Structural Biology of Calcium Phosphate Nanoclusters Sequestered by Phosphoproteins




Abstract
1. Introduction
2. Role of IDPs in the Control of Biocalcification
2.1. Secreted Calcium- or Calcium Phosphate-Binding Proteins
2.2. Calcium Phosphate-Binding Short Linear Motifs (CaP-SLiMs)
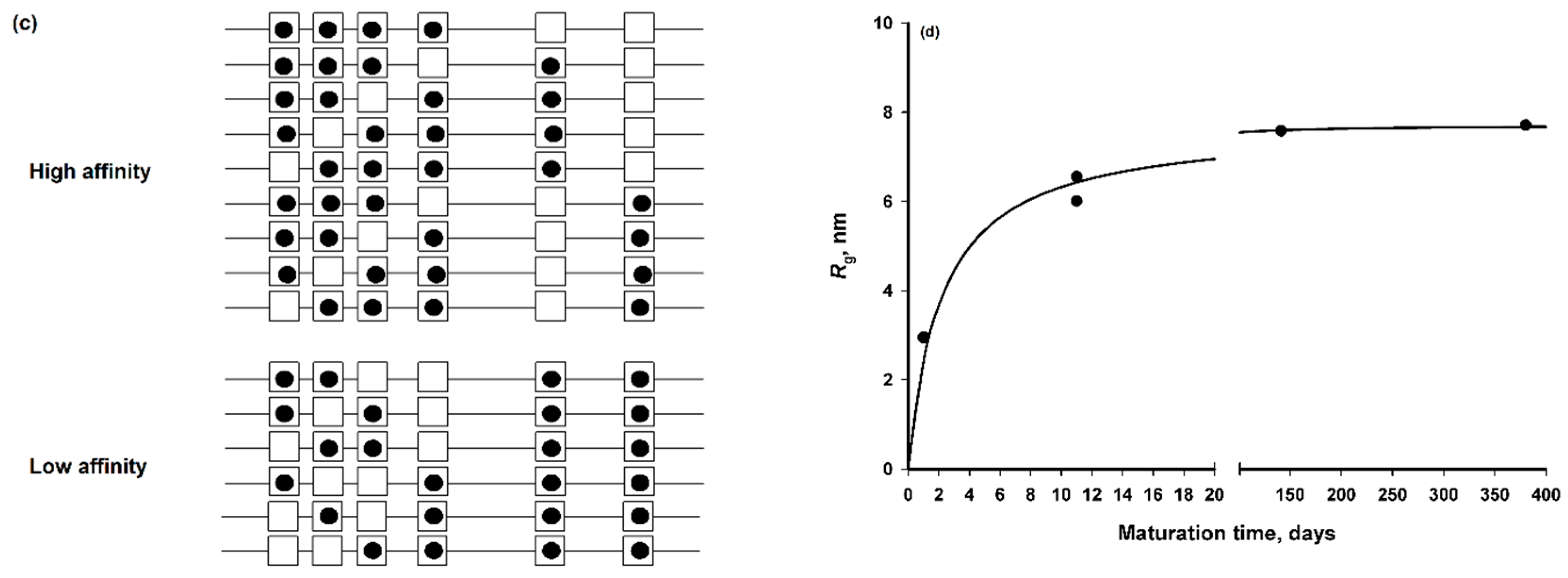
2.3. Conformational Flexibility and Adaptability
3. Basic Science of Amorphous Calcium Phosphate Sequestration by Phosphoproteins
3.1. Ostwald Rule of Stages and Non-Classical Nucleation of Calcium Phosphate
3.2. Effect of Phosphopeptides
3.2.1. Nucleation Versus Inhibition
3.2.2. Kinetic Stability of Complexes in Metastable Solutions
3.2.3. Thermodynamically Stable Complexes
3.2.4. Destabilization of Thermodynamically Stable Solutions
3.3. Ion Activity Product in the Continuous Phase of Solutions Containing Calcium Phosphate Nanocluster Complexes
3.4. Calculation of the Partition of Salts in a Biofluid Containing CaP Nanocluster Complexes
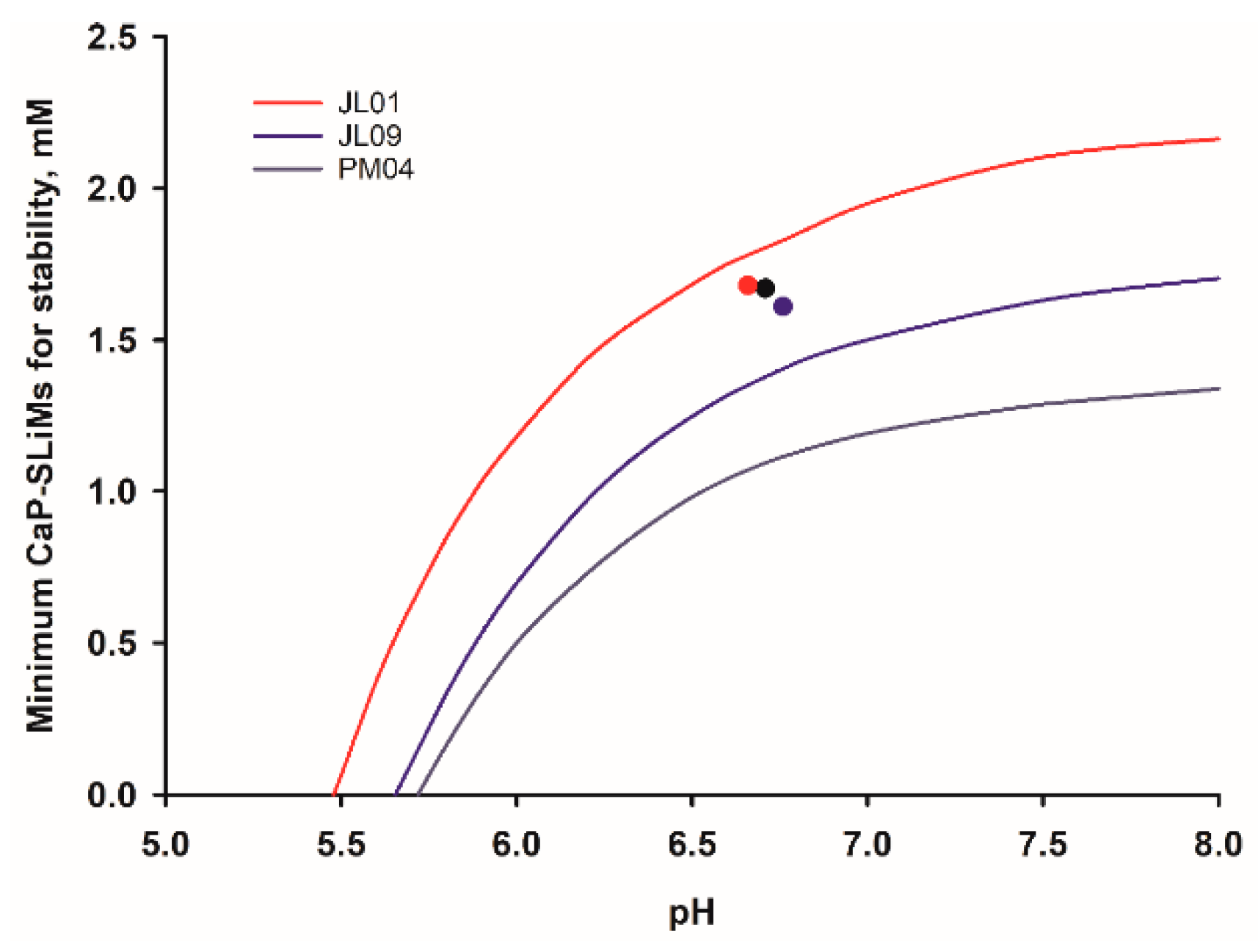
3.5. Thermodynamic Description of the Formation of CaP Nanoclusters Sequestered by Short Acidic Phosphopeptides
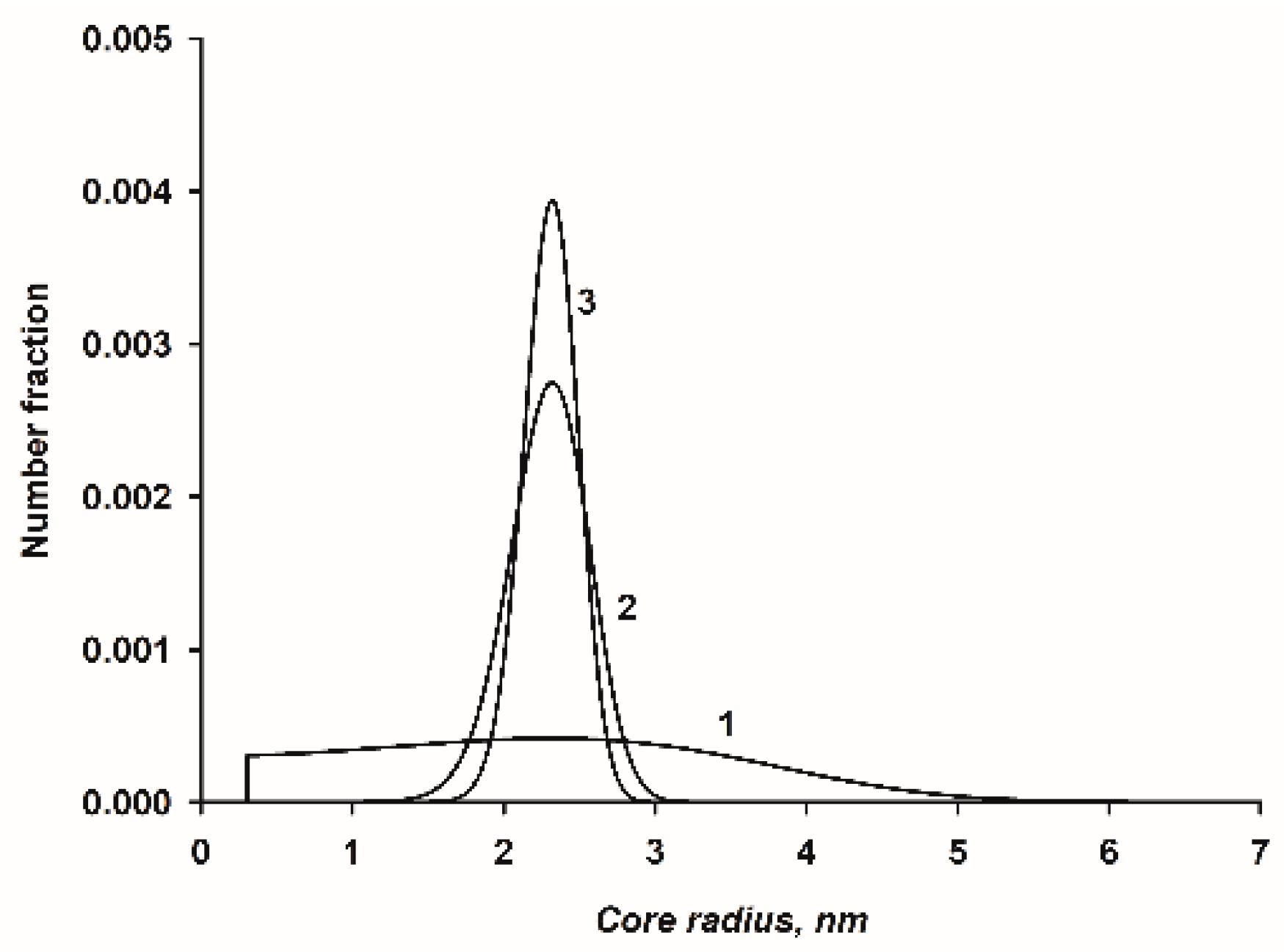
3.6. Evaluation of the Number Fraction Distribution
3.7. Stability of Biofluids and the Facile Co-Existence of Soft and Mineralized Tissues
4. Properties of Some Calcium Phosphate Nanocluster Complexes
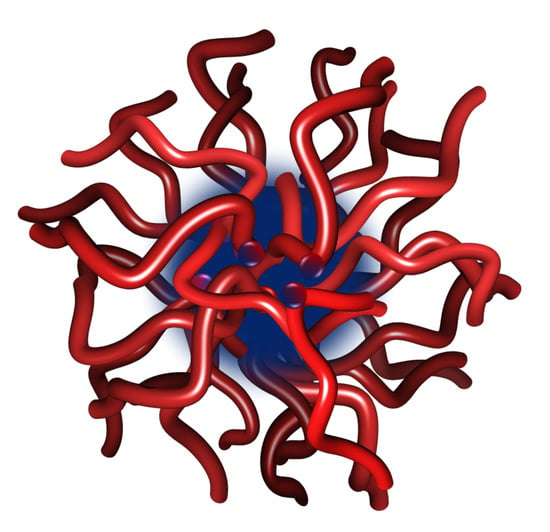
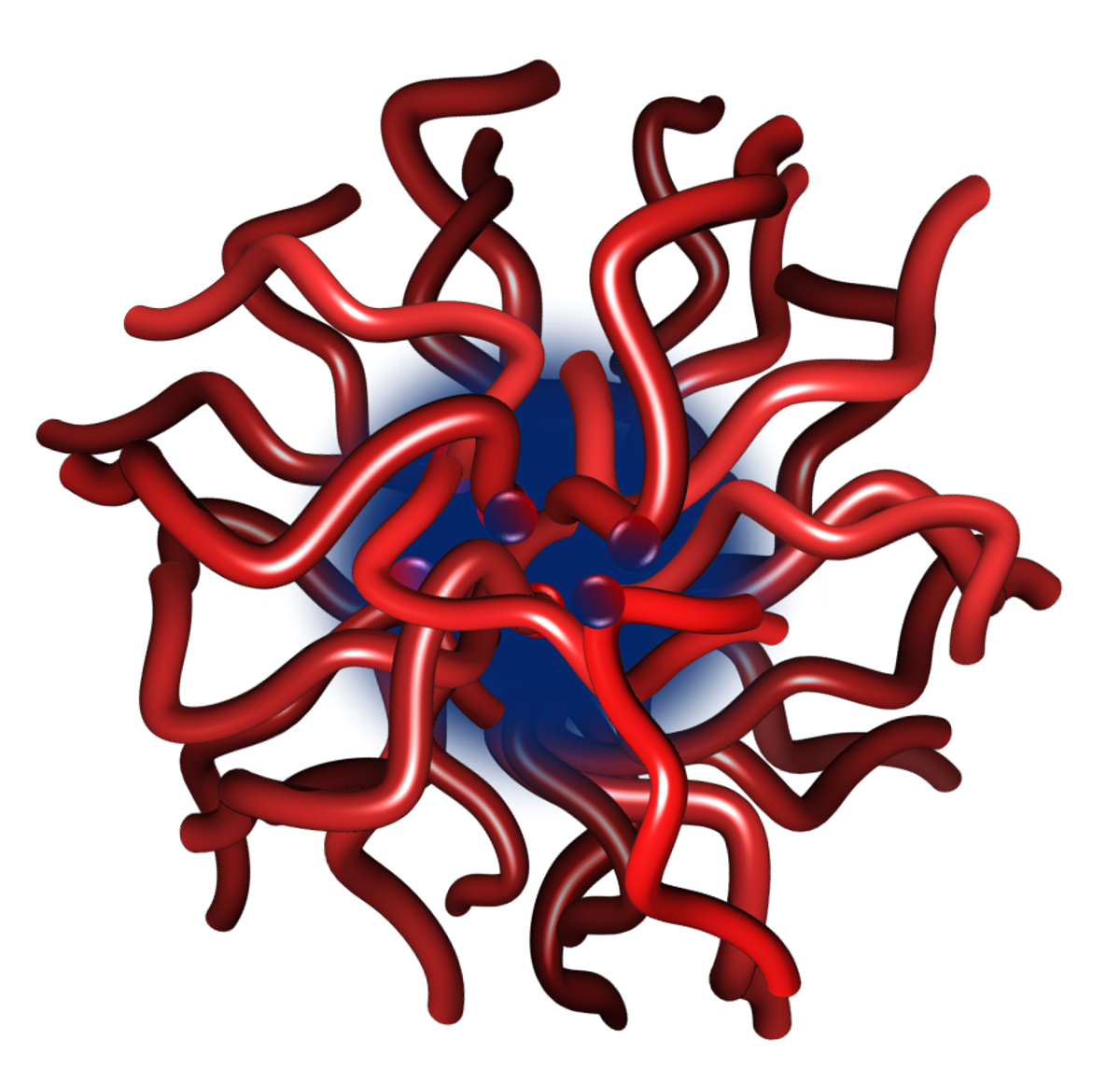
4.1. Size and Structure of Calcium Phosphate Nanoclusters Sequestered by β-Casein 1–25
4.2. Structure and Dynamics of a Calcium Phosphate Nanocluster Sequestered by OPN 1–149
4.3. Structures of Calcium Phosphate Nanoclusters Sequestered by Other Phosphopeptides or Mixtures of Phosphopeptides
5. Application to Biofluids
5.1. Stability of Milk
5.2. Stability of Blood Serum
5.3. Stability of Saliva
5.4. Tooth Remineralization
5.5. Stability of Urine
5.6. Mineralization in the Extracellular Matrix of Collagenous Hard Tissues
6. Summary and Conclusions
6.1. Equilibrium Complexes of CaP and Phosphoproteins
6.2. Protein Structural Requirements for CaP Sequestration
6.3. Core Radius of the Nanocluster Complexes
6.4. Biofluid Stability
6.5. Artificial and Biomimetic Materials
6.6. Basic Research Needs
Author Contributions
Funding
Conflicts of Interest
References
- George, A.; Veis, A. Phosphorylated Proteins and Control over Apatite Nucleation, Crystal Growth, and Inhibition. Chem. Rev. 2008, 108, 4670–4693. [Google Scholar] [CrossRef]
- Wang, L.; Nancollas, G.H. Pathways to Biomineralization and Biodemineralization of Calcium Phosphates: The Thermodynamic and Kinetic Controls. Dalton Trans. 2009, 15, 2665–2672. [Google Scholar] [CrossRef]
- Dorozhkin, S.V. Calcium Orthophosphates: Occurrence, Properties, Biomineralization, Pathological Calcification and Biomimetic Applications. Biomatter 2011, 1, 121–164. [Google Scholar] [CrossRef]
- Alvares, K. The Role of Acidic Phosphoproteins in Biomineralization. Connect. Tissue Res. 2014, 55, 34–40. [Google Scholar] [CrossRef]
- Dhami, N.K.; Sudhakara Reddy, M.; Mukherjee, A. Bionnineralization of Calcium Carbonates and Their Engineered Applications: A Review. Front. Microbiol. 2013, 4, 314. [Google Scholar] [CrossRef]
- Weiner, S.; Addadi, L. Crystallization Pathways in Biomineralization. Annu. Rev. Mater. Res. 2011, 41, 21–40. [Google Scholar] [CrossRef]
- Gelli, R.; Ridi, F.; Baglioni, P. The Importance of Being Amorphous: Calcium and Magnesium Phosphates in the Human Body. Adv. Colloid Interface Sci. 2019, 269, 219–235. [Google Scholar] [CrossRef]
- Jahnen-Dechent, W.; Heiss, A.; Schäfer, C.; Ketteler, M. Fetuin-A Regulation of Calcified Matrix Metabolism. Circ. Res. 2011, 108, 1494–1509. [Google Scholar] [CrossRef]
- Boskey, A.L.; Villarreal-Ramírez, E. Intrinsically Disordered Proteins and Biomineralization. Matrix Boil. 2016, 52, 43–59. [Google Scholar] [CrossRef]
- Tsuji, T.; Iijima, M.; Onuma, K. Biomineralization; Apatite Protein Interaction. In Bioinspiration: From Nano to Micro Scales, Biology, Medical Physics and Biomedical Engineering; Liu, X.Y., Ed.; Springer Science + Business Media: New York, NY, USA, 2012; pp. 135–159. [Google Scholar]
- Hunter, G.K.; O’Young, J.; Grohe, B.; Karttunen, M.; Goldberg, H.A. The Flexible Polyelectrolyte Hypothesis of Protein-Biomineral Interaction. Langmuir 2010, 26, 18639–18646. [Google Scholar] [CrossRef]
- Holt, C. Inorganic Constituents of Milk 3. The Colloidal Calcium Phosphate of Cows’ Milk. J. Dairy Res. 1982, 49, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Holt, C.; Lenton, S.; Nylander, T.; Sørensen, E.S.; Teixeira, S.C.M. Mineralisation of Soft and Hard Tissues and the Stability of Biofluids. J. Struct. Biol. 2014, 185, 383–396. [Google Scholar] [CrossRef]
- Kawasaki, K.; Suzuki, T.; Weiss, K.M. Genetic Basis for the Evolution of Vertebrate Mineralized Tissue. Proc. Natl. Acad. Sci. USA 2004, 101, 11356–11361. [Google Scholar] [CrossRef]
- Kawasaki, K.; Weiss, K.M. Mineralized Tissue and Vertebrate Evolution: The Secretory Calcium-Binding Phosphoprotein Gene Cluster. Proc. Natl. Acad. Sci. USA 2003, 100, 4060–4065. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, S.; Fuentealba, J.; Aze, A.; Hudson, C.; Yasuo, H.; Torrejon, M.; Escriva, H.; Marcellini, S. A Dynamic History of Gene Duplications and Losses Characterizes the Evolution of the SPARC Family in Eumetazoans. Proc. R. Soc. B Boil. Sci. 2013, 280, 20122963. [Google Scholar] [CrossRef]
- Mazzali, M.; Kipari, T.; Ophascharoensuk, V.; Wesson, J.; Johnson, R.; Hughes, J. Osteopontin—A Molecule for All Seasons. QJM Int. J. Med. 2002, 95, 3–13. [Google Scholar] [CrossRef]
- Wahlgren, N.M.; Holt, C.; Drakenberg, T. A Multi-Nuclear Magnetic Resonance Study of Calcium Phosphate Clusters Stabilised by ß-Casein 1–25. In Proceedings of the 2nd International Conference on Applications of Magnetic Resonance in Food Science, Aveiro, Portugal, 19–21 September 1994. [Google Scholar]
- Rao, A.; Drechsler, M.; Schiller, S.M.; Scheffner, M.; Gebauer, D.; Cölfen, H. Stabilization of Mineral Precursors by Intrinsically Disordered Proteins. Adv. Funct. Mater. 2018, 28. [Google Scholar] [CrossRef]
- Elsharkawy, S.; Al-Jawad, M.; Pantano, M.F.; Tejeda-Montes, E.; Mehta, K.; Jamal, H.; Agarwal, S.; Shuturminska, K.; Rice, A.; Tarakina, N.V.; et al. Protein disorder-Order Interplay to Guide the Growth of Hierarchical Mineralized Structures. Nat. Commun. 2018, 9, 2145. [Google Scholar] [CrossRef]
- Elsharkawy, S.; Mata, A. Hierarchical Biomineralization: From Nature’s Designs to Synthetic Materials for Regenerative Medicine and Dentistry. Adv. Heal. Mater. 2018, 7. [Google Scholar] [CrossRef]
- KalmarKalmár, L.; Homola, D.; Varga, G.; Tompa, P. Structural Disorder in Proteins Brings Order to Crystal Growth in Biomineralization. Bone 2012, 51, 528–534. [Google Scholar] [CrossRef]
- Ren, S.; Uversky, V.N.; Chen, Z.; Dunker, A.K.; Obradovic, Z. Short Linear Motifs Recognized by SH2, SH3 and Ser/Thr Kinase Domains Are Conserved in Disordered Protein Regions. BMC Genom. 2008, 9, S26. [Google Scholar] [CrossRef] [PubMed]
- Holt, C.; Sørensen, E.S.; Clegg, R.A. Role of calcium Phosphate Nanoclusters in the Control of Calcification. FEBS J. 2009, 276, 2308–2323. [Google Scholar] [CrossRef] [PubMed]
- Piovesan, D.; Tabaro, F.; Micetic, I.; Necci, M.; Quaglia, F.; Oldfield, C.J.; Aspromonte, M.C.; Davey, N.E.; Davidovic, R.; Dosztanyi, Z.; et al. Disprot 7.0: A Major Update of the Database of Disordered Proteins. Nucleic Acids Res. 2017, 45, D219–D227. [Google Scholar] [CrossRef]
- Kawasaki, K. The Scpp Gene Family and the Complexity of Hard Tissues in Vertebrates. Cells Tissues Organs 2011, 194, 108–112. [Google Scholar] [CrossRef]
- Fisher, L.W.; Torchia, D.; Fohr, B.; Young, M.; Fedarko, N.S. Flexible Structures of SIBLING Proteins, Bone Sialoprotein, and Osteopontin. Biochem. Biophys. Res. Commun. 2001, 280, 460–465. [Google Scholar] [CrossRef]
- Holt, C.; Raynes, J.K.; Carver, J.A. Sequence Characteristics Responsible for Protein-Protein Interactions in the Intrinsically Disordered Regions of Caseins, Amelogenins, and Small Heat-Shock Proteins. Biopolymers 2019, 110, e23319. [Google Scholar] [CrossRef]
- Arachchige, R.J.; Burton, S.D.; Lu, J.-X.; Ginovska, B.; Harding, L.K.; Taylor, M.E.; Tao, J.; Dohnalkova, A.; Tarasevich, B.J.; Buchko, G.W.; et al. Solid-State NMR Identification of Intermolecular Interactions in Amelogenin Bound to Hydroxyapatite. Biophys. J. 2018, 115, 1666–1672. [Google Scholar] [CrossRef]
- Ndao, M.; Dutta, K.; Bromley, K.M.; Lakshminarayanan, R.; Sun, Z.; Rewari, G.; Moradian-Oldak, J.; Evans, J.S. Probing the Self-Association, Intermolecular Contacts, and Folding Propensity of Amelogenin. Protein Sci. 2011, 20, 724–734. [Google Scholar] [CrossRef][Green Version]
- Wald, T.; Osickova, A.; Sulc, M.; Benada, O.; Semeradtova, A.; Rezabkova, L.; Veverka, V.; Bednarova, L.; Maly, J.; Macek, P.; et al. Intrinsically Disordered Enamel Matrix Protein Ameloblastin Forms Ribbon-like Supramolecular Structures via an N-terminal Segment Encoded by Exon 5*. J. Boil. Chem. 2013, 288, 22333–22345. [Google Scholar] [CrossRef]
- Carver, J.A.; Holt, C. Chapter Five-Functional and Dysfunctional Folding, Association and Aggregation of Caseins. In Advances in Protein Chemistry and Structural Biology; Donev, R., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 163–216. [Google Scholar]
- Carneiro, K.M.M.H.; Zhai, H.; Zhu, L.; Horst, J.A.; Sitlin, M.; Nguyen, M.; Wagner, M.; Simpliciano, C.; Milder, M.; Chen, C.-L.; et al. Amyloid-like Ribbons of Amelogenins in Enamel Mineralization. Sci. Rep. 2016, 6, 23105. [Google Scholar] [CrossRef]
- Wald, T.; Špoutil, F.; Osickova, A.; Prochazkova, M.; Benada, O.; Kasparek, P.; Bumba, L.; Klein, O.D.; Sedlacek, R.; Sebo, P.; et al. Intrinsically Disordered Proteins Drive Enamel Formation via an Evolutionarily Conserved Self-Assembly Motif. Proc. Natl. Acad. Sci. USA 2017, 114, E1641–E1650. [Google Scholar] [CrossRef] [PubMed]
- Dalgleish, D.G.; Parker, T.G. Binding of Calcium Ions to Bovine Alpha-S1-Casein and Precipitability of the Protein-Calcium Ion Complexes. J. Dairy Res. 1980, 47, 113–122. [Google Scholar] [CrossRef]
- Parker, T.G.; Dalgleish, D.G. Dalgleish. Binding of Calcium Ions to Bovine Beta-Casein. J. Dairy Res. 1981, 48, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Sleigh, R.W.; Mackinlay, A.G.; Pope, J. Nmr-Studies of the Phosphoserine Regions of Bovine Alpha-S1-Casein and Beta-Casein—Assignment of P-31 Resonances to Specific Phosphoserines and Cation Binding Studied by Measurement of Enhancement of H-1 Relaxation Rate. Biochim. Biophys. Acta 1983, 742, 175–183. [Google Scholar] [CrossRef]
- Sleigh, R.W.; Sculley, T.B.; Mackinlay, A.G. Mackinlay. Binding of Beta Casein to Hydroxyapatite-Effect of Phosphate Content and Location. J. Dairy Res. 1979, 46, 337–342. [Google Scholar] [CrossRef]
- Bijl, E.; Huppertz, T.; Van Valenberg, H.; Holt, C. A Quantitative Model of the Bovine Casein Micelle: Ion Equilibria and Calcium Phosphate Sequestration by Individual Caseins in Bovine Milk. Eur. Biophys. J. 2019, 48, 45–59. [Google Scholar] [CrossRef]
- Clegg, R.A.; Holt, C.; An, E. Coli over-Expression System for Multiply-Phosphorylated Proteins and Its Use in a Study of Calcium Phosphate Sequestration by Novel Recombinant Phosphopeptides. Protein Expr. Purif. 2009, 67, 23–34. [Google Scholar] [CrossRef]
- Rodriguez, D.E.; Thula-Mata, T.; Toro, E.J.; Yeh, Y.-W.; Holt, C.; Holliday, L.S.; Gower, L. Multifunctional Role of Osteopontin in Directing Intrafibrillar Mineralization of Collagen and Activation of Osteoclasts. Acta Biomater. 2014, 10, 494–507. [Google Scholar] [CrossRef]
- Wingender, B.; Bradley, P.; Saxena, N.; Ruberti, J.W.; Gower, L. Biomimetic Organization of Collagen Matrices to Template Bone-Like Microstructures. Matrix Biol. 2016, 52–54, 384–396. [Google Scholar] [CrossRef]
- Gower, L.B. Biomimetic Model Systems for Investigating the Amorphous Precursor Pathway and Its Role in Biomineralization. Chem. Rev. 2008, 108, 4551–4627. [Google Scholar] [CrossRef]
- Addison, W.N.; Nakano, Y.; Loisel, T.; Crine, P.; McKee, M.D. Mepe-Asarm Peptides Control Extracellular Matrix Mineralization by Binding to Hydroxyapatite: An Inhibition Regulated by Phex Cleavage of Asarm. J. Bone Miner. Res. 2008, 23, 1638–1649. [Google Scholar] [CrossRef]
- Christensen, B.; Schytte, G.N.; Scavenius, C.; Enghild, J.J.; McKee, M.D.; Sørensen, E.S. Fam20c-Mediated Phosphorylation of Mepe and Its Acidic Serine- and Aspartate-Rich Motif. JBMR Plus 2020, 4, e10378. [Google Scholar] [CrossRef]
- Salmon, B.; Bardet, C.; Khaddam, M.; Naji, J.; Coyac, B.R.; Baroukh, B.; Letourneur, F.; Lesieur, J.; Decup, F.; Le Denmat, D.; et al. Mepe-Derived Asarm Peptide Inhibits Odontogenic Differentiation of Dental Pulp Stem Cells and Impairs Mineralization in Tooth Models of X-Linked Hypophosphatemia. PLoS ONE 2013, 8, e56749. [Google Scholar] [CrossRef]
- Boskey, A.L.; Chiang, P.; Fermanis, A.; Brown, J.; Taleb, H.; David, V.; Rowe, P.S.N. Mepe’s Diverse Effects on Mineralization. Calcif. Tissue Int. 2010, 86, 42–46. [Google Scholar] [CrossRef]
- Minamizaki, T.; Yoshiko, Y. The Bioactive Acidic Serine- and Aspartate-Rich Motif Peptide. Curr. Protein Pept. Sci. 2015, 16, 196–202. [Google Scholar] [CrossRef]
- Schwartz, S.S.; Hay, D.I.; Schluckebier, S.K. Inhibition of Calcium Phosphate Precipitation by Human Salivary Statherin: Structure-Activity Relationships. Calcif. Tissue Int. 1992, 50, 511–517. [Google Scholar] [CrossRef]
- Gorski, J.P.; Kremer, E.A.; Chen, Y.; Ryan, S.; Fullenkamp, C.; DelViscio, J.; Jensen, K.; McKee, M.D. Bone Acidic Glycoprotein-75 Self-Associates to Form Macromolecular Complexes in Vitro and in Vivo with the Potential to Sequester Phosphate Ions. J. Cell. Biochem. 1997, 64, 547–564. [Google Scholar] [CrossRef]
- Walker, J.; Saraste, M.; Runswick, M.; Gay, N. Distantly Related Sequences in the Alpha-Subunits and Beta-Subunits of Atp Synthase, Myosin, Kinases and Other Atp-Requiring Enzymes and a Common Nucleotide Binding Fold. EMBO J. 1982, 1, 945–951. [Google Scholar] [CrossRef]
- Hirsch, A.K.H.; Fischer, F.R.; Diederich, F. Phosphate Recognition in Structural Biology. Angew. Chem. Int. Ed. 2007, 46, 338–352. [Google Scholar] [CrossRef]
- Bianchi, A.; Giorgi, C.; Ruzza, P.; Toniolo, C.; Milner-White, E.J. A Synthetic Hexapeptide Designed to Resemble a Proteinaceous P-Loop Nest Is Shown to Bind Inorganic Phosphate. Proteins Struct. Funct. Bioinform. 2012, 80, 1418–1424. [Google Scholar] [CrossRef]
- BBuckle, E.L.; Prakash, A.; Bonomi, M.; Sampath, J.; Pfaendtner, J.; Drobny, G.P. Solid-State Nmr and Md Study of the Structure of the Statherin Mutant Sna15 on Mineral Surfaces. J. Am. Chem. Soc. 2019, 141, 1998–2011. [Google Scholar] [CrossRef] [PubMed]
- Goobes, R.; Goobes, G.; Shaw, W.J.; Drobny, G.P.; Campbell, C.T.; Stayton, P.S. Thermodynamic Roles of Basic Amino Acids in Statherin Recognition of Hydroxyapatite. Biochemistry 2007, 46, 4725–4733. [Google Scholar] [CrossRef] [PubMed]
- Ndao, M.; Ash, J.T.; Stayton, P.S.; Drobny, G.P. The Role of Basic Amino Acids in the Molecular Recognition of Hydroxyapatite by Statherin Using Solid State Nmr. Surf. Sci. 2010, 604, L39–L42. [Google Scholar] [CrossRef] [PubMed]
- Holt, C.; Sawyer, L. Caseins as Rheomorphic Proteins-Interpretation of Primary and Secondary Structures of the Alpha-S1-Caseins, Beta-Caseins and Kappa-Caseins. J. Chem. Soc. Faraday Trans. 1993, 89, 2683–2692. [Google Scholar] [CrossRef]
- Holt, C.; Sawyer, L. Primary and Predicted Secondary Structures of the Caseins in Relation to Their Biological Functions. Protein Eng. Des. Sel. 1988, 2, 251–259. [Google Scholar] [CrossRef]
- Dyson, H.J. Making Sense of Intrinsicaly Disordered Proteins. Biophys. J. 2016, 110, 1013–1016. [Google Scholar] [CrossRef]
- Fujisawa, R.; Kuboki, Y. Conformation of Dentin Phosphophoryn Adsorbed on Hydroxyapatite Crystals. Eur. J. Oral Sci. 1998, 106, 249–253. [Google Scholar] [CrossRef]
- Samoshina, Y.; Nylander, T.; Shubin, V.; Bauer, R.; Eskilsson, K. Equilibrium Aspects of Polycation Adsorption on Silica Surface: How the Adsorbed Layer Responds to Changes in Bulk Solution. Langmuir 2005, 21, 5872–5881. [Google Scholar] [CrossRef]
- Lenton, S.; Seydel, T.; Nylander, T.; Holt, C.; Härtlein, M.; Teixeira, S.; Zaccai, G. Dynamic Footprint of Sequestration in the Molecular Fluctuations of Osteopontin. J. R. Soc. Interface 2015, 12, 20150506. [Google Scholar] [CrossRef]
- Moreno, E.C.; Varughese, K.; Hay, D.I. Effect of Human Salivary Proteins on the Precipitation Kinetics of Calcium Phosphate. Calcif. Tissue Int. 1979, 28, 7–16. [Google Scholar] [CrossRef]
- Hay, D.I.; Carlson, E.R.; Schluckebier, S.K.; Moreno, E.C.; Schlesinger, D.H. Inhibition of Calcium Phosphate Precipitation by Human Salivary Acidic Proline-Rich Proteins-Structure-Activity Relationships. Calcif. Tissue Int. 1987, 40, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Uversky, V.N.; Gillespie, J.R.; Fink, A.L. Why Are Natively Unfolded Proteins Unstructured under Physiologic Conditions? Proteins Struct. Funct. Gene. 2000, 41, 415–427. [Google Scholar] [CrossRef]
- Tagliabracci, V.S.; Wiley, S.E.; Guo, X.; Kinch, L.N.; Durrant, E.; Wen, J.; Xiao, J.; Cui, J.; Nguyen, K.B.; Engel, J.L.; et al. A Single Kinase Generates the Majority of the Secreted Phosphoproteome. Cell 2015, 161, 1619–1632. [Google Scholar] [CrossRef] [PubMed]
- Bah, A.; Forman-Kay, J.D. Modulation of Intrinsically Disordered Protein Function by Post-Translational Modifications. J. Biol. Chem. 2016, 291, 6696–6705. [Google Scholar] [CrossRef]
- Gao, J.J.; Xu, D. Correlation between Posttranslational Modification and Intrinsic Disorder in Protein. In Pacific Symposium on Biocomputing 2012; Altman, R.B., Dunker, A.K., Hunter, L., Murray, T., Klein, T.E., Eds.; World Scientific Publ. Co. Pte Ltd.: Singapore, 2012; pp. 94–103. [Google Scholar]
- He, G.; Gajjeraman, S.; Schultz, D.; Cookson, D.; Qin, C.; Butler, W.T.; Hao, J.; George, A. Spatially and Temporally Controlled Biomineralization is Facilitated by Interaction between Self-Assembled Dentin Matrix Protein 1 and Calcium Phosphate Nuclei in Solution. Biochemistry 2005, 44, 16140–16148. [Google Scholar] [CrossRef]
- Tartaix, P.H.; Doulaverakis, M.; George, A.; Fisher, L.W.; Butler, W.T.; Qin, C.L.; Salih, E.; Tan, M.L.; Fujimoto, Y.; Spevak, L.; et al. In Vitro Effects of Dentin Matrix Protein-1 on Hydroxyapatite Formation Provide Insights into in Vivo Functions. J. Biol. Chem. 2004, 279, 18115–18120. [Google Scholar] [CrossRef]
- Raj, P.A.; Johnsson, M.; Levine, M.J.; Nancollas, G.H. Salivary Statherin-Dependence on Sequence, Charge, Hydrogen-Bonding Potency and Helical Conformation for Adsorption to Hydroxyapatite and Inhibition of Mineralization. J. Biol. Chem. 1992, 267, 5968–5976. [Google Scholar]
- Zhang, J.; Wang, L.; Zhang, W.; Putnis, C.V. Phosphorylated/Nonphosphorylated Motifs in Amelotin Turn off/on the Acidic Amorphous Calcium Phosphate-to-Apatite Phase Transformation. Langmuir 2020, 36, 2102–2109. [Google Scholar] [CrossRef]
- He, G.; Ramachandran, A.; Dahl, T.; George, S.; Schultz, D.; Cookson, D.; Veis, A.; George, A. Phosphorylation of Phosphophoryn Is Crucial for Its Function as a Mediator of Biomineralization. J. Biol. Chem. 2005, 280, 33109–33114. [Google Scholar] [CrossRef]
- Baht, G.S.; O’Young, J.; Borovina, A.; Chen, H.; Tye, C.E.; Karttunen, M.; Lajoie, G.A.; Hunter, G.K.; Goldberg, H.A. Phosphorylation of Ser(136) Is Critical for Potent Bone Sialoprotein-Mediated Nucleation of Hydroxyapatite Crystals. Biochem. J. 2010, 428, 385–395. [Google Scholar] [CrossRef]
- Boskey, A.L.; Christensen, B.; Taleb, H.; Sørensen, E.S. Post-Translational Modification of Osteopontin: Effects on in Vitro Hydroxyapatite Formation and Growth. Biochem. Biophys. Res. Commun. 2012, 419, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Gericke, A.; Qin, C.; Spevak, L.; Fujimoto, Y.; Butler, W.T.; Sørensen, E.S.; Boskey, A.L. Importance of Phosphorylation for Osteopontin Regulation of Biomineralization. Calcif. Tissue Int. 2005, 77, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, B.A.; Portman, J.J.; Wolynes, P.G. Speeding Molecular Recognition by Using the Folding Funnel: The Fly-Casting Mechanism. Proc. Natl. Acad. Sci. USA 2000, 97, 8868–8873. [Google Scholar] [CrossRef] [PubMed]
- Nývlt, J. The Ostwald Rule of Stages. Cryst. Res. Technol. 1995, 30, 443–449. [Google Scholar] [CrossRef]
- Levin, A.; Mason, T.O.; Adler-Abramovich, L.; Buell, A.K.; Meisl, G.; Galvagnion, C.; Bram, Y.; Stratford, S.A.; Dobson, C.M.; Knowles, T.P.J.; et al. Ostwald’s Rule of Stages Governs Structural Transitions and Morphology of Dipeptide Supramolecular Polymers. Nat. Commun. 2014, 5, 5219. [Google Scholar] [CrossRef]
- Ostwald, W. Studien über die Bildung und Umwandlung Fester Körper. Z. Phys. Chem. 1897, 22, 289. [Google Scholar] [CrossRef]
- Threlfall, T. Structural and Thermodynamic Explanations of Ostwald’s Rule. Org. Process Res. Dev. 2003, 7, 1017–1027. [Google Scholar] [CrossRef]
- Combes, C.; Rey, C. Amorphous Calcium Phosphates: Synthesis, Properties and Uses in Biomaterials. Acta Biomater. 2010, 6, 3362–3378. [Google Scholar] [CrossRef]
- Christoffersen, M.R.; Christoffersen, J.; Kibalczyc, W. Apparent Solubilities of 2 Amorphous Calcium Phosphates and of Octacalcium Phosphate in the Temperature-Range 30–42-Degrees-C. J. Cryst. Growth 1990, 106, 349–354. [Google Scholar] [CrossRef]
- Meyer, J.L.; Eanes, E.D. Thermodynamic Analysis of Secondary Transition in Spontaneous Precipitation of Calcium-Phosphate. Calcif. Tissue Res. 1978, 25, 209–216. [Google Scholar] [CrossRef]
- Termine, J.D.; Eanes, E.D. Comparative Chemistry of Amorphous and Apatitic Calcium Phosphate Preparations. Calcif. Tissue Res. 1972, 10, 171–197. [Google Scholar] [CrossRef] [PubMed]
- Wuthier, R.E.; Rice, G.S.; Wallace, J.E.B.; Weaver, R.L.; LeGeros, R.Z.; Eanes, E.D. In Vitro Precipitation of Calcium-Phosphate under Intracellular Conditions—Formation of Brushite from an Amorphous Precursor in the Absence of Atp. Calcif. Tissue Int. 1985, 37, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Holt, C.; Van Kemenade, M.; Nelson, L.; Hukins, D.; Bailey, R.; Harries, J.; Hasnain, S.; De Bruyn, P. Amorphous Calcium Phosphates Prepared at PH 6.5 and 6.0. Mater. Res. Bull. 1989, 24, 55–62. [Google Scholar] [CrossRef]
- Posner, A.S.; Betts, F. Synthetic Amorphous Calcium-Phosphate and Its Relation to Bone-Mineral Structure. Accounts Chem. Res. 1975, 8, 273–281. [Google Scholar] [CrossRef]
- Lu, B.-Q.; Garcia, N.A.; Chevrier, D.M.; Zhang, P.; Raiteri, P.; Gale, J.D.; Gebauer, D. Short-Range Structure of Amorphous Calcium Hydrogen Phosphate. Cryst. Growth Des. 2019, 19, 3030–3038. [Google Scholar] [CrossRef]
- Betts, F.; Posner, A.S. X-Ray Radial Distribution Study of Amorphous Calcium Phosphate. Mater. Res. Bull. 1974, 9, 353–360. [Google Scholar] [CrossRef]
- Holt, C.; Hukins, D.W.L. Structural Analysis of the Environment of Calcium Ions in Crystalline and Amorphous Calcium Phosphates by X-ray Absorption Spectroscopy and a Hypothesis Concerning the Biological Function of the Casein Micelle. Int. Dairy J. 1991, 1, 151–165. [Google Scholar] [CrossRef]
- Williams, R.J.P. Biochemistry of Sodium, Potassium, Magnesium and Calcium. Q. Rev. 1970, 24, 331. [Google Scholar] [CrossRef]
- Lu, B.-Q.; Willhammar, T.; Sun, B.-B.; Hedin, N.; Gale, J.D.; Gebauer, D. Introducing the Crystalline Phase of Dicalcium Phosphate Monohydrate. Nat. Commun. 2020, 11, 1546. [Google Scholar] [CrossRef]
- Greco, K.; Bogner, R.H. Solution-Mediated Phase Transformation: Significance During Dissolution and Implications for Bioavailability. J. Pharm. Sci. 2012, 101, 2996–3018. [Google Scholar] [CrossRef] [PubMed]
- Gal, A.; Weiner, S.; Addadi, L. A Perspective on Underlying Crystal Growth Mechanisms in Biomineralization: Solution Mediated Growth versus Nanosphere Particle Accretion. Crystengcomm 2015, 17, 2606–2615. [Google Scholar] [CrossRef]
- Elliot, J.C. Structure and Chemistry of the Apatites and Other Calcium Orthophosphates; Studies in Inorganic Chemistry; Elsevier: Amsterdam, The Netherlands, 1994. [Google Scholar]
- Von Euw, S.; Wang, Y.; Laurent, G.; Drouet, C.; Babonneau, F.; Nassif, N.; Azais, T. Bone Mineral: New Insights into Its Chemical Composition. Sci. Rep. 2019, 9, 8456. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wang, L.; Qin, Y.; Sun, Z.; Henneman, Z.J.; Moradian-Oldak, J.; Nancollas, G.H. How Amelogenin Orchestrates the Organization of Hierarchical Elongated Microstructures of Apatite. J. Phys. Chem. B 2010, 114, 2293–2300. [Google Scholar] [CrossRef] [PubMed]
- Fincham, A.; Moradian-Oldak, J.; Simmer, J.P. The Structural Biology of the Developing Dental Enamel Matrix. J. Struct. Boil. 1999, 126, 270–299. [Google Scholar] [CrossRef]
- Enriquez, D.J.P.; Dell′arciprete, M.L.; Dittler, M.L.; Miñan, A.; Prieto, E.; Gonzalez, M.C. Amorphous Calcium Organophosphate Nanoshells as Potential Carriers for Drug Delivery to Ca2+-Enriched Surfaces. New J. Chem. 2020, 44, 7541–7551. [Google Scholar] [CrossRef]
- Liu, X.; Jing, X.; Liu, P.; Pan, M.; Liu, Z.; Dai, X.; Lin, J.; Li, Q.; Wang, F.; Yang, S.; et al. DNA Framework-Encoded Mineralization of Calcium Phosphate. Chem 2020, 6, 472–485. [Google Scholar] [CrossRef]
- Napper, D.H. Steric Stabilization. J. Colloid Interface Sci. 1977, 58, 390–407. [Google Scholar] [CrossRef]
- Van Der Lee, R.; Buljan, M.; Lang, B.; Weatheritt, R.J.; Daughdrill, G.W.; Dunker, A.K.; Fuxreiter, M.; Gough, J.; Gsponer, J.; Jones, D.T.; et al. Classification of Intrinsically Disordered Regions and Proteins. Chem. Rev. 2014, 114, 6589–6631. [Google Scholar] [CrossRef]
- Burke, E.; Guo, Y.; Colon, L.A.; Rahima, M.; Veis, A.; Nancollas, G.H. Influence of Polyaspartic Acid and Phosphophoryn on Octacalcium Phosphate Growth Kinetics. Colloids Surf. B Biointerfaces 2000, 17, 49–57. [Google Scholar] [CrossRef]
- De Bruyn, J.R.; Goiko, M.; Mozaffari, M.; Bator, D.; Dauphinee, R.L.; Liao, Y.; Flemming, R.L.; Bramble, M.S.; Hunter, G.K.; Goldberg, H.A. Dynamic Light Scattering Study of Inhibition of Nucleation and Growth of Hydroxyapatite Crystals by Osteopontin. PLoS ONE 2013, 8, e56764. [Google Scholar] [CrossRef]
- Goldberg, H.A.; Warner, K.J.; Li, M.C.; Hunter, G.K. Binding of Bone Sialoprotein, Osteopontin and Synthetic Polypeptides to Hydroxyapatite. Connect. Tissue Res. 2001, 42, 25–37. [Google Scholar] [CrossRef]
- Hunter, G.K.; Hauschka, P.V.; Poole, R.A.; Rosenberg, L.C.; Goldberg, H.A. Nucleation and Inhibition of Hydroxyapatite Formation by Mineralized Tissue Proteins. Biochem. J. 1996, 317, 59–64. [Google Scholar] [CrossRef]
- Pampena, D.A.; Robertson, K.A.; Litvinova, O.; Lajoie, G.; Goldberg, H.A.; Hunter, G.K. Inhibition of Hydroxyapatite Formation by Osteopontin Phosphopeptides. Biochem. J. 2004, 378, 1083–1087. [Google Scholar] [CrossRef]
- Tarasevich, B.J.; Howard, C.J.; Larson, J.L.; Snead, M.L.; Simmer, J.P.; Paine, M.; Shaw, W.J. The Nucleation and Growth of Calcium Phosphate by Amelogenin. J. Cryst. Growth 2007, 304, 407–415. [Google Scholar] [CrossRef]
- Holt, C.; Wahlgren, N.M.; Drakenberg, T. Ability of a Beta-Casein Phosphopeptide to Modulate the Precipitation of Calcium Phosphate by Forming Amorphous Dicalcium Phosphate Nanoclusters. Biochem. J. 1996, 314, 1035–1039. [Google Scholar] [CrossRef]
- Holt, C.; Timmins, P.A.; Errington, N.; Leaver, J. A Core-Shell Model of Calcium Phosphate Nanoclusters Stabilized by Beta-Casein Phosphopeptides, Derived from Sedimentation Equilibrium and Small-Angle X-Ray and Neutron-Scattering Measurements. Eur. J. Biochem. 1998, 252, 73–78. [Google Scholar] [CrossRef]
- Little, E.M.; Holt, C. An Equilibrium Thermodynamic Model of the Sequestration of Calcium Phosphate by Casein Phosphopeptides. Eur.Biophys. J. 2004, 33, 435–447. [Google Scholar] [CrossRef]
- Tercinier, L.; Ye, A.; Anema, S.G.; Singh, A.; Singh, H. Interactions of Casein Micelles with Calcium Phosphate Particles. J. Agric. Food Chem. 2014, 62, 5983–5992. [Google Scholar] [CrossRef]
- Fan, H.-X.; Gou, B.-D.; Gao, Y.-X.; Wu, G.; Zhang, T.-L. Pseudo-Equilibrium Equation of Calcium Phosphate Precipitation from Aqueous Solution. Phys. Chem. Chem. Phys. 2019, 21, 22057–22066. [Google Scholar] [CrossRef]
- Gebauer, D.; Cölfen, H. Prenucleation Clusters and Non-Classical Nucleation. Nano Today 2011, 6, 564–584. [Google Scholar] [CrossRef]
- Weber, E.M.M.; Kress, T.; Abergel, D.; Sewsurn, S.; Azais, T.; Kurzbach, D. Assessing the Onset of Calcium Phosphate Nucleation by Hyperpolarized Real-Time Nmr. Anal. Chem. 2020, 92, 7666–7673. [Google Scholar] [CrossRef] [PubMed]
- Garcia, N.A.; Malini, R.I.; Freeman, C.L.; Demichelis, R.; Raiteri, P.; Sommerdijk, N.A.J.M.; Harding, J.H.; Gale, J.D.; Freeeman, C.L. Simulation of Calcium Phosphate Prenucleation Clusters in Aqueous Solution: Association beyond Ion Pairing. Cryst. Growth Des. 2019, 19, 6422–6430. [Google Scholar] [CrossRef] [PubMed]
- Ashby, R.A.; Byrne, J.P.; Györy, Á.Z. Urine Is a Saturated Equilibrium and Not a Metastable Supersaturated Solution: Evidence from Crystalluria and the General Composition of Calcium Salt and Uric Acid Calculi. Urol. Res. 1999, 27, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Györy, A.Z.; Ashby, R. Equilibrium Versus Supersaturated Urine Hypothesis in Calcium Salt Urolithiasis: A New Theoretical and Practical Approach to a Clinical Problem. Scanning Microsc. 1999, 13, 261–265. [Google Scholar]
- Kavanagh, J.P. A Critical Appraisal of the Hypothesis That Urine Is a Saturated Equilibrium with Respect to Stone-Forming Calcium Salts. BJU Int. 2001, 87, 589–598. [Google Scholar] [CrossRef]
- Eidelman, N.; Chow, L.C.; Brown, W.E. Calcium Phosphate Saturation Levels in Ultrafiltered Serum. Calcif. Tissue Int. 1987, 40, 71–78. [Google Scholar] [CrossRef]
- Grøn, P. Saturation of Human Saliva with Calcium Phosphates. Arch. Oral Biol. 1973, 18, 1385–1392. [Google Scholar] [CrossRef]
- May, P.M.; Rowland, D. Jess, a Joint Expert Speciation System-Vi: Thermodynamically-Consistent Standard Gibbs Energies of Reaction for Aqueous Solutions. New J. Chem. 2018, 42, 7617–7629. [Google Scholar] [CrossRef]
- Larsen, M.J.; Pearce, E.I.F. Pearce. Saturation of Human Saliva with Respect to Calcium Salts. Arch. Oral Biol. 2003, 48, 317–322. [Google Scholar] [CrossRef]
- Walser, M. Separate Effects of Hyperparathyroidism, Hypercalcaemia of Malignancy, Renal Failure, and Acidosis on State of Calcium, Phosphate and Other Ions in Plasma. J. Clin. Investig. 1962, 41, 1454–1471. [Google Scholar] [CrossRef]
- Walser, M. Ion Association 6. Interactions between Calcium, Magnesium, Inorganic Phosphate, Citrate and Protein in Normal Human Plasma. J. Clin. Investig. 1961, 40, 723–730. [Google Scholar] [CrossRef] [PubMed]
- White, J.C.D.; Davies, D.T. The Relation between the Chemical Composition of Milk and the Stability of the Caseinate Complex. 1. General Introduction, Description of Samples, Methods and Chemical Composition of Samples. J. Dairy Res. 1958, 25, 236–255. [Google Scholar] [CrossRef]
- Holt, C. An Equilibrium Thermodynamic Model of the Sequestration of Calcium Phosphate by Casein Micelles and Its Application to the Calculation of the Partition of Salts in Milk. Eur. Biophys. J. Biophys. Lett. 2004, 33, 421–434. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, J.K.; Jakobsen, H.J.; Nielsen, N.; Petersen, T.E.; Rasmussen, L.K. Solid-State Magic-Angle Spinning 31p-Nmr Studies of Native Casein Micelles. Eur. J. Biochem. 1995, 230, 454–459. [Google Scholar] [CrossRef] [PubMed]
- de Kruif, C.G. The Structure of Casein Micelles: A Review of Small-Angle Scattering Data. J. Appl. Crystallogr. 2014, 47, 1479–1489. [Google Scholar] [CrossRef]
- Holt, C. The Milk Salts and Their Interaction with Casein. In Advanced Dairy Chemistry; Fox, P.F., Ed.; Chapman and Hall: London, UK, 1997; pp. 233–254. [Google Scholar]
- Ingham, B.; Erlangga, G.D.; Smialowska, A.; Kirby, N.M.; Wang, C.; Matia-Merino, L.; Haverkamp, R.G.; Carr, A.J. Solving the Mystery of the Internal Structure of Casein Micelles. Soft Matter 2015, 11, 2723–2725. [Google Scholar] [CrossRef]
- Kamigaki, T.; Ito, Y.; Nishino, Y.; Miyazawa, A. Microstructural Observation of Casein Micelles in Milk by Cryo-Electron Microscopy of Vitreous Sections (Cemovis). Microscopy 2018, 67, 164–170. [Google Scholar] [CrossRef]
- Hettiarachchi, C.A.; Swulius, M.T.; Harte, F. Assessing Constituent Volumes and Morphology of Bovine Casein Micelles Using Cryo-Electron Tomography. J. Dairy Sci. 2020, 103, 3971–3979. [Google Scholar] [CrossRef]
- McMahon, D.; Oommen, B.S. Casein Micelle Structure, Functions and Interactions. In Advanced Dairy Chemistry; Fox, P.F., McSweeney, P.L.H., Eds.; Springer: New York, NY, USA, 2012; pp. 185–210. [Google Scholar]
- Volmer, M.; Weber, A. Germ-Formation in Oversaturated Figures. Z. Phys. Chem. Stochiom. Verwandtschaftslehre 1926, 119, 277–301. [Google Scholar]
- Prigogine, I.; Defay, R.; Everett, D.H. Chemical Thermodynamics; Everett, D.H., Ed.; John Wiley & Sons: New York, NY, USA, 1962. [Google Scholar]
- Holt, C. A Quantitative Calcium Phosphate Nanocluster Model of the Casein Micelle: The Average Size, Size Distribution and Surface Charge Density. 2020; Unpublished work. [Google Scholar]
- Holt, C. Unfolded Phosphopolypeptides Enable Soft and Hard Tissues to Coexist in the Same Organism with Relative Ease. Curr. Opin. Struct. Boil. 2013, 23, 420–425. [Google Scholar] [CrossRef]
- Neville, M.C.; Keller, R.P.; Casey, C.; Allen, J.C. Calcium Partitioning in Human and Bovine Milk. J. Dairy Sci. 1994, 77, 1964–1975. [Google Scholar] [CrossRef]
- Jahnen-Dechent, W.; Schäfer, C.; Ketteler, M.; McKee, M.D. Mineral Chaperones: A Role for Fetuin-a and Osteopontin in the Inhibition and Regression of Pathologic Calcification. J. Mol. Med. 2008, 86, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Hill, A.V. Proceedings of the Physiological Society: 22 January 1910. J. Physiol. 1910, 40, i–vii. [Google Scholar]
- Pyne, G.T.; McGann, T.C.A. The Colloidal Phosphate of Milk: II. Influence of Citrate. J. Dairy Res. 1960, 27, 9–17. [Google Scholar] [CrossRef]
- McGann, T.C.A.; Pyne, G.T. The Colloidal Phosphate of Milk: III. Nature of its Association with Casein. J. Dairy Res. 1960, 27, 403–417. [Google Scholar] [CrossRef]
- Holt, C.; van Kemenade, M.J.J.M.; Nelson, L.S.; Sawyer, L.; Harries, J.E.; Bailey, R.T.; Hukins, D.W.L. Composition and Structure of Micellar Calcium-Phosphate. J. Dairy Res. 1989, 56, 411–416. [Google Scholar] [CrossRef]
- Lyster, R.L.J.; Mann, S.; Parker, S.B.; Williams, R.J.P. Nature of Micellar Calcium Phosphate in Cows’ Milk as Studied by High Resolution Electron Microscopy. Biochim. Biophys. Acta 1984, 801, 315–317. [Google Scholar] [CrossRef]
- McGann, T.C.A.; Kearney, R.D.; Buchheim, W.; Posner, A.S.; Betts, F.; Blumenthal, N.C. Amorphous Calcium-Phosphate in Casein Micelles of Bovine-Milk. Calcif. Tissue Int. 1983, 35, 821–823. [Google Scholar] [CrossRef]
- Ono, T.; Ohotawa, T.; Takagi, Y. Complexes of Casein Phosphopetides and Calcium Phosphate Prepared from Casein Micelles by Tryptic Digestion. Biosci. Biotechnol. Biochem. 1994, 58, 1376–1380. [Google Scholar] [CrossRef]
- Holt, C.; de Kruif, C.G.; Tuinier, R.; Timmins, P.A. Substructure of Bovine Casein Micelles by Small-Angle X-Ray and Neutron Scattering. Colloids Surf. Physicochem. Eng. Asp. 2003, 213, 275–284. [Google Scholar] [CrossRef]
- Ellegård, K.H.; Gammelgård-Larsen, C.; Sørensen, E.S.; Fedosov, S. Process Scale Chromatographic Isolation, Characterization and Identification of Tryptic Bioactive Casein Phosphopeptides. Int. Dairy J. 1999, 9, 639–652. [Google Scholar] [CrossRef]
- Cochrane, N.J.; Cai, F.; Huq, N.L.; Burrow, M.F.; Reynolds, E.C. New Approaches to Enhanced Remineralization of Tooth Enamel. J. Dent. Res. 2010, 89, 1187–1197. [Google Scholar] [CrossRef] [PubMed]
- Cross, K.J.; Huq, N.L.; Reynolds, E.C. Casein Phosphopeptide-Amorphous Calcium Phosphate Nanocomplexes: A Structural Model. Biochemistry 2016, 55, 4316–4325. [Google Scholar] [CrossRef] [PubMed]
- On-Nom, N.; Grandison, A.S.; Lewis, M.J. Measurement of Ionic Calcium, PH, and Soluble Divalent Cations in Milk at High Temperature. J. Dairy Sci. 2010, 93, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Gaucheron, F. (Ed.) Minéraux et Produits Laitiers; Tec & Doc Paris: Paris, France, 2004. [Google Scholar]
- Pouliot, Y.; Boulet, M.; Paquin, P. Observations on the Heat-Induced Salt Balance Changes in Milk 1. Effect of Heating Time between 4 Degrees C and 90 Degrees C. J. Dairy Res. 1989, 56, 185–192. [Google Scholar] [CrossRef]
- Depalle, B.; McGilvery, C.M.; Nobakhti, S.; Aldegaither, N.; Shefelbine, S.J.; Porter, A.E. Osteopontin Regulates Type I Collagen Fibril Formation in Bone Tissue. Acta Biomater. 2020, in press. [Google Scholar] [CrossRef]
- Foster, B.L.; Ao, M.; Salmon, C.R.; Chavez, M.B.; Kolli, T.N.; Tran, A.B.; Chu, E.Y.; Kantovitz, K.R.; Yadav, M.; Narisawa, S.; et al. Osteopontin Regulates Dentin and Alveolar Bone Development and Mineralization. Bone 2018, 107, 196–207. [Google Scholar] [CrossRef]
- Jinyan, S.; Wang, C.; Zhang, D.; Wang, B.; Hou, W.; Zhou, Y. Osteopontin in Bone Metabolism and Bone Diseases. Med. Sci. Monit. 2020, 26, e919159-1. [Google Scholar]
- Franca, C.M.; Thrivikraman, G.; Athirasala, A.; Tahayeri, A.; Gower, L.B.; Bertassoni, L.E. The Influence of Osteopontin-Guided Collagen Intrafibrillar Mineralization on Pericyte Differentiation and Vascularization of Engineered Bone Scaffolds. J. Biomed. Mater. Res. Part B Appl. Biomater. 2019, 107, 1522–1532. [Google Scholar] [CrossRef]
- Thurgood, L.A.; Ryall, R.L. Proteomic Analysis of Proteins Selectively Associated with Hydroxyapatite, Brushite, and Uric Acid Crystals Precipitated from Human Urine. J. Proteome Res. 2010, 9, 5402–5412. [Google Scholar] [CrossRef]
- Hoyer, J.R.; Asplin, J.R.; Otvos, L. Phosphorylated Osteopontin Peptides Suppress Crystallization by Inhibiting the Growth of Calcium Oxalate Crystals. Kidney Int. 2001, 60, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Li, S.Y.; Wang, L.J. Phosphorylated Osteopontin Peptides Inhibit Crystallization by Resisting the Aggregation of Calcium Phosphate Nanoparticles. Crystengcomm 2012, 14, 8037–8043. [Google Scholar] [CrossRef]
- Lenton, S.; Grimaldo, M.; Roosen-Runge, F.; Schreiber, F.; Nylander, T.; Clegg, R.A.; Holt, C.; Hartlein, M.; Sakai, V.G.; Seydel, T.; et al. Effect of Phosphorylation on a Human-Like Osteopontin Peptide. Biophys. J. 2017, 112, 1586–1596. [Google Scholar] [CrossRef] [PubMed]
- Platzer, G.; Żerko, S.; Saxena, S.; Koźmiński, W.; Konrat, R. 1h, 15n, 13c Resonance Assignment of Human Osteopontin. Biomol. NMR Assign. 2015, 9, 289–292. [Google Scholar] [CrossRef][Green Version]
- Schack, L.; Lange, A.; Kelsen, J.; Agnholt, J.; Christensen, B.; Petersen, T.E.; Sorensen, E.S. Considerable Variation in the Concentration of Osteopontin in Human Milk, Bovine Milk, and Infant Formulas. J. Dairy Sci. 2009, 92, 5378–5385. [Google Scholar] [CrossRef]
- Christensen, B.; Sorensen, E.S. Osteopontin Is Highly Susceptible to Cleavage in Bovine Milk and the Proteolytic Fragments Bind the Alpha(V)Beta(3)-Integrin Receptor. J. Dairy Sci. 2014, 97, 136–146. [Google Scholar] [CrossRef]
- Sørensen, E.S.; Hojrup, P.; Petersen, T.E. Posttranslational Modifications of Bovine Osteopontin-Identification of 28 Phosphorylation and 3 O-Glycosylation Sites. Protein Sci. 1995, 4, 2040–2049. [Google Scholar] [CrossRef]
- Ibsen, C.J.S.; Gebauer, D.; Birkedal, H. Osteopontin Stabilizes Metastable States Prior to Nucleation during Apatite Formation. Chem. Mater. 2016, 28, 8550–8555. [Google Scholar] [CrossRef]
- Lenton, S.; Nylander, T.; Teixeira, S.C.M.; Holt, C. A Review of the Biology of Calcium Phosphate Sequestration with Special Reference to Milk. Dairy Sci. Technol. 2015, 95, 3–14. [Google Scholar] [CrossRef]
- Lenton, S.; Nylander, T.; Holt, C.; Sawyer, L.; Härtlein, M.; Müller, H.; Teixeira, S.C.M. Structural Studies of Hydrated Samples of Amorphous Calcium Phosphate and Phosphoprotein Nanoclusters. Eur. Biophys. J. Biophys. Lett. 2016, 45, 405–412. [Google Scholar] [CrossRef]
- Cross, K.J.; Huq, N.L.; Stanton, D.P.; Sum, M.; Reynolds, E.C. Nmr Studies of a Novel Calcium, Phosphate and Fluoride Delivery Vehicle-Alpha(S1)-Casein(59–79) by Stabilized Amorphous Calcium Fluoride Phosphate Nanocomplexes. Biomaterials 2004, 25, 5061–5069. [Google Scholar] [CrossRef]
- Cross, K.J.; Huq, N.L.; Palamara, J.E.; Perich, J.W.; Reynolds, E.C. Physicochemical Characterization of Casein Phosphopeptide-Amorphous Calcium Phosphate Nanocomplexes. J. Biol. Chem. 2005, 280, 15362–15369. [Google Scholar] [CrossRef] [PubMed]
- Cesaro, L.; Pinna, L.A. Prevalence and Significance of the Commonest Phosphorylated Motifs in the Human Proteome: A Global Analysis. Cell. Mol. Life Sci. 2020, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Hay, D.I.; Smith, D.J.; Schluckebier, S.K.; Moreno, E.C. Basic Biological Sciences Relationship between Concentration of Human Salivary Statherin and Inhibition of Calcium Phosphate Precipitation in Stimulated Human Parotid Saliva. J. Dent. Res. 1984, 63, 857–863. [Google Scholar] [CrossRef] [PubMed]
- Hay, D.I.; Moreno, E.C.; Schlesinger, D.H. Phosphoprotein Inhibitors of Calcium Phosphate Precipitation from Salivary Secretions. Inorg. Perspect. Biol. Med. 1979, 2, 271–285. [Google Scholar]
- Davenport, H.W. The ABC of Acid-Base Chemistry. The Elements of Physiological Blood-Gas Chemistry for Medical Students and Physicians, 6th ed.; University of Chicago Press: Chicago, IL, USA, 1974. [Google Scholar]
- Miura, Y.; Iwazu, Y.; Shiizaki, K.; Akimoto, T.; Kotani, K.; Kurabayashi, M.; Kurosu, H.; Kuro-o, M. Identification and Quantification of Plasma Calciprotein Particles with Distinct Physical Properties in Patients with Chronic Kidney Disease. Sci. Rep. 2018, 8, 1256. [Google Scholar] [CrossRef] [PubMed]
- Price, P.A.; Nguyen, T.M.T.; Williamson, M.K. Biochemical Characterization of the Serum Fetuin-Mineral Complex. J. Biol. Chem. 2003, 278, 22153–22160. [Google Scholar] [CrossRef]
- Brylka, L.; Jahnen-Dechent, W. The Role of Fetuin-A in Physiological and Pathological Mineralization. Calcif. Tissue Int. 2013, 93, 355–364. [Google Scholar] [CrossRef]
- Jahnen-Dechent, W.; Smith, E.R. Nature’s Remedy to Phosphate Woes: Calciprotein Particles Regulate Systemic Mineral Metabolism. Kidney Int. 2020, 97, 648–651. [Google Scholar] [CrossRef]
- Heiss, A.; Eckert, T.; Aretz, A.; Richtering, W.; van Dorp, W.; Schafer, C.; Jahnen-Dechent, W. Hierarchical Role of Fetuin-A and Acidic Serum Proteins in the Formation and Stabilization of Calcium Phosphate Particles. J. Biol. Chem. 2008, 283, 14815–14825. [Google Scholar] [CrossRef]
- Jahnen-Dechent, W.; Büscher, A.; Köppert, S.; Heiss, A.; Kuro-o, M.; Smith, E.R. Mud in the Blood the Role of Protein-Mineral Complexes and Extracellular Vesicles in Biomineralisation and Calcification. J. Struct. Biol. 2020, 212, 107577. [Google Scholar] [CrossRef]
- Koppert, S.; Buscher, A.; Babler, A.; Ghallab, A.; Buhl, E.M.; Latz, E.; Hengstler, J.G.; Smith, E.R.; Jahnen-Dechent, W. Cellular Clearance and Biological Activity of Calciprotein Particles Depend on Their Maturation State and Crystallinity. Front. Immunol. 2018, 9, 17. [Google Scholar] [CrossRef] [PubMed]
- Holt, C. Interrelationships of the Concentrations of Some Ionic Constituents of Human-Milk and Comparison with Cow and Goat Milk. Comp. Biochem. Physiol. 1993, 104, 35–41. [Google Scholar] [CrossRef]
- Halperin, M.L.; Dhadli, S.C.; Kamel, K.S. Physiology of Acid-Base Balance: Links with Kidney Stone Prevention. Semin. Nephrol. 2006, 26, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Hill Michael, G.; Königsberger, E.; May Peter, M. Mineral Precipitation and Dissolution in the Kidney. Am. Mineral. 2017, 102, 701–710. [Google Scholar] [CrossRef]
- Khan, S.R.; Kok, D.J. Modulators of Urinary Stone Formation. Front. Biosci. Landmark 2004, 9, 1450–1482. [Google Scholar] [CrossRef]
- Kuruppath, S.; Bisana, S.; Sharp, J.A.; Lefevre, C.; Kumar, S.; Nicholas, K.R. Monotremes and Marsupials: Comparative Models to Better Understand the Function of Milk. J. Biosci. 2012, 37, 581–588. [Google Scholar] [CrossRef]
- McClellan, H.L.; Miller, S.J.; Hartmann, P.E. Evolution of Lactation: Nutrition V. Protection with Special Reference to Five Mammalian Species. Nutr. Res. Rev. 2008, 21, 97–116. [Google Scholar] [CrossRef]
- Oftedal, O.T. The Evolution of Milk Secretion and Its Ancient Origins. Animal 2012, 6, 355–368. [Google Scholar] [CrossRef]
- Holt, C.; Carver, J.A. Darwinian Transformation of a ‘Scarcely Nutritious Fluid’ into Milk. J. Evol. Biol. 2012, 25, 1253–1263. [Google Scholar] [CrossRef]
- Jenness, R.; Sloan, R.E. The Composition of Milk of Various Species: A Review. Dairy Sci. Abstr. 1970, 32, 599–612. [Google Scholar]
- Eisert, R.; Oftedal, O.T. Capital expenditure and income (foraging) during pinniped lactation: The example of the weddell seal (Leptonychotes weddellii). In Smithsonian at the Poles: Contributions to International Polar Year Science; Lang, M.A., Miller, S.E., Krupnik, I., Eds.; Smithsonian Institute: Washington, DC, USA, 2009; pp. 335–346. [Google Scholar]
- Jenness, R.; Holt, C. Casein and Lactose Concentrations in Milk of 31 Species Are Negatively Correlated. Experientia 1987, 43, 1015–1018. [Google Scholar] [CrossRef] [PubMed]
- Aoki, T.; Sakamoto, H.; Kako, Y. Cross-Linking of Caseins by Colloidal Calcium Phosphate in the Presence of Urea. Int. Dairy J. 1991, 1, 67–75. [Google Scholar] [CrossRef]
- Umeda, T.; Aoki, T. Relation between Micelle Size and Micellar Calcium Phosphate. Milchwiss. Milk Sci. Int. 2002, 57, 131–133. [Google Scholar]
- Wang, Q.; Holt, C.; Nylander, T.; Ma, Y. Salt Partition, Ion Equilibria and the Structure, Composition and Solubility of Micellar Calcium Phosphate in Bovine Milk with Added Calcium Salts. J. Dairy Sci. 2020, in press. [Google Scholar]
- Follows, D.; Holt, C.; Nylander, T.; Thomas, R.K.; Tiberg, F. Beta-Casein Adsorption at the Silicon Oxide-Aqueous Solution Interface: Calcium Ion Effects. Biomacromolecules 2004, 5, 319–325. [Google Scholar] [CrossRef]
- Follows, D.; Holt, C.; Thomas, R.K.; Tiberg, F.; Fragneto, G.; Nylander, T. Co-Adsorption of Beta-Casein and Calcium Phosphate Nanoclusters (Cpn) at Hydrophilic and Hydrophobic Solid-Solution Interfaces Studied by Neutron Reflectometry. Food Hydrocoll. 2011, 25, 724–733. [Google Scholar] [CrossRef]
- Blackburn, D.G.; Hayssen, V.; Murphy, C.J. The Origin of Lactation and the Evolution of Milk: A Review with New Hypotheses. Mammal. Rev. 1989, 19, 1–26. [Google Scholar] [CrossRef]
- Capuco, A.V.; Michael Akers, R. The Origin and Evolution of Lactation. J. Biol. 2009, 8, 37. [Google Scholar] [CrossRef]
- Haldane, J.B.S. The Possible Evolution of Lactation. Zool. Jb. Syst. 1965, 92, 41–48. [Google Scholar]
- Kawasaki, K.; Lafont, A.-G.; Sire, J.-Y. The Evolution of Casein Genes from Tooth Genes before the Origin of Mammals. Mol. Biol. Evol. 2011, 28, 2053–2061. [Google Scholar] [CrossRef] [PubMed]
- Lefèvre, C.M.; Sharp, J.A.; Nicholas, K.R. Evolution of Lactation: Ancient Origin and Extreme Adaptations of the Lactation System. Ann. Rev. Genom. Hum. Gene 2010, 11, 219–238. [Google Scholar] [CrossRef] [PubMed]
- Vorbach, C.; Capecchi, M.R.; Penninger, J.M. Evolution of the Mammary Gland from the Innate Immune System? Bioessays 2006, 28, 606–616. [Google Scholar] [CrossRef] [PubMed]
- Beems, R.B.; Gruys, E.; Spit, B.J. Amyloid in Corpora Amylacea of Rat Mammary Gland. Vet. Pathol. 1978, 15, 347–352. [Google Scholar] [CrossRef]
- Brooker, B.E. Origin Structure and Occurrence of Corpora Amylacea in Bovine Mammary Gland and in Milk. Cell Tissue Res. 1978, 191, 525–538. [Google Scholar] [CrossRef]
- Niewold, T.A.; Murphy, C.L.; Hulskamp-Koch, C.A.M.; Tooten, P.C.J.; Gruys, E. Casein Related Amyloid, Characterization of a New and Unique Amyloid Protein Isolated from Bovine Corpora Amylacea. Amyloid Int. J. Exp. Clin. Investig. 1999, 6, 244–249. [Google Scholar] [CrossRef]
- Reid, I.M. Corpora Amylacea of Bovine Mammary Gland-Histochemical and Electron Microscopic Evidence for Their Amyloid Nature. J. Comp. Pathol. 1972, 82, 409–413. [Google Scholar] [CrossRef]
- Hagsten, C.; Altskar, A.; Gustafsson, S.; Loren, N.; Hamberg, L.; Innings, F.; Paulsson, M.; Nylander, T. Composition and Structure of High Temperature Dairy Fouling. Food Struct. 2016, 7, 13–20. [Google Scholar] [CrossRef]
- Qi, P.X.; Wickham, E.D.; Farrell, H.M. Thermal and Alkaline Denaturation of Bovine Beta-Casein. Protein J. 2004, 23, 389–402. [Google Scholar] [CrossRef]
- Carver, J.A.; Holt, C. Functional and Dysfunctional Folding, Association and Aggregation of Caseins. Adv. Protein Chem. Struct. Biol. 2020, 118, 163–216. [Google Scholar]
- Carver, J.A.; Ecroyd, H.; Truscott, R.J.W.; Thorn, D.C.; Holt, C. Proteostasis and the Regulation of Intra- and Extracellular Protein Aggregation by Atp-Independent Molecular Chaperones: Lens A-Crystallins and Milk Caseins. Acc. Chem. Res. 2018, 51, 745–752. [Google Scholar] [CrossRef] [PubMed]
- McMeekin, T.L. Milk Proteins in Retrospect. In Milk Proteins Chemistry and Molecular Biology; McKenzie, H.A., Ed.; Academic Press: New York, NY, USA; London, UK, 1970; pp. 3–15. [Google Scholar]
- Foley, R.N.; Parfrey, P.S.; Harnett, J.D.; Kent, G.M.; Martin, C.J.; Murray, D.C.; Barre, P.E. Clinical and Echocardiographic Disease in Patients Starting End-Stage Renal Disease Therapy. Kidney Int. 1995, 47, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Pasch, A.; Jahnen-Dechent, W.; Smith, E.R. Phosphate, Calcification in Blood, and Mineral Stress: The Physiologic Blood Mineral Buffering System and Its Association with Cardiovascular Risk. Int. J. Nephrol. 2018, 2018, 9182078. [Google Scholar] [CrossRef] [PubMed]
- Tintut, Y.; Demer, L.L. Recent Advances in Multifactorial Regulation of Vascular Calcification. Curr. Opin. Lipidol. 2001, 12, 555–560. [Google Scholar] [CrossRef] [PubMed]
- Sage, A.P.; Tintut, Y.; Demer, L.L. Regulatory Mechanisms in Vascular Calcification. Nat. Rev. Cardiol. 2010, 7, 528–536. [Google Scholar] [CrossRef] [PubMed]
- Heiss, A.; Pipich, V.; Jahnen-Dechent, W.; Schwahn, D. Fetuin-a Is a Mineral Carrier Protein: Small Angle Neutron Scattering Provides New Insight on Fetuin-A Controlled Calcification Inhibition. Biophys. J. 2010, 99, 3986–3995. [Google Scholar] [CrossRef]
- Wald, J.; Wiese, S.; Eckert, T.; Jahnen-Dechent, W.; Heiss, A.; Richtering, W. Fetuin-A Mediated Formation and Ripening of Colloidal Calciprotein Particles. Eur. Biophys. J. Biophys. Lett. 2011, 40, 64–65. [Google Scholar]
- Wald, J.; Wiese, S.; Eckert, T.; Jahnen-Dechent, W.; Richtering, W.; Heiss, A. Formation and Stability Kinetics of Calcium Phosphate-Fetuin-a Colloidal Particles Probed by Time-Resolved Dynamic Light Scattering. Soft Matter 2011, 7, 2869–2874. [Google Scholar] [CrossRef]
- Proctor, G.B. The Physiology of Salivary Secretion. Periodontology 2000 2016, 70, 11–25. [Google Scholar] [CrossRef]
- Castagnola, M.; Cabras, T.; Iavarone, F.; Vincenzoni, F.; Vitali, A.; Pisano, E.; Nemolato, S.; Scarano, E.; Fiorita, A.; Vento, G.; et al. Top-Down Platform for Deciphering the Human Salivary Proteome. J. Matern. Fetal Neonatal Med. 2012, 25, 27–43. [Google Scholar] [CrossRef]
- Brandao, E.; Soares, S.; Mateus, N.; de Freitas, V. Human Saliva Protein Profile: Influence of Food Ingestion. Food Res. Int. 2014, 64, 508–513. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.W.; Rhodus, N.L.; Griffin, R.J.; Carlis, J.V.; Griffin, T.J. A Catalogue of Human Saliva Proteins Identified by Free Flow Electrophoresis-Based Peptide Separation and Tandem Mass Spectrometry. Mol. Cell. Proteom. 2005, 4, 1826–1830. [Google Scholar] [CrossRef] [PubMed]
- Amado, F.; Lobo, M.J.C.; Domingues, P.; Alberto Duarte, J.; Vitorino, R. Salivary Peptidomics. Expert Rev. Proteom. 2010, 7, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Grøn, P. State of Calcium and Inorganic Orthophosphate in Human Saliva. Arch. Oral Biol. 1973, 18, 1365–1378. [Google Scholar] [CrossRef]
- Poff, A.M.; Pearce, E.I.F.; Larsen, M.J.; Cutress, T.W. Human Supragingival in Vivo Calculus Formation in Relation to Saturation of Saliva with Respect to Calcium Phosphates. Arch. Oral Biol. 1997, 42, 93–99. [Google Scholar] [CrossRef]
- Jin, Y.; Yip, H.K. Supragingival Calculus: Formation and Control. Crit. Rev. Oral Biol. Med. 2002, 13, 426–441. [Google Scholar] [CrossRef]
- Hay, D.I.; Schlesinger, D.H. Human Salivary Statherin: A Peptide Inhibitor of Calcium Phosphate Precipitation. In Calcium-Binding Proteins and Calcium Functions, Proceedings of the International Symposium on Calcium Binding Proteins and Calcium Functions in Health and Disease, San Diego, CA, USA, 5–9 June 1977; Wassermann, R.H., MacLennan, D.H., Siegel, F., Eds.; Elsevier North Holland: New York, NY, USA, 1977; pp. 401–408. [Google Scholar]
- Reynolds, E.C. Remineralization of Enamel Subsurface Lesions by Casein Phosphopeptide-Stabilized Calcium Phosphate Solutions. J. Dent. Res. 1997, 76, 1587–1595. [Google Scholar] [CrossRef]
- Bautista, D.S.; Denstedt, J.; Chambers, A.F.; Harris, J.F. Low-Molecular-Weight Variants of Osteopontin Generated by Serine Proteinases in Urine of Patients with Kidney Stones. J. Cell. Biochem. 1996, 61, 402–409. [Google Scholar] [CrossRef]
- Wei, M.; Shiraga, H.; Chalko, C.; Goldfarb, S.; Gopal Krishna, G.; Hoyer, J.R. Quantitative Studies of Human Urinary Excretion of Uropontin. Kidney Int. 1998, 53, 189–193. [Google Scholar]
- Schlieper, G.; Westenfeld, R.; Brandenburg, V.; Ketteler, M. Inhibitors of Calcification in Blood and Urine. Semin. Dial. 2007, 20, 113–121. [Google Scholar] [CrossRef]
- Anderson, H. Clarke. Matrix Vesicles and Calcification. Curr. Rheumatol. Rep. 2003, 5, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Landis, W.J.; Jacquet, R. Association of Calcium and Phosphate Ions with Collagen in the Mineralization of Vertebrate Tissues. Calcif. Tissue Int. 2013, 93, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Niu, L.; Jee, S.E.; Jiao, K.; Tonggu, L.; Li, M.; Wang, L.; Yang, Y.; Bian, J.; Breschi, L.; Jang, S.S.; et al. Collagen Intrafibrillar Mineralization as a Result of the Balance between Osmotic Equilibrium and Electroneutrality. Nat. Mater. 2017, 16, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Silver, F.H.; Landis, W.J. Deposition of Apatite in Mineralizing Vertebrate Extracellular Matrices: A Model of Possible Nucleation Sites on Type I Collagen. Connect. Tissue Res. 2011, 52, 242–254. [Google Scholar] [CrossRef]
- Zou, Z.Y.; Tang, T.T.; Macias-Sanchez, E.; Sviben, S.; Landis, W.J.; Bertinetti, L.; Fratzl, P. Three-Dimensional Structural Interrelations between Cells, Extracellular Matrix, and Mineral in Normally Mineralizing Avian Leg Tendon. Proc. Natl. Acad. Sci. USA 2020, 117, 14102–14109. [Google Scholar] [CrossRef]
- Mertz, E.L.; Leikin, S. Interactions of Inorganic Phosphate and Sulfate Anions with Collagen. Biochemistry 2004, 43, 14901–14912. [Google Scholar] [CrossRef]
- Olszta, M.J.; Cheng, X.G.; Jee, S.S.; Kumar, R.; Kim, Y.Y.; Kaufman, M.J.; Douglas, E.P.; Gower, L.B. Bone Structure and Formation: A New Perspective. Mater. Sci. Eng. R Rep. 2007, 58, 77–116. [Google Scholar] [CrossRef]
- Thula, T.T.; Rodriguez, D.E.; Lee, M.H.; Pendi, L.; Podschun, J.; Gower, L.B. In Vitro Mineralization of Dense Collagen Substrates: A Biomimetic Approach toward the Development of Bone-Graft Materials. Acta Biomater. 2011, 7, 3158–3169. [Google Scholar] [CrossRef]
- Jiang, Y.; Gower, L.; Volkmer, D.; Coelfen, H. The Existence Region and Composition of a Polymer-Induced Liquid Precursor Phase for Dl-Glutamic Acid Crystals. Phys. Chem. Chem. Phys. 2012, 14, 914–919. [Google Scholar] [CrossRef]
- Olszta, M.J.; Douglas, E.P.; Gower, L.B. Scanning Electron Microscopic Analysis of the Mineralization of Type I Collagen via a Polymer-Induced Liquid-Precursor (Pilp) Process. Calcif. Tissue Int. 2003, 72, 583–591. [Google Scholar] [CrossRef]
- Thula, T.T.; Svedlund, F.; Rodriguez, D.E.; Podschun, J.; Pendi, L.; Gower, L.B. Mimicking the Nanostructure of Bone: Comparison of Polymeric Process-Directing Agents. Polymers 2011, 3, 10–35. [Google Scholar] [CrossRef] [PubMed]
- Jee, S.-S.; Thula, T.T.; Gower, L.B. Development of Bone-Like Composites Via the Polymer-Induced Liquid-Precursor (Pilp) Process. Part 1: Influence of Polymer Molecular Weight. Acta Biomater. 2010, 6, 3676–3686. [Google Scholar] [CrossRef] [PubMed]
- Jee, S.S.; Kasinath, R.K.; DiMasi, E.; Kim, Y.-Y.; Gower, L. Oriented Hydroxyapatite in Turkey Tendon Mineralized via the Polymer-Induced Liquid-Precursor (Pilp) Process. Crystengcomm 2011, 13, 2077–2083. [Google Scholar] [CrossRef]
- Burwell, A.K.; Thula-Mata, T.; Gower, L.B.; Habeliz, S.; Kurylo, M.; Ho, S.P.; Chien, Y.-C.; Cheng, J.; Cheng, N.F.; Gansky, S.A.; et al. Functional Remineralization of Dentin Lesions Using Polymer-Induced Liquid-Precursor Process. PLoS ONE 2012, 7, e38852. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.R.; Rodriguez, D.E.; Gower, L.B.; Monga, M. Association of Randall Plaque with Collagen Fibers and Membrane Vesicles. J. Urol. 2012, 187, 1094–1100. [Google Scholar] [CrossRef]
- Chidambaram, A.; Rodriguez, D.; Khan, S.; Gower, L. Biomimetic Randall’s Plaque as an in Vitro Model System for Studying the Role of Acidic Biopolymers in Idiopathic Stone Formation. Urolithiasis 2015, 43, S77–S92. [Google Scholar] [CrossRef][Green Version]
- O’Kell, A.L.; Lovett, A.C.; Canales, B.K.; Gower, L.B.; Khan, S.R. Development of a Two-Stage Model System to Investigate the Mineralization Mechanisms Involved in Idiopathic Stone Formation: Stage 2 in Vivo Studies of Stone Growth on Biomimetic Randall’s Plaque. Urolithiasis 2019, 47, 335–346. [Google Scholar] [CrossRef]
- Kuliasha, C.A.; Rodriguez, D.; Lovett, A.; Gower, L.B. In Situ Flow Cell Platform for Examining Calcium Oxalate and Calcium Phosphate Crystallization on Films of Basement Membrane Extract in the Presence of Urinary ‘Inhibitors’. Crystengcomm 2020, 22, 1448–1458. [Google Scholar] [CrossRef]
- Thrivikraman, G.; Athirasala, A.; Gordon, R.; Zhang, L.M.; Bergan, R.; Keene, D.R.; Jones, J.M.; Xie, H.; Chen, Z.Q.; Tao, J.; et al. Rapid Fabrication of Vascularized and Innervated Cell-Laden Bone Models with Biomimetic Intrafibrillar Collagen Mineralization. Nat. Commun. 2019, 10, 3520. [Google Scholar] [CrossRef]
- Lovett, A.C.; Khan, S.R.; Gower, L.B. Development of a Two-Stage in Vitro Model System to Investigate the Mineralization Mechanisms Involved in Idiopathic Stone Formation: Stage 1-Biomimetic Randall’s Plaque Using Decellularized Porcine Kidneys. Urolithiasis 2019, 47, 321–334. [Google Scholar] [CrossRef]
- Akiva, A.; Malkinson, G.; Masic, A.; Kerschnitzki, M.; Bennet, M.; Fratzl, P.; Addadi, L.; Weiner, S.; Yaniv, K. On the Pathway of Mineral Deposition in Larval Zebrafish Caudal Fin Bone. Bone 2015, 75, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Larsson, K. Lyotropic Liquid Crystals and Their Dispersions Relevant in Foods. Curr. Opin. Colloid Interface Sci. 2009, 14, 16–20. [Google Scholar] [CrossRef]
- Kerschnitzki, M.; Akiva, A.; Shoham, A.B.; Koifman, N.; Shimoni, E.; Rechav, K.; Arraf, A.A.; Schultheiss, T.M.; Talmon, Y.; Zelzer, E.; et al. Transport of Membrane-Bound Mineral Particles in Blood Vessels During Chicken Embryonic Bone Development. Bone 2016, 83, 65–72. [Google Scholar] [CrossRef]
- Mahamid, J.; Aichmayer, B.; Shimoni, E.; Ziblat, R.; Li, C.; Siegel, S.; Paris, O.; Fratzl, P.; Weiner, S.; Addadi, L. Mapping Amorphous Calcium Phosphate Transformation into Crystalline Mineral from the Cell to the Bone in Zebrafish Fin Rays. Proc. Natl. Acad. Sci. USA 2010, 107, 6316–6321. [Google Scholar] [CrossRef] [PubMed]
- Mahamid, J.; Sharir, A.; Gur, D.; Zelzer, E.; Addadi, L.; Weiner, S. Bone Mineralization Proceeds through Intracellular Calcium Phosphate Loaded Vesicles: A Cryo-Electron Microscopy Study. J. Struct. Biol. 2011, 174, 527–535. [Google Scholar] [CrossRef]
- Nudelman, F.; Pieterse, K.; George, A.; Bomans, P.H.H.; Friedrich, H.; Brylka, L.J.; Hilbers, P.A.J.; de With, G.; Sommerdijk, N. The Role of Collagen in Bone Apatite Formation in the Presence of Hydroxyapatite Nucleation Inhibitors. Nat. Mater. 2010, 9, 1004–1009. [Google Scholar] [CrossRef]
- Nudelman, F.; Sommerdijk, N. Biomineralization as an Inspiration for Materials Chemistry. Angew. Chem. Int. Ed. 2012, 51, 6582–6596. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Species | UniProt | CaP-SLiM | Protein | Species | UniProt | CaP-SLiM |
|---|---|---|---|---|---|---|---|
| SIBLING SCPPs | Non-SIBLING SCPPS | ||||||
| MEPE | Human | Q9NQ76 | 493-DDSSESSDSGSSSESDGD-510 | CSN1S1 | Guinea | P04656 | 19-SSSSSSSEER-28 |
| DSPP | Human | Q9NZW4 | 661-DSSSSSDSSNSSDSSDS-677 | pig | 54-IISESTEERE-63 | ||
| IBSP | Human | P21815 | 55-GDDSSEEEEE-64 | 65-SSISSSEEV-73 | |||
| OPN | Cow | P31096 | 6-TSSGSSEEKQ-15 | CSN1S2 | Pig | P39036 | 5-EHVSSSEESI-14 |
| 42-QNSVSSEETD-51 | 54-ASSSSSEESVE-63 | ||||||
| 99-SDESHHSDES-108 | 130-ELSTSEEPVS-139 | ||||||
| DMP1 | Mouse | O55188 | 8-NTESESSEER-17 | CSN2 | Human | P05814 | 5-ESLSSSEESI-14 |
| 28-PTNSESSEES-37 | Statherin | Human | P02808 | 1-MIGADSSEE-10 | |||
| 161-DSAQDSESEE-170 | ENAM | Human | Q9NRM1 | 36-SSKSEE-41 | |||
| Non-SCPPs | AMBN | Human | Q9NP70 | 26-SLSLE-30 | |||
| FETUA | Human | P02765 | 307-SLGSPSGEVS-316 | 246-SSEE-249 | |||
| SPP24 | Human | Q13103 | 108-SWSSSTSESYSSEE-117 | AMTN | Human | Q6UX39 | 100-SSEE-103 |
| MGP | Human | P08493 | 2-ESHESMESYE-11 | SCPPPQ1 | Anolis | E0YCE6 | 4-RRSASSSEERRR-15 |
| PRB4 | Human | P10163 | 2-SSSEDVSQEE-11 | ODAM | Human | A1E959 | 25-SASNSNE-31 |
| RBP | Chicken | P02752 | 192-ESSSMSSSEE-201 | FDCSP | Human | Q8NFU4 | 11-KRSISDSDELA-21 |
| Parameter | Experiment | Parameter | Calculation |
|---|---|---|---|
| Empirical volume CaP, nm3 | 0.167 | 0.12 | |
| Surface area per peptide, nm2 | 1.62 | 1.7 | |
| 10−4 Da | 6.0 | 6.0 | |
| 10−5 Da | 1.37 | 1.37 | |
| , nm | 2.44 | 2.44 | |
| Empirical formula CaP | Ca(HPO4)0.4(PO4)0.4 | 3.65 | |
| pKS | 8.0 | 0.940 | |
| 0.785 | |||
| Modal | 436 | ||
| ± SD | 468 ± 147 | ||
| 0.099 |
| Phosphopeptide | Sequence |
|---|---|
| αS1-casein B 59–79 4P | QMEAESISSSEEIVPNSVEQK |
| β-casein A2 1–25 4P | RELEELNVPGEIVESLSSSEESITR |
| CK2-S 4P | MRELEELNVPGADDSSSDDDSDDDDRINKK |
| CK2-SS 4P | MRELEELNVPGADDSSSSDDDDDDDRINKK |
| CK2-S-6H 5P | MRELEELNVPGADDSSSDDDSDDDDRINKKIEDPNSSSVDKLAAALEHHHHHH |
| Biofluid | Cmax, mM | n | pH1/2-pHc | pHc |
|---|---|---|---|---|
| Bovine milk | 1977 ± 3.0 | 1.190 ± 0.003 | 0.508 ± 0.001 | 5.716 ± 0.004 |
| Human milk | 743 ±1.6 | 0.914 ± 0.006 | 1.458 ± 0.583 | 7.200 ± 0.003 |
| Human blood serum | 166 ± 7.6 | 0.949 ± 0.024 | 1.240 ± 0.011 | 7.760 ± 0.002 |
| Human stimulated saliva | 133 ± 3.6 | 1.036 ±0.003 | 0.468 ± 0.003 | 6.880 ± 0.002 |
| Human urine | 398 ± 5.8 | 1.075 ± 0.022 | 0.531 ± 0.015 | 6.529 ± 0.002 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lenton, S.; Wang, Q.; Nylander, T.; Teixeira, S.; Holt, C. Structural Biology of Calcium Phosphate Nanoclusters Sequestered by Phosphoproteins. Crystals 2020, 10, 755. https://doi.org/10.3390/cryst10090755
Lenton S, Wang Q, Nylander T, Teixeira S, Holt C. Structural Biology of Calcium Phosphate Nanoclusters Sequestered by Phosphoproteins. Crystals. 2020; 10(9):755. https://doi.org/10.3390/cryst10090755
Chicago/Turabian StyleLenton, Samuel, Qian Wang, Tommy Nylander, Susana Teixeira, and Carl Holt. 2020. "Structural Biology of Calcium Phosphate Nanoclusters Sequestered by Phosphoproteins" Crystals 10, no. 9: 755. https://doi.org/10.3390/cryst10090755
APA StyleLenton, S., Wang, Q., Nylander, T., Teixeira, S., & Holt, C. (2020). Structural Biology of Calcium Phosphate Nanoclusters Sequestered by Phosphoproteins. Crystals, 10(9), 755. https://doi.org/10.3390/cryst10090755