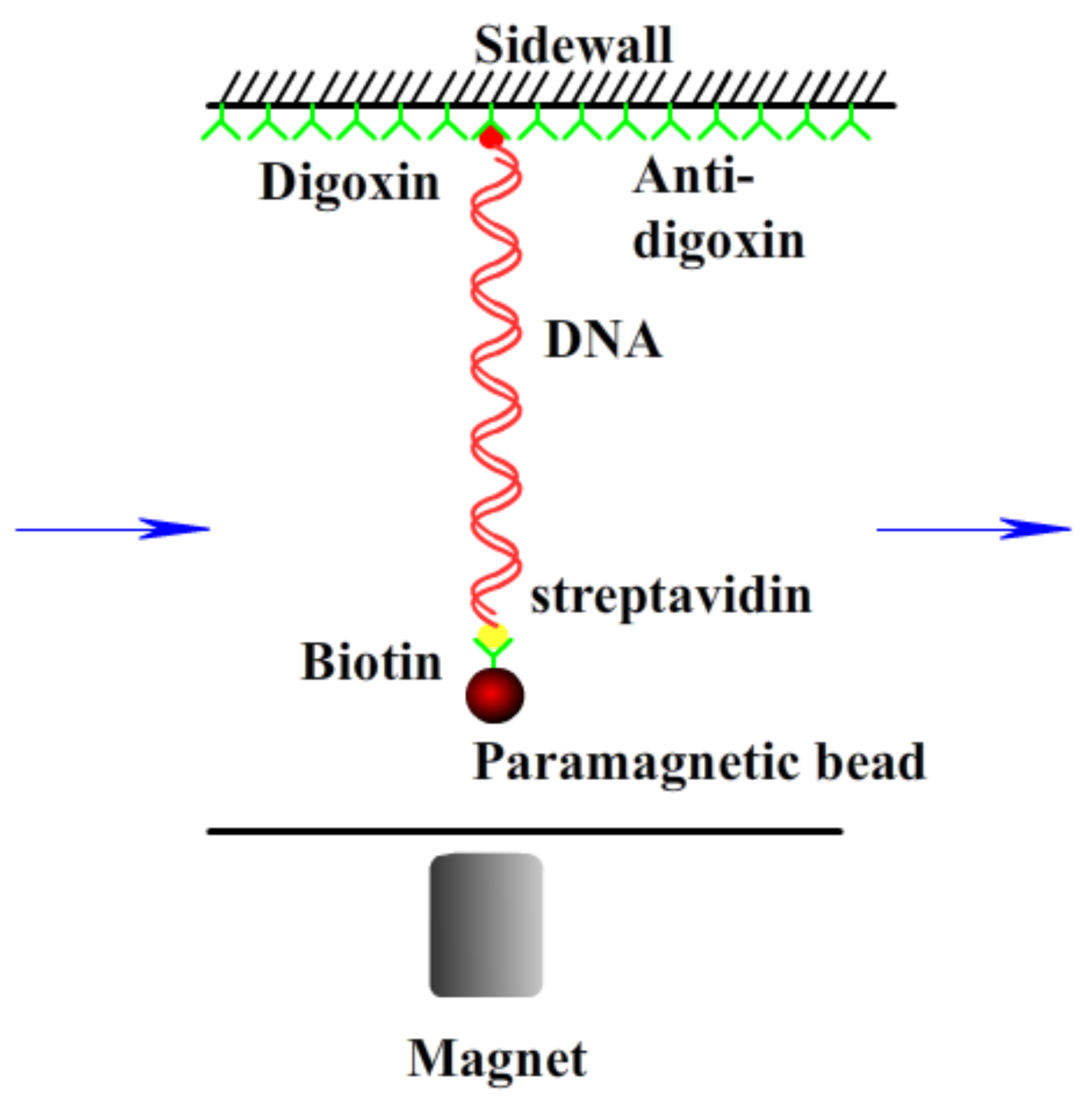
2.3. AFM Imaging and Magnetic Tweezers Tethering
3. Results and Discussion
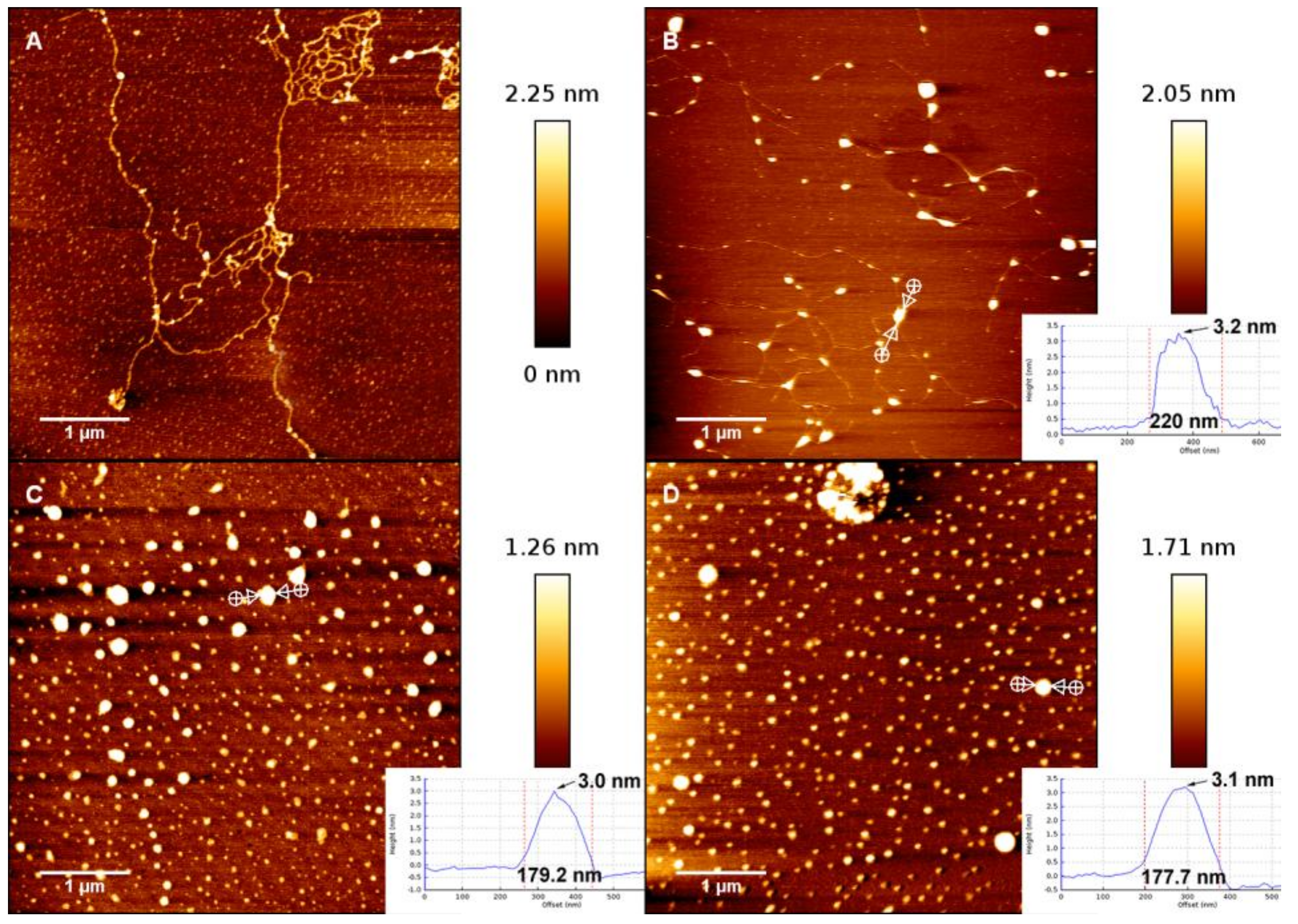
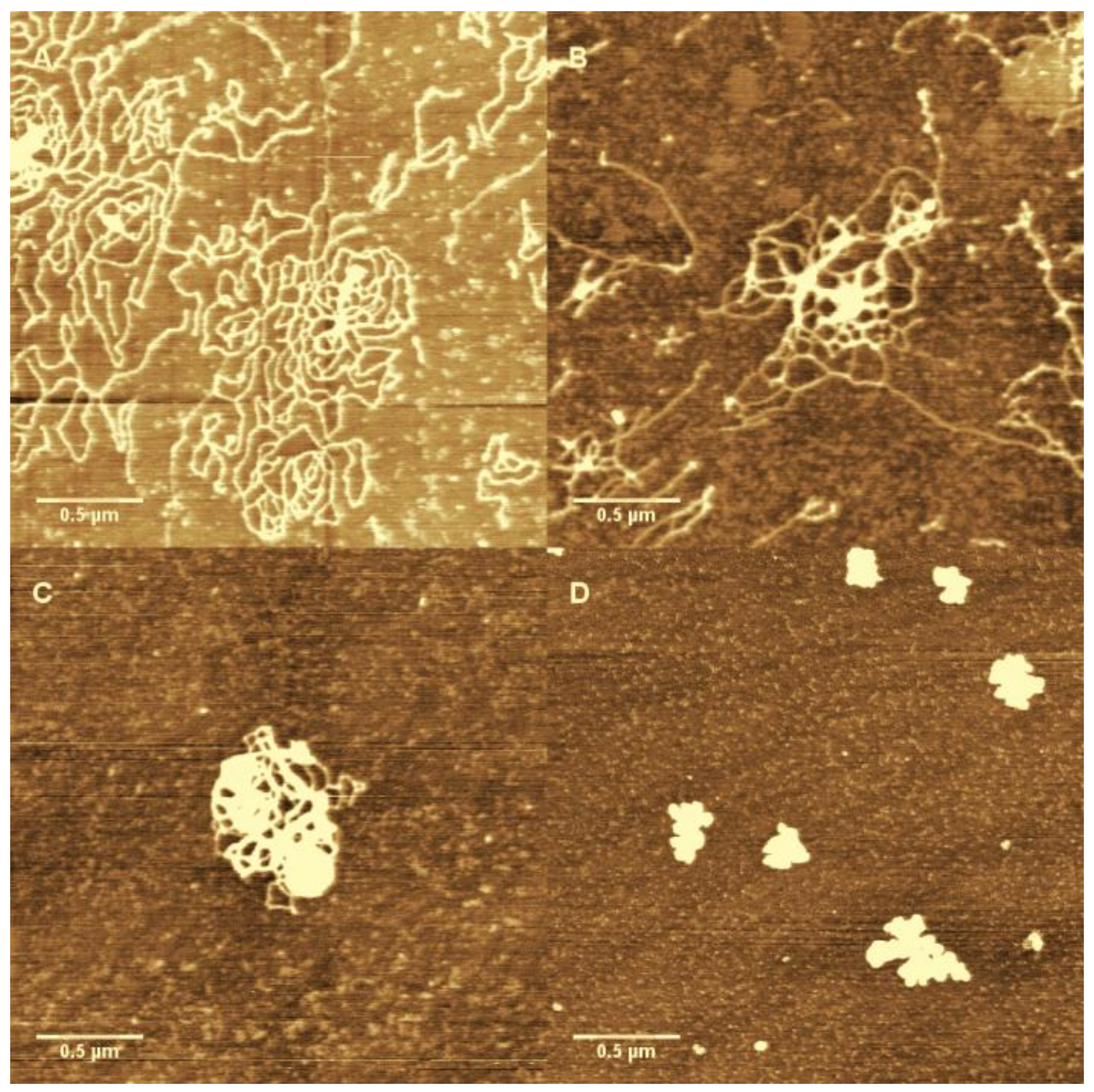
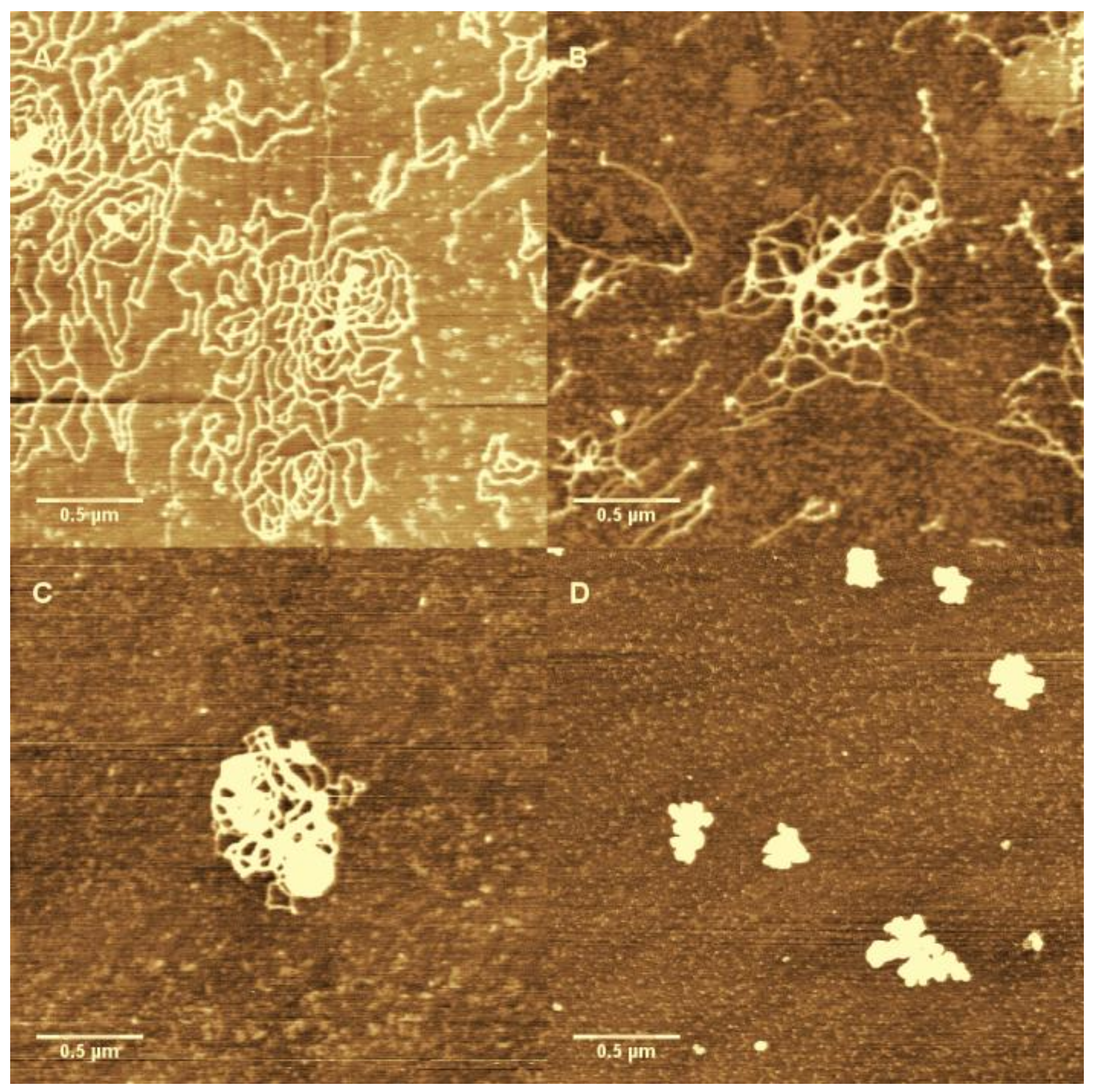
3.1. AFM Morphology of DNA
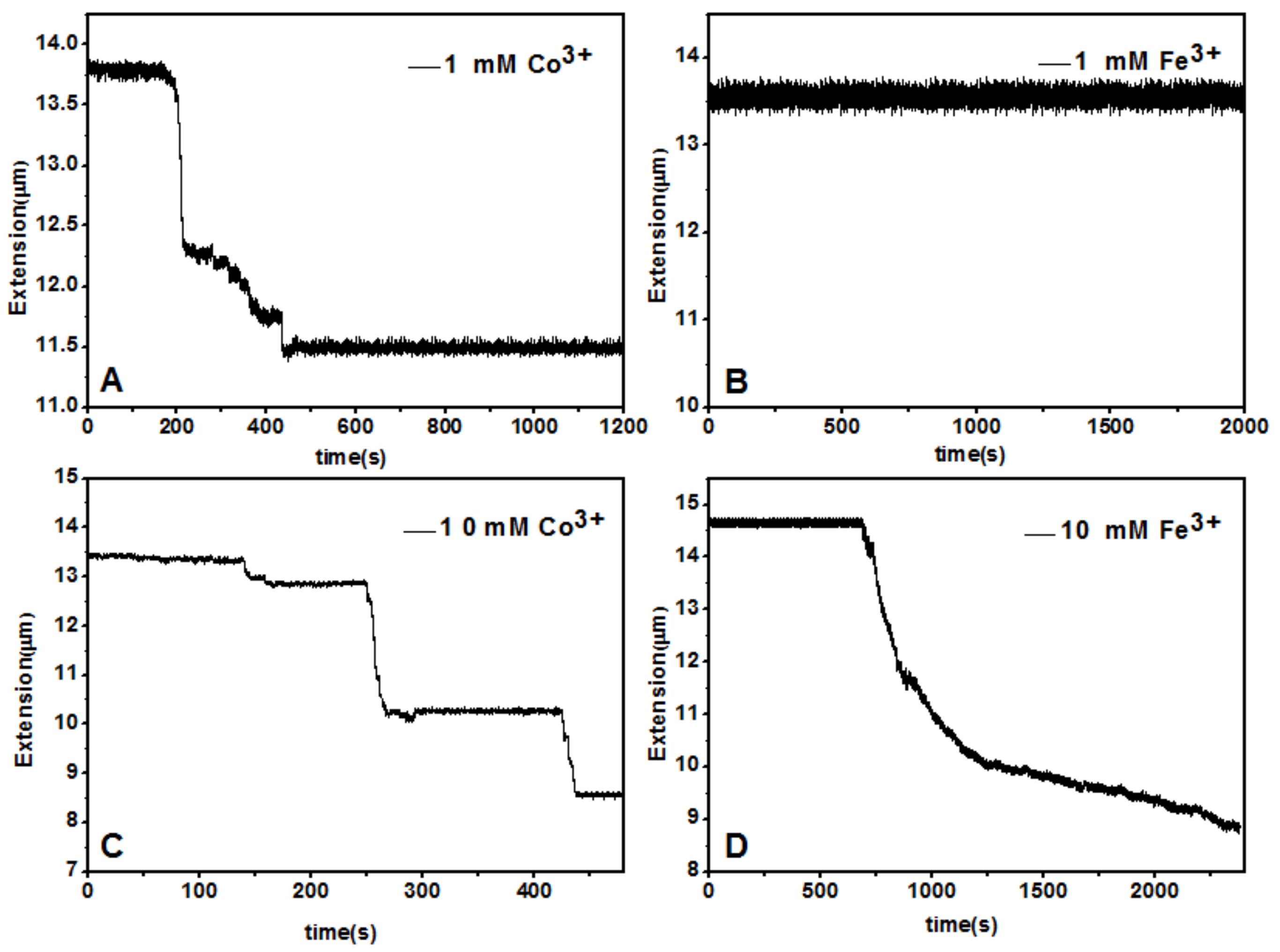
3.2. Force Spectroscopy of DNA Complexes
4. Conclusions
- (1)
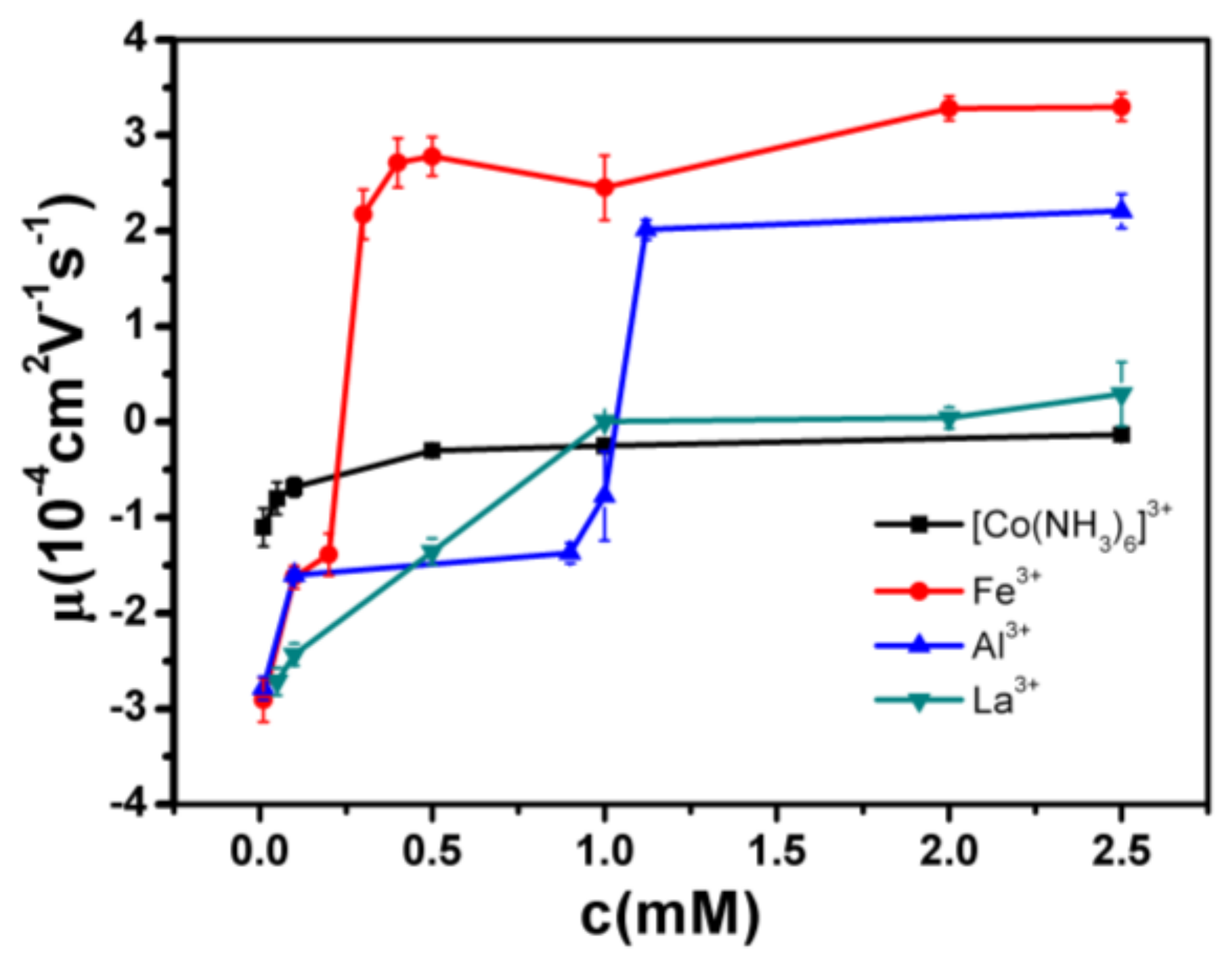
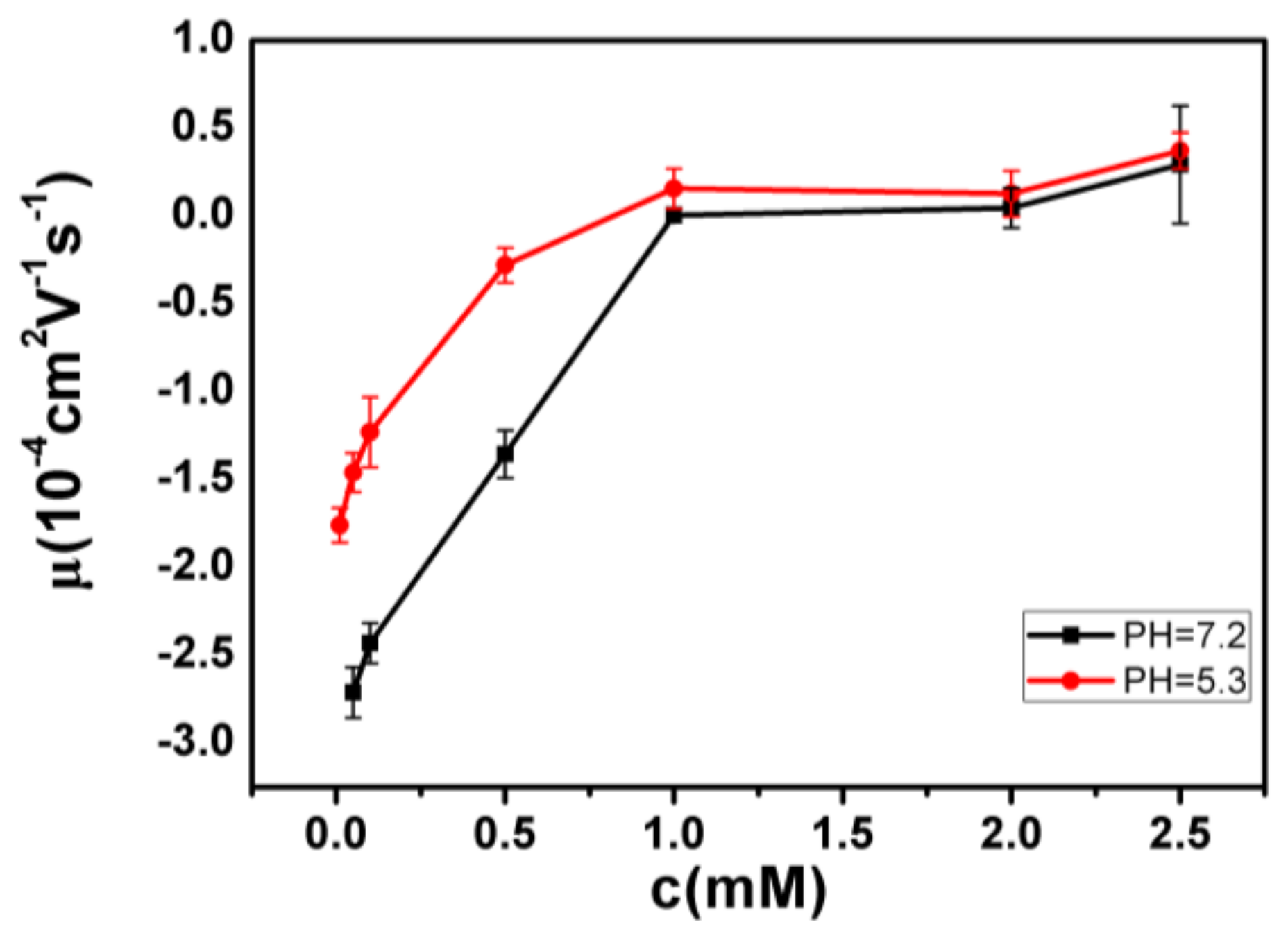
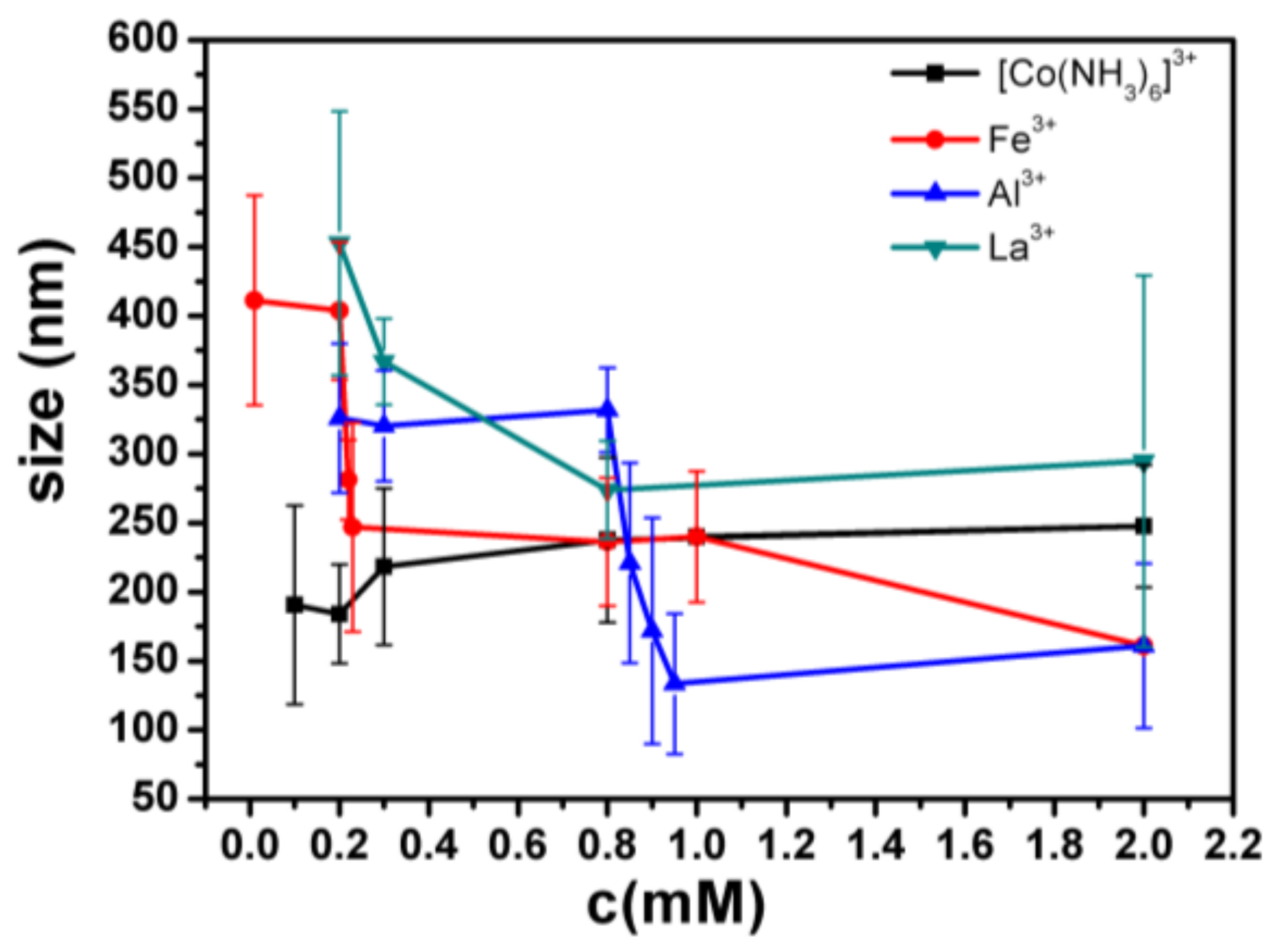
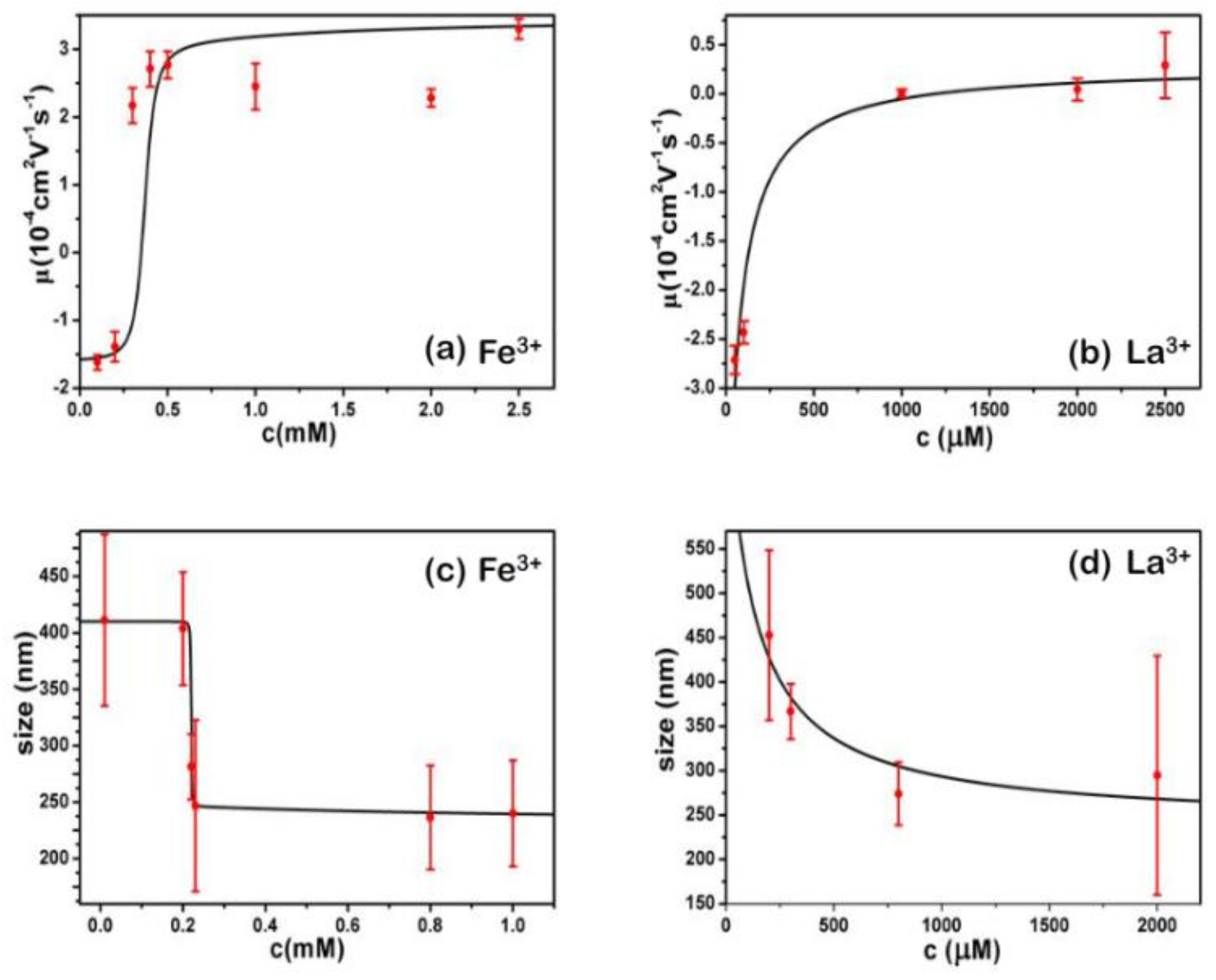
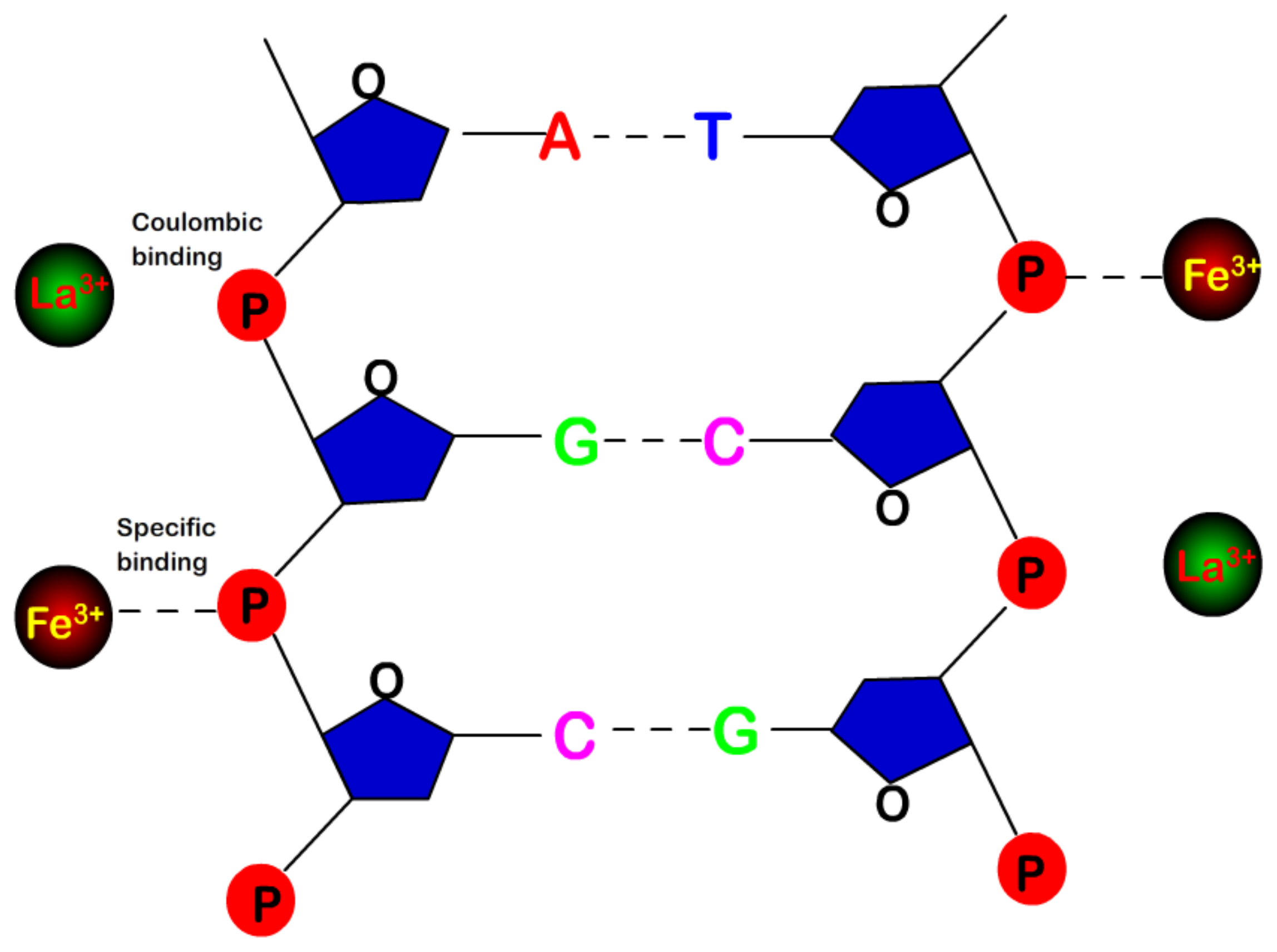
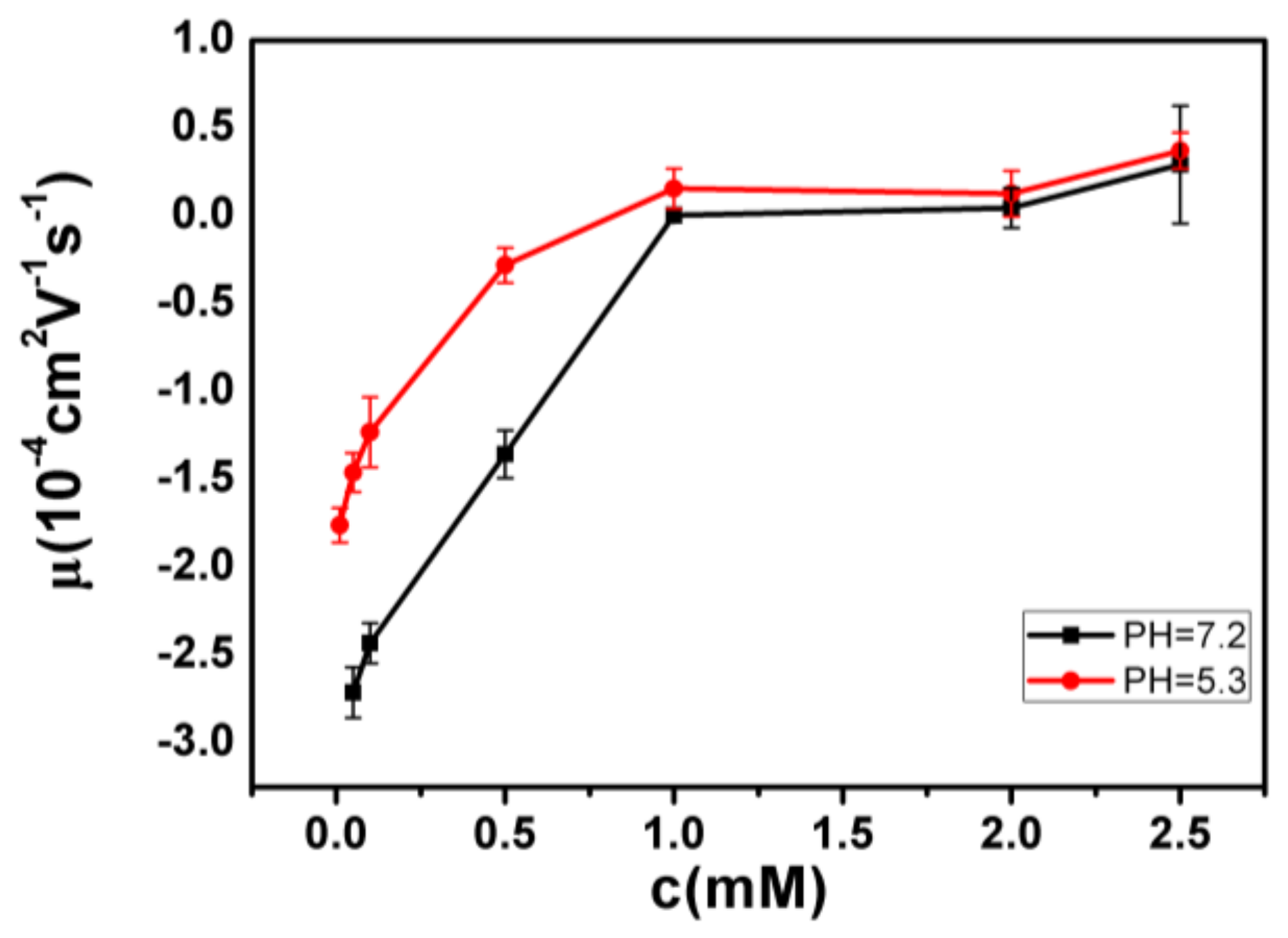
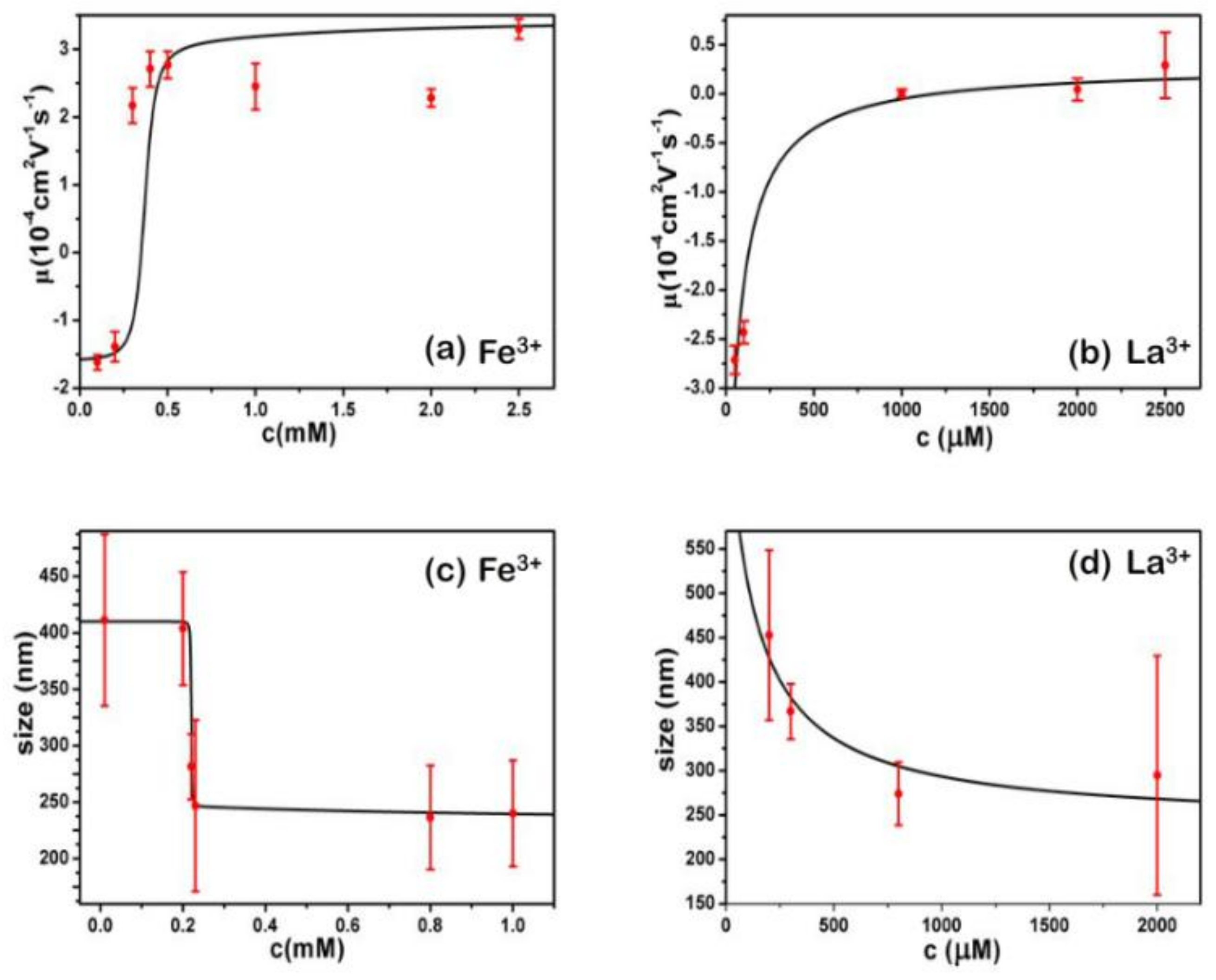
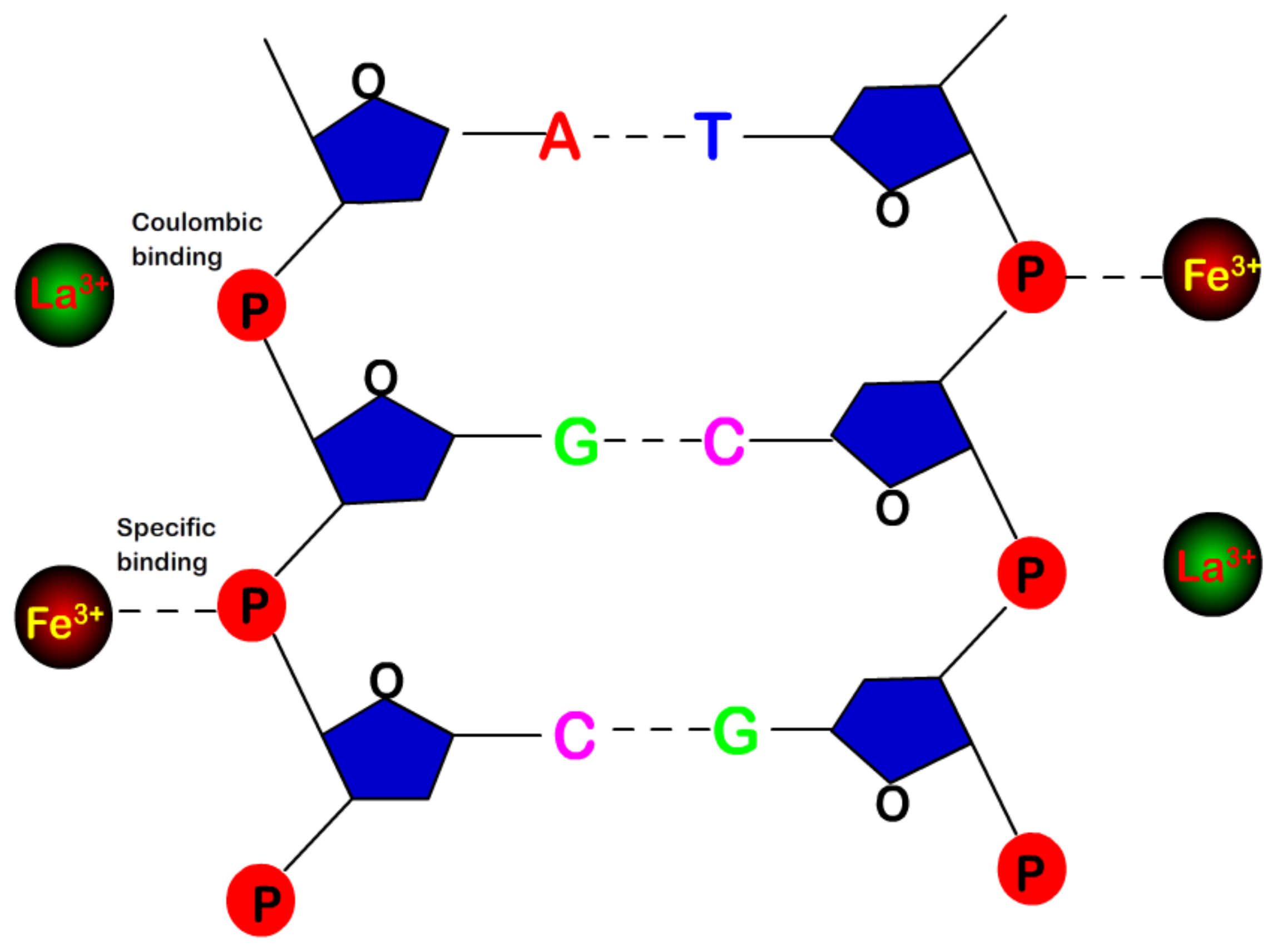
- Despite the fact that Fe3+ or Al3+, and La3+ or Co3+, have the same valence, they result in significantly different effects upon DNA in solution. Both kinds of trivalent counterions can lead to DNA compaction; the easily hydrolysed metal ions are able to invert the charge of DNA, while neutral La3+ and Co3+ are not.
- (2)
- The two kinds of trivalent counterions behave distinctly differently in the charge neutralization and compaction of DNA. La3+ or Co3+ provide reasonable representations of “classical” ions, whose interaction with DNA can be described by a simple two-state model in statistical mechanics. In contrast, the hydrolysed Fe3+ and Al3+ show ionic specific interactions with phosphate groups of DNA. We used a cooperative Zimm–Bragg model to explain the observed conformational and electrophoretic mobility transition of DNA.
- (3)
- DNA condensing force by hydrolysed Fe3+ and Al3+ is weaker than those by neutral La3+ and Co3+, and there are no stepwise jumps, but there is continuous shrinking in the pulling curves. Due to hydrolysis of Fe3+ and Al3+, pH variation not only regulates DNA charge but also plays a role in binding of the ions to DNA.
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Livolant, F. Ordered phases of DNA in vivo and in vitro. Phys. A Stat. Mech. Appl. 1991, 176, 117–137. [Google Scholar] [CrossRef]
- Thomas, T.J.; Tajmir-Riahi, H.A. Polyamine–DNA interactions and development of gene delivery vehicles. Amino Acids 2016, 48, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.; Klibanov, A.M. Non-viral gene therapy, polycation-mediated DNA delivery. Appl. Microbiol. Biot. 2003, 62, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Kanasty, R.L. Non-viral vectors for gene-based therapy. Nat. Rev. Genet. 2014, 15, 541–555. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Iwaki, T. Small Anion with Higher Valency Retards the Compaction of DNA in the Presence of Multivalent Cation. Biophys. J. 2009, 96, 1068–1075. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, R. Single Molecular Demonstration of Modulating Charge Inversion of DNA. Sci. Rep. 2016, 6, 38628. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Yang, G. The effect of pH on charge inversion and condensation of DNA. Soft Matter 2016, 12, 6669–6674. [Google Scholar] [CrossRef] [PubMed]
- Qiu, S.; Yang, G. The suppression and promotion of DNA charge inversion by mixing counterions. Soft Matter 2015, 11, 4099–4105. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, G. Ethanol induces condensation of single DNA molecules. Soft Matter 2011, 7, 4425–4434. [Google Scholar] [CrossRef]
- Wang, Y.-W.; Yang, G.-C. Modulation and control of DNA charge inversion. Chin. Phys. B 2017, 26, 7–11. [Google Scholar] [CrossRef]
- Xia, W.; Yang, G. DNA Compaction and Charge Inversion Induced by Organic Monovalent Ions. Polymers 2017, 9, 128. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, G. DNA condensations on mica surfaces induced collaboratively by alcohol and hexammine cobalt. Colloids Surf. B 2011, 83, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Mengarelli, V.; Auvray, L. Charge inversion, condensation and decondensation of DNA and polystyrene sulfonate by polyethylenimine. Eur. Phys. J. E 2011, 34, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Besteman, K.; Zevenbergen, M.A.G. Direct Observation of Charge Inversion by Multivalent Ions as a Universal Electrostatic Phenomenon. Phys. Rev. Lett. 2004, 93, 170802. [Google Scholar] [CrossRef] [PubMed]
- Hinterdorfer, P.; Dufrêne, Y.F. Detection and localization of single molecular recognition events using atomic force microscopy. Nat. Methods 2006, 3, 347. [Google Scholar] [CrossRef] [PubMed]
- Neuman, K.C.; Nagy, A. Single-molecule force spectroscopy, optical tweezers, magnetic tweezers and atomic force microscopy. Nat. Methods 2008, 5, 491–505. [Google Scholar] [CrossRef] [PubMed]
- Japaridze, A.; Benke, A. Influence of DNA Binding Dyes on Bare DNA Structure Studied with Atomic Force Microscopy. Macromolecules 2015, 48, 1860–1865. [Google Scholar] [CrossRef]
- Besteman, K.; Eijk, K.V. Charge inversion accompanies DNA condensation by multivalent ions. Nat. Phys. 2006, 3, 641–644. [Google Scholar] [CrossRef]
- Fu, W.B.; Wang, X.L. Compaction dynamics of single DNA molecules under tension. JACS 2006, 128, 15040. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Skoko, D. Near-field-magnetic-tweezer manipulation of single DNA molecules. Phys. Rev. E 2004, 70, 011905. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Anderson, N.A. Regulation of the electric charge in phosphatidic acid domains. J. Phys. Chem. B 2012, 116, 7213. [Google Scholar] [CrossRef] [PubMed]
- Mohanakrishnan, P.; Chignell, C.F. Ion-specific electrode study of copper binding to serum albumins. J. Pharm. Sci. 1982, 71, 1180–1182. [Google Scholar] [CrossRef] [PubMed]
- Li, G.Y.; Guan, R.L. DNA condensation induced by metal complexes. Coordination Chemistry Reviews. Coord. Chem. Rev. 2014, 281, 100–113. [Google Scholar] [CrossRef]
- Le Calvez, E.; Blaudez, D. Effect of cations on the dissociation of arachidic acid monolayers on water studied by polarization-modulated infrared reflection–absorption spectroscopy. Langmuir 2001, 17, 670–674. [Google Scholar] [CrossRef]
- Kundu, S.; Langevin, D. Fatty acid monolayer dissociation and collapse, Effect of pH and cations. Colloid Surf. A 2008, 325, 81–85. [Google Scholar] [CrossRef]
- Bu, W.; Flores, K. Preferential affinity of calcium ions to charged phosphatidic acid surface from a mixed calcium/barium solution, X-ray reflectivity and fluorescence studies. Langmuir 2008, 25, 1068–1073. [Google Scholar] [CrossRef] [PubMed]
- Pittler, J.; Bu, W. Charge inversion at minute electrolyte concentrations. Phys. Rev. Lett. 2006, 97, 046102. [Google Scholar] [CrossRef] [PubMed]
- Lam, D.M.; Furrer, R. Interplay of pH and binding of multivalent metal ions, charge inversion and reentrant condensation in protein solutions. J. Phys. Chem. B 2013, 117, 5777. [Google Scholar] [CrossRef]
- And, Y.Y.; Yoshikawa, K. Higher Order Structure of DNA Controlled by the Redox State of Fe2+/Fe3+. JACS 1997, 119, 10573–10578. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, H. Iron ion and iron hydroxide adsorption to charge-neutral phosphatidylcholine templates. Langmuir 2016, 32, 7664–7670. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Park, R.Y. Ionic specificity in pH regulated charged interfaces, Fe3+ versus La3+. Langmuir 2011, 27, 11917–11924. [Google Scholar] [CrossRef] [PubMed]
- Aust, S.D.; Morehouse, L.A. Role of metals in oxygen radical reactions. J. Mech. Med. Biol. 1985, 1, 3–25. [Google Scholar] [CrossRef]
- Meneghini, R. Genotoxicity of active oxygen species in mammalian cells. Mutat. Res. Rev. Genet. Toxicol. 1988, 195, 215–230. [Google Scholar] [CrossRef]
- Gosse, C. Magnetic tweezers: Micromanipulation and force measurement at the molecular level. Biophys. J. 2002, 82, 3314. [Google Scholar] [CrossRef]
- Strick, T.R.; Dessinges, M.N. Stretching of macromolecules and proteins. Rep. Prog. 2003, 66, 1. [Google Scholar] [CrossRef]
- Buyukdagli, S.; Blossey, R. Correlation-induced DNA adsorption on like-charged membranes. Phys. Rev. Lett. 2016, 94, 042502. [Google Scholar] [CrossRef] [PubMed]
- Zimm, B.H.; Bragg, J. Theory of the phase transition between helix and random coil in polypeptide chains. J. Chem. Phys. 1959, 31, 526–535. [Google Scholar] [CrossRef]
- Phillips, R.; Kondev, J. Physical Biology of the Cell; Garland Science: New York, NY, USA, 2012; pp. 44–45. ISBN 978-0815344506. [Google Scholar]
- Trotta, A.; Paulsen, A.B. The dynamics of 57Fe nuclei in FeIII—DNA condensates. J. Inorg. Biochem. 2002, 88, 14–18. [Google Scholar] [CrossRef]
- Kasyanenko, N.; Arikainen, N. Investigation of DNA complexes with iron ions in solution. Biophys. Chem. 1998, 70, 93–100. [Google Scholar] [CrossRef]
- Wang, W.; Park, R.Y. Ion-specific induced charges at aqueous soft interfaces. Phys. Rev. Lett. 2011, 106, 056102. [Google Scholar] [CrossRef] [PubMed]
- Toshio, I.; Kenichi, Y. Cooperativity vs. Phase Transition in a Giant Single DNA Molecule. JACS 2000, 122, 9891–9896. [Google Scholar] [CrossRef]
- Ouameur, A.A.; Arakawa, H. A Comparative study of Fe(II) and Fe(III) interactions with DNA duplex, major and minor grooves bindings. DNA Cell Biol. 2005, 24, 394. [Google Scholar] [CrossRef] [PubMed]
- Mawhinney, M.T.; Liu, R. CTCF-Induced Circular DNA Complexes Observed by Atomic Force Microscopy. J. Mol. Biol. 2018. [Google Scholar] [CrossRef] [PubMed]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, Z.; Wang, Y.; Li, S.; Yang, G. DNA Phase Transition in Charge Neutralization and Comformation Induced by Trivalent-Hydrolysed Metal Ions. Polymers 2018, 10, 394. https://doi.org/10.3390/polym10040394
Luo Z, Wang Y, Li S, Yang G. DNA Phase Transition in Charge Neutralization and Comformation Induced by Trivalent-Hydrolysed Metal Ions. Polymers. 2018; 10(4):394. https://doi.org/10.3390/polym10040394
Chicago/Turabian StyleLuo, Zhaoxu, Yanwei Wang, Shuhang Li, and Guangcan Yang. 2018. "DNA Phase Transition in Charge Neutralization and Comformation Induced by Trivalent-Hydrolysed Metal Ions" Polymers 10, no. 4: 394. https://doi.org/10.3390/polym10040394
APA StyleLuo, Z., Wang, Y., Li, S., & Yang, G. (2018). DNA Phase Transition in Charge Neutralization and Comformation Induced by Trivalent-Hydrolysed Metal Ions. Polymers, 10(4), 394. https://doi.org/10.3390/polym10040394