
Insights into Crystallization of Neuronal Nicotinic α4β2 Receptor in Polarized Lipid Matrices

 ,
,
Abstract
1. Introduction
2. Materials and Methods
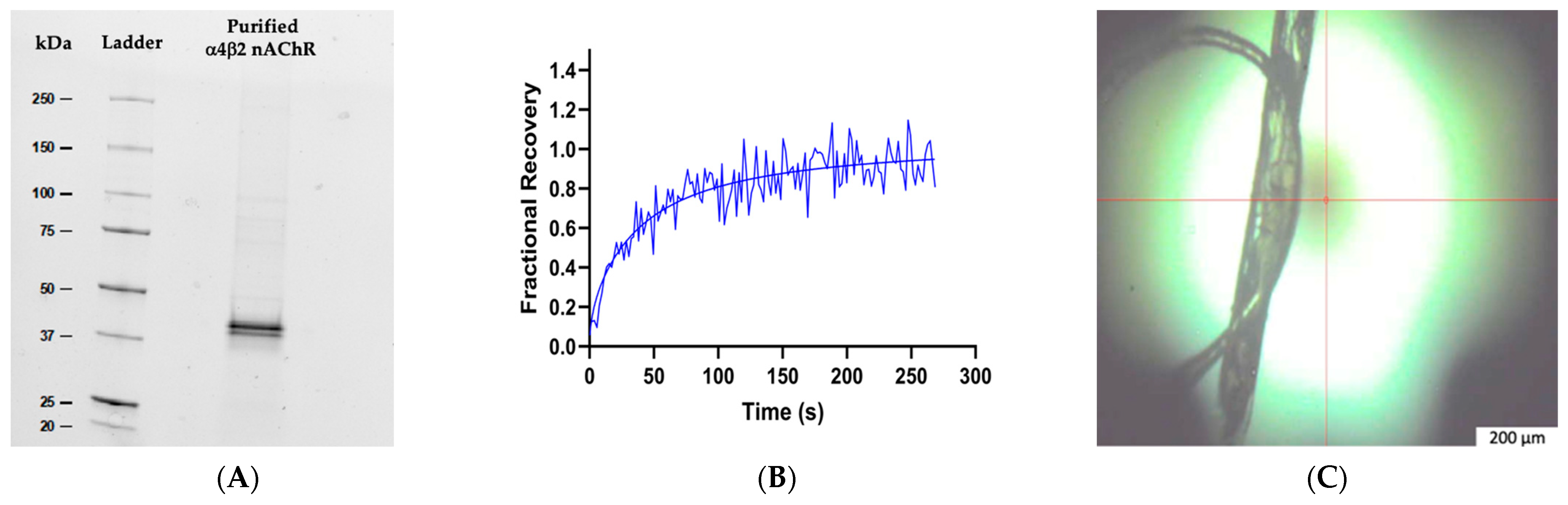
2.1. Expression and Purification of Human α4β2 nAChR
2.2. In Meso Crystallization with the RPM@LMx Device and X-ray Diffraction
2.3. Lipidic Cubic Phase Fluorescence Recovery after Photobleaching (LCP–FRAP)
2.4. Analysis of Phospholipid Molecular Species by Ultra-Performance Liquid Chromatography (UPLC) Coupled to Electrospray Ionization Tandem Mass Spectrometry (ESI-MS/MS)
3. Results
3.1. Lipid Analysis of Purified Human α4β2 nAChR-DCs
3.2. α4β2 nAChR-DC Diffusion in LCP
3.3. Polarized In Meso Crystallization Screening of Human α4β2 nAChR
3.4. Synchrotron X-ray Diffraction
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Delgado-Vélez, M.; Quesada, O.; Villalobos-Santos, J.C.; Maldonado-Hernández, R.; Asmar-Rovira, G.; Stevens, R.C.; Lasalde-Dominicci, J.A. Pursuing high-resolution structures of nicotinic acetylcholine receptors: Lessons learned from five decades. Molecules 2021, 26, 5753. [Google Scholar] [CrossRef] [PubMed]
- Changeux, J.P. Discovery of the First Neurotransmitter Receptor: The Acetylcholine Nicotinic Receptor. Biomolecules 2020, 10, 547. [Google Scholar] [CrossRef]
- Delgado-Vélez, M.; Báez-Pagán, C.A.; Gerena, Y.; Quesada, O.; Santiago-Pérez, L.I.; Capó-Vélez, C.M.; Wojna, V.; Meléndez, L.; León-Rivera, R.; Silva, W.; et al. The α 7-nicotinic receptor is upregulated in immune cells from HIV-seropositive women: Consequences to the cholinergic anti-in flammatory response. Clin. Transl. Immunol. 2015, 4, e53. [Google Scholar] [CrossRef] [PubMed]
- Hurst, R.; Rollema, H.; Bertrand, D. Nicotinic acetylcholine receptors: From basic science to therapeutics. Pharmacol. Ther. 2013, 137, 22–54. [Google Scholar] [CrossRef]
- Polosa, R.B.N.L. Treatment of nicotine addiction: Present therapeutic options and pipeline developments. Trends Pharmacol Sci. 2011, 32, 281–289. [Google Scholar] [CrossRef]
- Quik, M.; O’Leary, K.; Tanner, C.M. Nicotine and Parkinson’s disease; implications for therapy. Physiol. Behav. 2008, 23, 1641–1652. [Google Scholar] [CrossRef] [PubMed]
- Cooper, S.Y.; Akers, A.T.; Journigan, V.B.; Henderson, B.J. Novel putative positive modulators of α4β2 nachrs potentiate nicotine reward-related behavior. Molecules 2021, 26, 4793. [Google Scholar] [CrossRef]
- Buisson, B.; Bertrand, D. Open-channel blockers at the human α4β2 neuronal nicotinic acetylcholine receptor. Mol. Pharmacol. 1998, 53, 555–563. [Google Scholar] [CrossRef]
- Vallés, A.S.; Barrantes, F.J. Nicotinic acetylcholine receptor dysfunction in addiction and in some neurodegenerative and neuropsychiatric diseases. Cells 2023, 12, 2051. [Google Scholar] [CrossRef]
- Walsh, R.M., Jr.; Roh, S.H.; Gharpure, A.; Morales-Perez, C.L.; Teng, J.; Hibbs, R.E. Structural principles of distinct assemblies of the human α 4 β 2 nicotinic receptor. Nature 2018, 557, 261–265. [Google Scholar] [CrossRef]
- Gharpure, A.; Teng, J.; Zhuang, Y.; Noviello, C.M.; Walsh, R.M., Jr.; Cabuco, R.; Howard, R.J.; Zaveri, N.T.; Lindahl, E.; Hibbs, R.E. Agonist Selectivity and Ion Permeation in the α3β4 Ganglionic Nicotinic Receptor. Neuron 2019, 104, 501–511.e6. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Teng, J.; Worrell, B.T.; Noviello, C.M.; Lee, M.; Karlin, A.; Stowell, M.H.B.; Hibbs, R.E. Structure of the Native Muscle-type Nicotinic Receptor and Inhibition by Snake Venom Toxins. Neuron 2020, 106, 952–962.e5. [Google Scholar] [CrossRef]
- Rahman, M.; Basta, T.; Teng, J.; Lee, M.; Worrell, B.T.; Stowell, M.H.B.; Hibbs, R.E. Structural mechanism of muscle nicotinic receptor desensitization and block by curare. Nat. Struct. Mol. Biol. 2022, 4, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Noviello, C.M.; Gharpure, A.; Mukhtasimova, N.; Cabuco, R.; Baxter, L.; Borek, D.; Sine, S.M.; Hibbs, R.E. Structure and gating mechanism of the α7 nicotinic acetylcholine receptor. Cell 2021, 184, 2121–2134. [Google Scholar] [CrossRef]
- Zhang, D.; Liu, Y.; Zaidi, S.A.; Xu, L.; Zhan, Y.; Chen, A.; Guo, J.; Huang, X.; Roth, B.L.; Katritch, V.; et al. Structural insight into angiotensin receptor signaling modulation by balanced and biased agonists. EMBO J. 2023, 42, e112940. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y. Structural insight into apelin receptor-G protein stoichiometry. Nat. Struct. Mol. Biol. 2022, 29, 688–697. [Google Scholar]
- Mazzaferro, S.; Kang, G.; Natarajan, K.; Hibbs, R.E.; Sine, S.M. Structural bases for Stoichiometry-Selective Calcium Potentiation of a Neuronal Nicotinic Receptor. Br. J. Pharmacol. 2024, 181, 1973–1992. [Google Scholar] [CrossRef]
- Morales-Perez, C.L.; Noviello, C.M.; Hibbs, R.E. X-ray structure of the human α4β2 nicotinic receptor. Nature 2016, 538, 411–415. [Google Scholar] [CrossRef]
- Padilla-Morales, L.F.; Colón-Sáez, J.O.; González-Nieves, J.E.; Quesada-González, O.; Lasalde-Dominicci, J.A. Functionality and stability data of detergent purified nAChR from Torpedo using lipidic matrixes and macroscopic electrophysiology. Data Brief 2016, 6, 433–437. [Google Scholar] [CrossRef]
- Padilla-Morales, L.F.; Colón-Sáez, J.O.; González-Nieves, J.E.; Quesada-González, O.; Lasalde-Dominicci, J.A. Assessment of the Functionality and Stability of Detergent Purified nAChR from Torpedo using Lipidic Matrixes and Macroscopic Electrophysiology. Biochim. Biophys. Acta (BBA)-Biomembr. 2017, 1858, 47–56. [Google Scholar] [CrossRef]
- Caffrey, M.; Cherezov, V. Crystallizing Membrane Proteins Using Lipidic Mesophases. Nat. Protoc. 2010, 4, 706–731. [Google Scholar] [CrossRef] [PubMed]
- Luecke, H.; Schobert, B.; Richter, H.T.; Cartailler, J.P.; Lanyi, J.K. Structure of bacteriorhodopsin at 1.55 Å resolution. J. Mol. Biol. 1999, 291, 899–911. [Google Scholar] [CrossRef] [PubMed]
- Caffrey, M. A comprehensive review of the lipid cubic phase or in meso method for crystallizing membrane and soluble proteins and complexes. Acta Crystallogr. Sect. FStructural Biol. Commun. 2015, 71, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Cherezov, V.; Rosenbaum, D.M.; Hanson, M.A.; Rasmussen, S.G.; Thian, F.S.; Kobilka, T.S.; Choi, H.J.; Kuhn, P.; Weis, W.I.; Kobilka, B.K.; et al. High-Resolution Crystal Structure of an Engineered Human β2-adrenergic G protein–coupled receptor. Science 2007, 318, 1258–1265. [Google Scholar] [CrossRef]
- Martynowycz, M.W.; Shiriaeva, A.; Ge, X.; Hattne, J.; Nannenga, B.L.; Cherezov, V.; Gonen, T. MicroED structure of the human adenosine receptor determined from a single nanocrystal in LCP. Proc. Natl. Acad. Sci. USA 2021, 118, e2106041118. [Google Scholar] [CrossRef]
- Michaelian, N.; Sadybekov, A.; Besserer-Offroy, É.; Han, G.W.; Krishnamurthy, H.; Zamlynny, B.A.; Fradera, X.; Siliphaivanh, P.; Presland, J.; Spencer, K.B.; et al. Structural insights on ligand recognition at the human leukotriene B4 receptor 1. Nat. Commun. 2021, 12, 2971. [Google Scholar] [CrossRef]
- Morales-Perez, C.L.; Noviello, C.M.; Hibbs, R.E. Manipulation of Subunit Stoichiometry in Heteromeric Membrane Proteins Resource Manipulation of Subunit Stoichiometry in Heteromeric Membrane Proteins. Struct. Des. 2016, 24, 797–805. [Google Scholar] [CrossRef]
- Liu, W.; Cherezov, V. Crystallization of membrane proteins in lipidic mesophases. J. Vis. Exp. 2011, 49. [Google Scholar] [CrossRef]
- Quesada, O.; González-Freire, J.E.; Colón, J.; Maldonado-Hernández, R.; González-Freire, C.; Acevedo-Cintrón, J.; Rosado-Millán, I.D.; Lasalde-Dominicci, J.A. Assessment of Purity, Functionality, Stability, and Lipid Composition of Cyclofos-nAChR-Detergent Complexes from Torpedo californica Using Lipid Matrix and Macroscopic Electrophysiology. J. Mem. Biol. 2023, 256, 271–285. [Google Scholar] [CrossRef]
- Quesada, O.; Freire, C.G.; Ferrer, M.C.; -Sáez, J.O.C.; Fernández-García, E.; Mercado, J.; Dávila, A.; Morales, R.; Lasalde-Dominicci, J.A. Uncovering the lipidic basis for the preparation of functional nicotinic acetylcholine receptor detergent complexes for structural studies. Sci. Rep. 2016, 6, 32766. [Google Scholar] [CrossRef]
- Symons, J.L.; Cho, K.J.; Chang, J.T.; Du, G.; Waxham, M.N.; Hancock, J.F.; Levental, I.; Levental, K.R. Lipidomic atlas of mammalian cell membranes reveals hierarchical variation induced by culture conditions, subcellular membranes, and cell lineages. Soft Matter 2021, 17, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Palsdottir, H.; Hunte, C. Lipids in membrane protein structures. Biochim. Biophys. Acta 2004, 1666, 2–18. [Google Scholar] [CrossRef] [PubMed]
- McPherson, A.; Gavira, J.A. Introduction to protein crystallization. Acta Cryst. 2013, F70, 2–20. [Google Scholar]
- Shoemaker, S.C.; Ando, N. X-rays in the Cryo-EM era: Structural Biology’s dynamic future. Biochemistry 2019, 57, 277–285. [Google Scholar] [CrossRef]
- Barrantes, F.J. Structure and function meet at the nicotinic acetylcholine receptor-lipid interface. Pharmacol. Res. 2023, 190, 106729. [Google Scholar] [CrossRef] [PubMed]
- Barrantes, F.J. Modulation of a rapid neurotransmitter receptor-ion channel by membrane lipids. Front. Cell Dev. Biol. 2024, 11, 1328875. [Google Scholar] [CrossRef]
- Dawaliby, R.; Trubbia, C.; Delporte, C.; Noyon, C.; Ruysschaert, J.M.; Antwerpen, P.V.; Govaerts, C. Phosphatidylethanolamine is a key regulator of membrane fluidity in eukaryotic cells. J. Biol. Chem. 2016, 291, 3658–3668. [Google Scholar] [CrossRef]
- Hejazi, S.; Pahlavandanzadeh, H.; Elliott, J.A.W. Thermodynamic investigation of the effect of electric field on solid-liquid equilibrium. J. Phys. Chem. B 2021, 125, 1271–1281. [Google Scholar] [CrossRef]
- Nanev, C.N. Recent Insights into the Crystallization Process; Protein Crystal Nucleation and Growth Peculiarities; Processes in the Presence of Electric Fields. Crystals 2017, 7, 310. [Google Scholar] [CrossRef]
- Van’t Hag, L.; Knoblich, K.; Seabrook, S.A.; Kirby, N.M.; Mudie, S.T.; Lau, D.; Li, X.; Gras, S.L.; Mulet, X.; Call, M.E.; et al. Exploring the in meso crystallization mechanism by characterizing the lipid mesophase microenvironment during the growth of single transmembrane alpha-helical peptide crystals. Phil. Trans. R. Soc. A 2016, 374, 20150125. [Google Scholar] [CrossRef]
- Al-Haq, M.I.; Lebrasseur, E.; Tsuchiya, H.; Torri, T. Protein crystallization under an electric field. Cryst. Rev. 2007, 13, 29–64. [Google Scholar] [CrossRef]
- Ogata, M.; Aoki, D.; Suzuki, M.; Tanaka, D.; Wakamatsu, T. The role of an applied electric field in protein crystallization at low temperature. Jpn. J. Appl. Phys. 2019, 58, 110903. [Google Scholar] [CrossRef]
- Kan, K. The effect of fatty acids, ionic strength, and electric fields on the microscopic dynamics of BSA aggregates. Front. Phys. 2023, 11, 1282099. [Google Scholar]
- Yager, P.; Price, R.R.; Schnur, J.M.; Schoen, P.E.; Singh, A.; Rhodes, D.G. The mechanism of formation of lipid tubules from liposomes. Chem. Phys. Lipids 1988, 46, 171–179. [Google Scholar] [CrossRef]
- Stachowiak, J.C.; Hayden, C.C.; Sasaki, D.Y. Steric confinement of proteins on lipid membranes can drive curvature and tubulation. Proc. Natl. Acad. Sci. USA 2010, 107, 7781–7786. [Google Scholar] [CrossRef]
- Pebay-Peyroula, E.; Rummel, G.; Rosenbusch, J.P.; Landau, E.M. X-ray structure of bacteriorhodopsin at 2.5 angstroms from microcrystals grown in lipidic cubic phases. Science 1997, 277, 1676–1681. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Molecular Species | m/z (M + H)+ | Mean Peak Area |
|---|---|---|
| LPC 16:0 | 496.3398 | 17,941,873.71 |
| Cer m22:0 | 338.3417 | 12,200,000.00 |
| TG P-23:6/21:1 | 721.5766 | 4,509,407.999 |
| LPC 17:0 | 554.3463 | 3,323,174.888 |
| LPMt 16:0 | 423.2517 | 1,824,723.222 |
| LPC (PC 16:0) | 518.3217 | 1,595,348.877 |
| DAP O-11:0/O-5:0 | 391.2843 | 922,029.400 |
| DAP O-7:0/O-7:0 | 363.2529 | 769,424.966 |
| SM d34:1 | 703.5749 | 480,092.577 |
| LPC 18:0 | 524.3711 | 266,859.633 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villalobos-Santos, J.C.; Carrasquillo-Rivera, M.; Rodríguez-Cordero, J.A.; Quesada, O.; Lasalde-Dominicci, J.A. Insights into Crystallization of Neuronal Nicotinic α4β2 Receptor in Polarized Lipid Matrices. Crystals 2024, 14, 889. https://doi.org/10.3390/cryst14100889
Villalobos-Santos JC, Carrasquillo-Rivera M, Rodríguez-Cordero JA, Quesada O, Lasalde-Dominicci JA. Insights into Crystallization of Neuronal Nicotinic α4β2 Receptor in Polarized Lipid Matrices. Crystals. 2024; 14(10):889. https://doi.org/10.3390/cryst14100889
Chicago/Turabian StyleVillalobos-Santos, Juan C., Mallerie Carrasquillo-Rivera, Josué A. Rodríguez-Cordero, Orestes Quesada, and José Antonio Lasalde-Dominicci. 2024. "Insights into Crystallization of Neuronal Nicotinic α4β2 Receptor in Polarized Lipid Matrices" Crystals 14, no. 10: 889. https://doi.org/10.3390/cryst14100889
APA StyleVillalobos-Santos, J. C., Carrasquillo-Rivera, M., Rodríguez-Cordero, J. A., Quesada, O., & Lasalde-Dominicci, J. A. (2024). Insights into Crystallization of Neuronal Nicotinic α4β2 Receptor in Polarized Lipid Matrices. Crystals, 14(10), 889. https://doi.org/10.3390/cryst14100889