Screening of a Novel Glycoside Hydrolase Family 51 α-L-Arabinofuranosidase from Paenibacillus polymyxa KF-1: Cloning, Expression, and Characterization

Abstract
1. Introduction
2. Results
2.1. Measurement of the Extracellular Hemicellulytic Activities
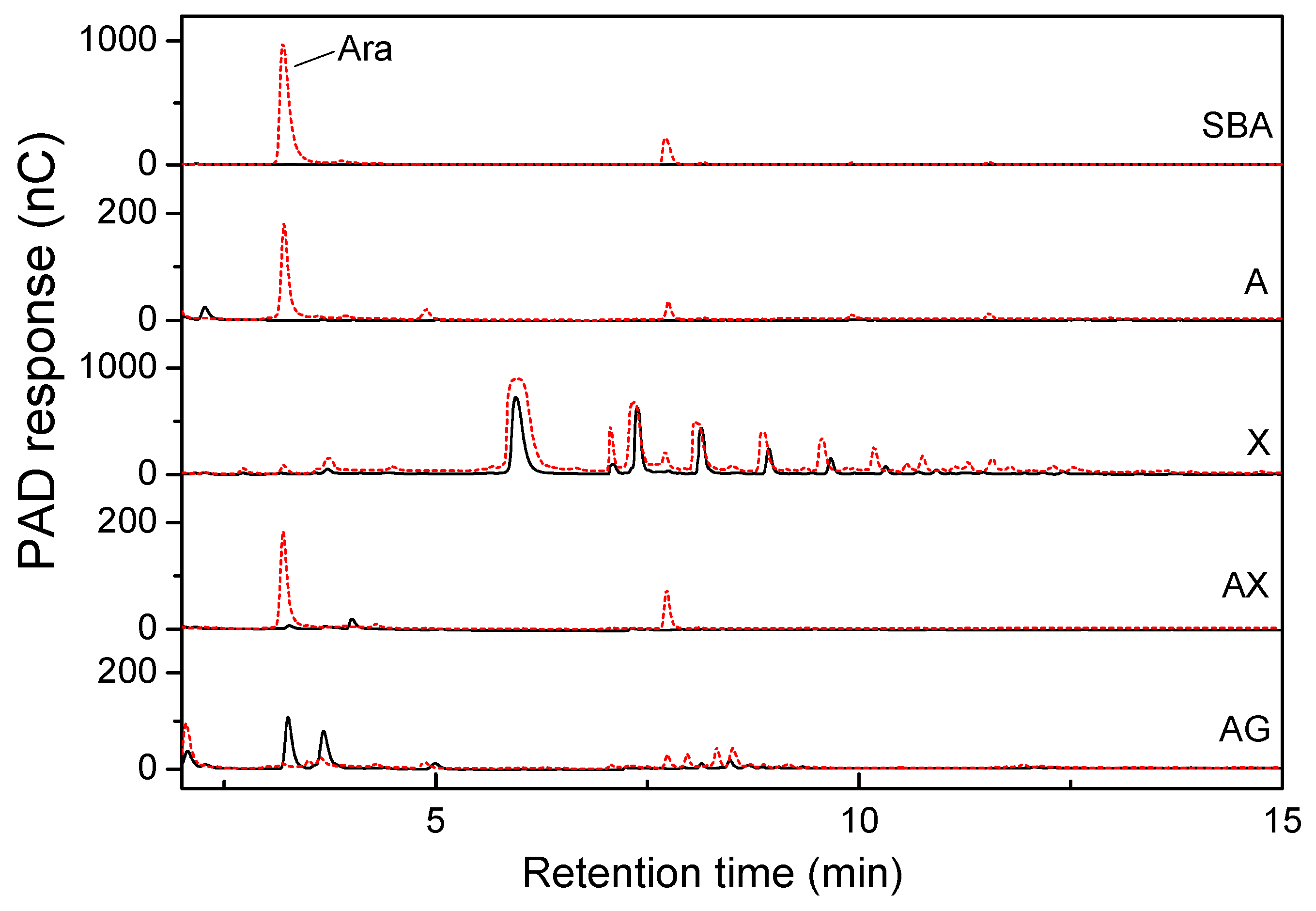
2.2. Hemicellulose-Degrading Enzymes Secreted by P. polymyxa
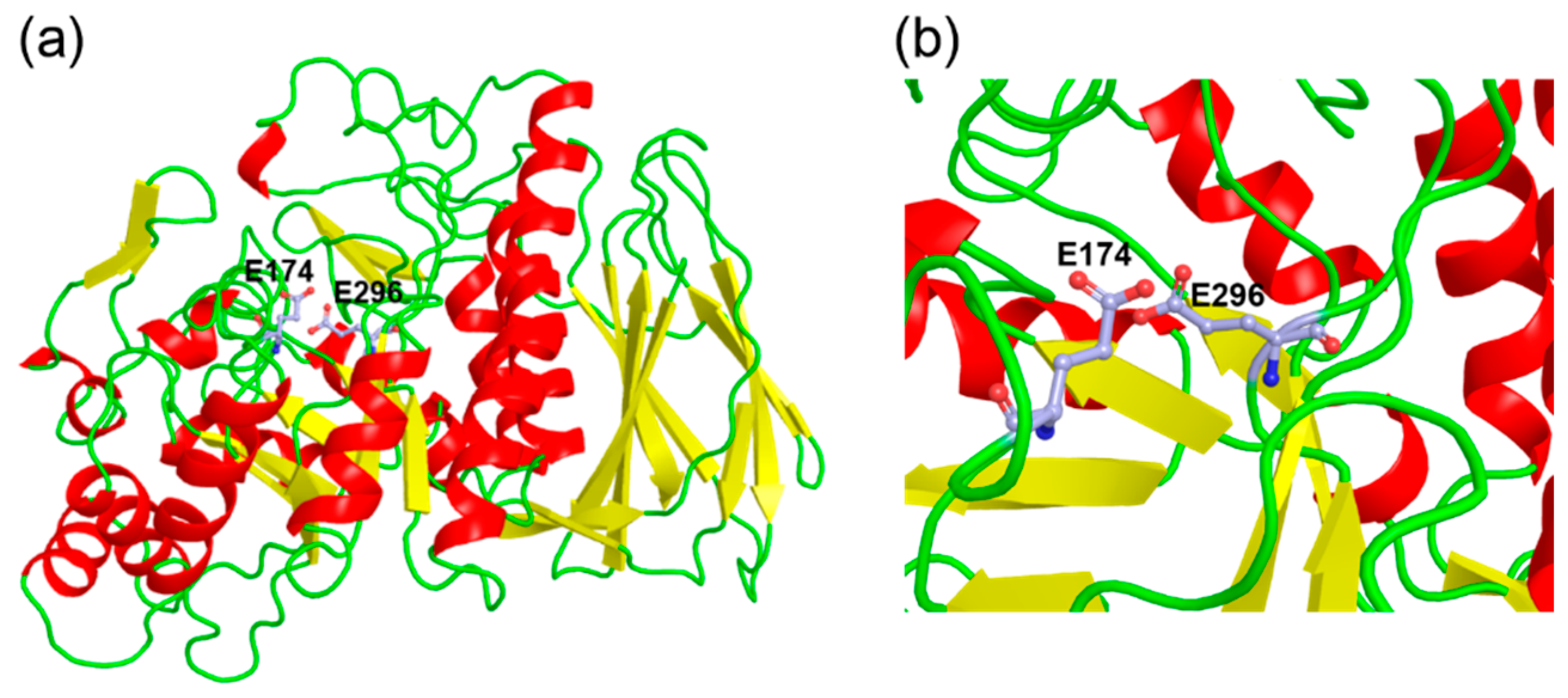
2.3. Cloning and Sequence Analysis of α-L-Arabinofuranosidase
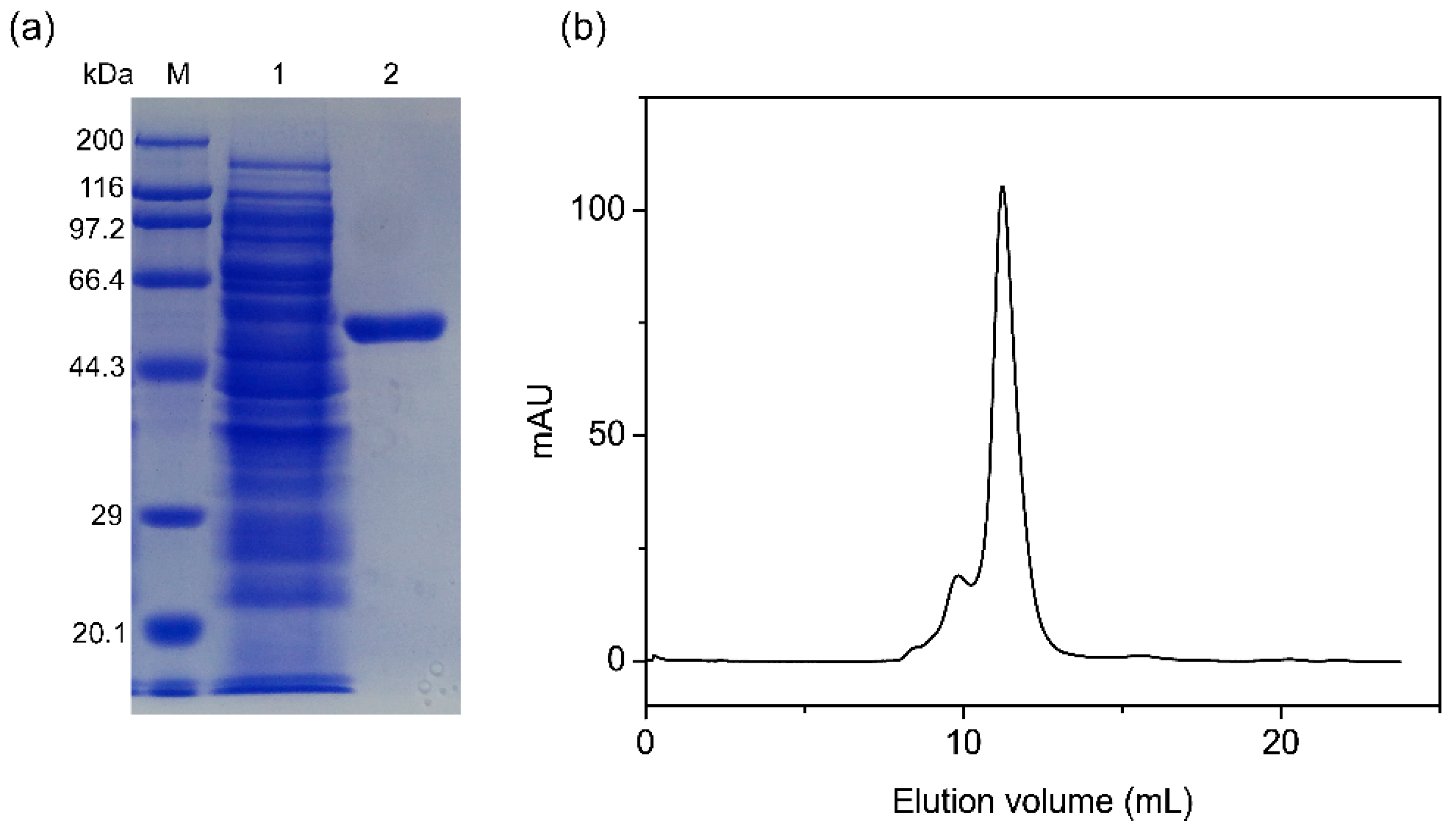
2.4. Expression and Purification of PpAbf51b
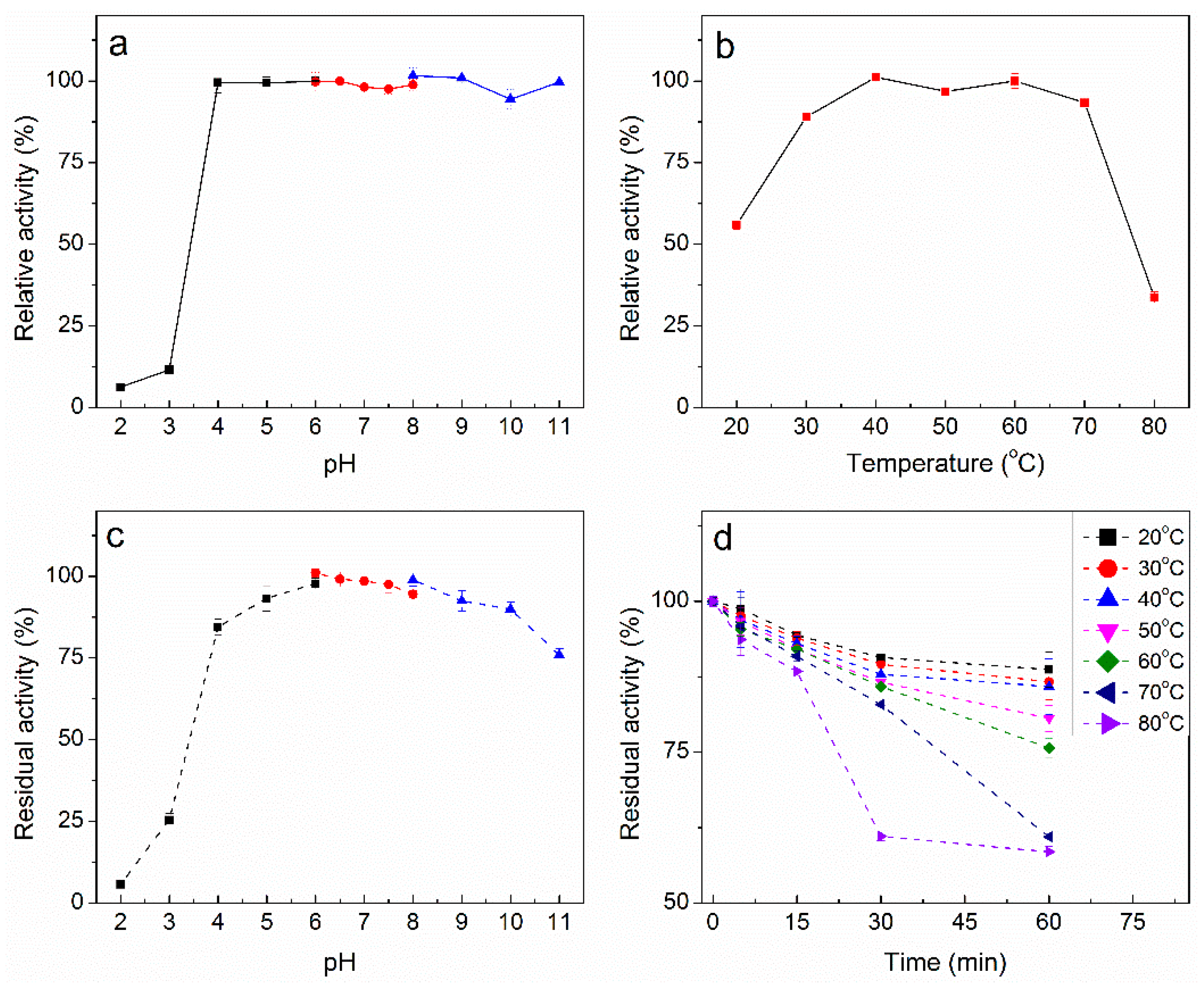
2.5. Characterization of Purified PpAbf51b
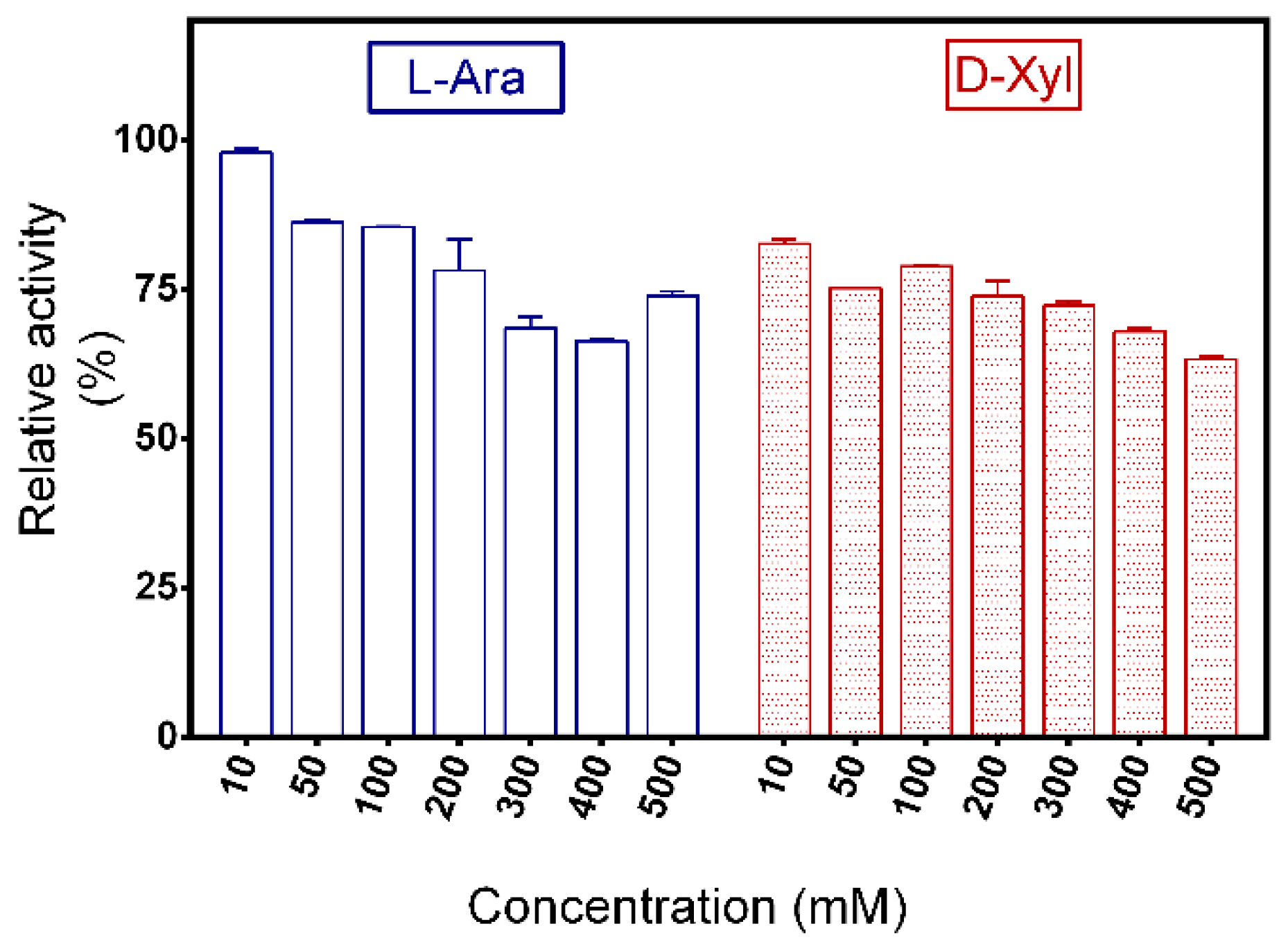
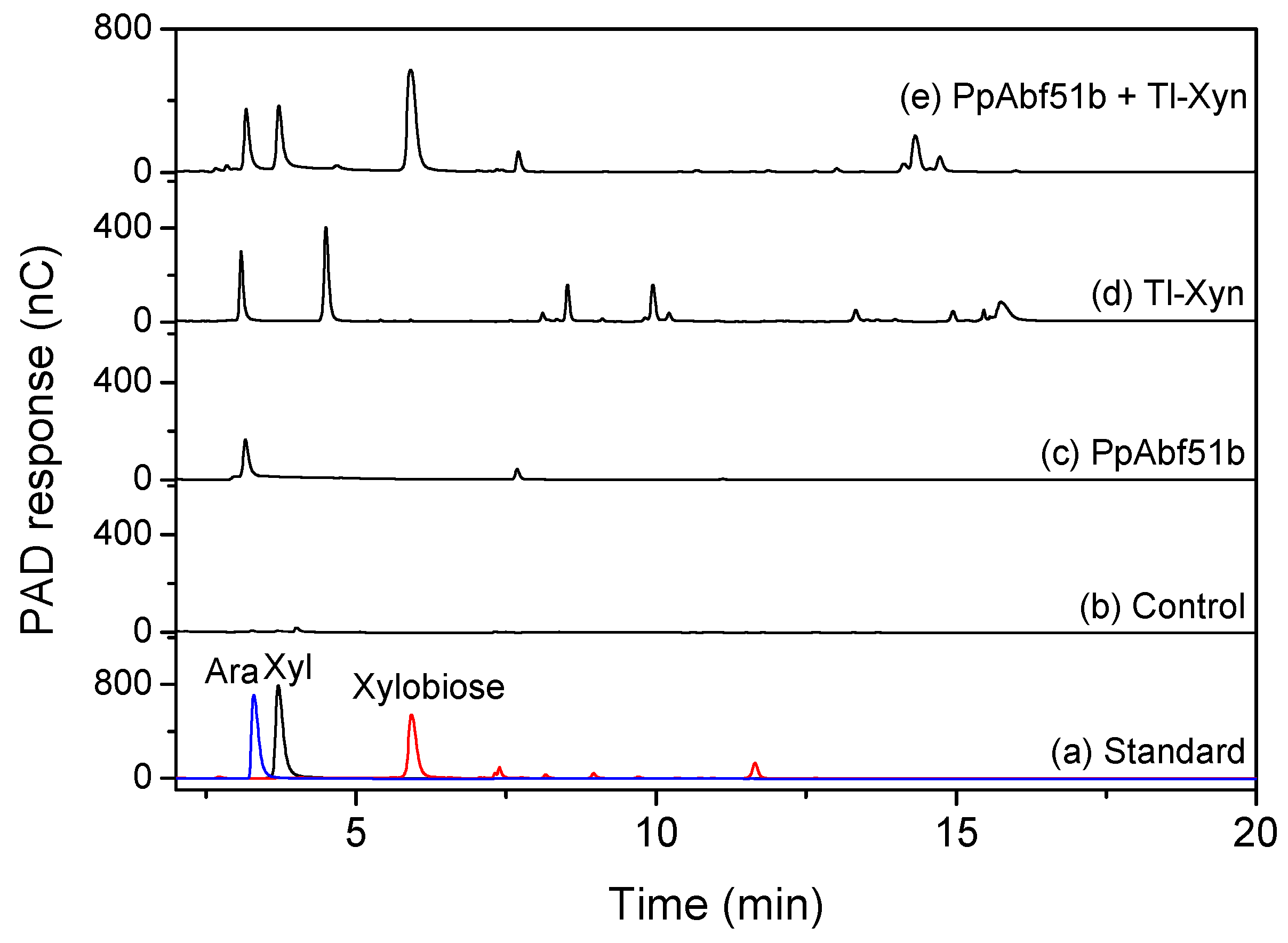
2.6. Substrate Specificity of PpAbf51b
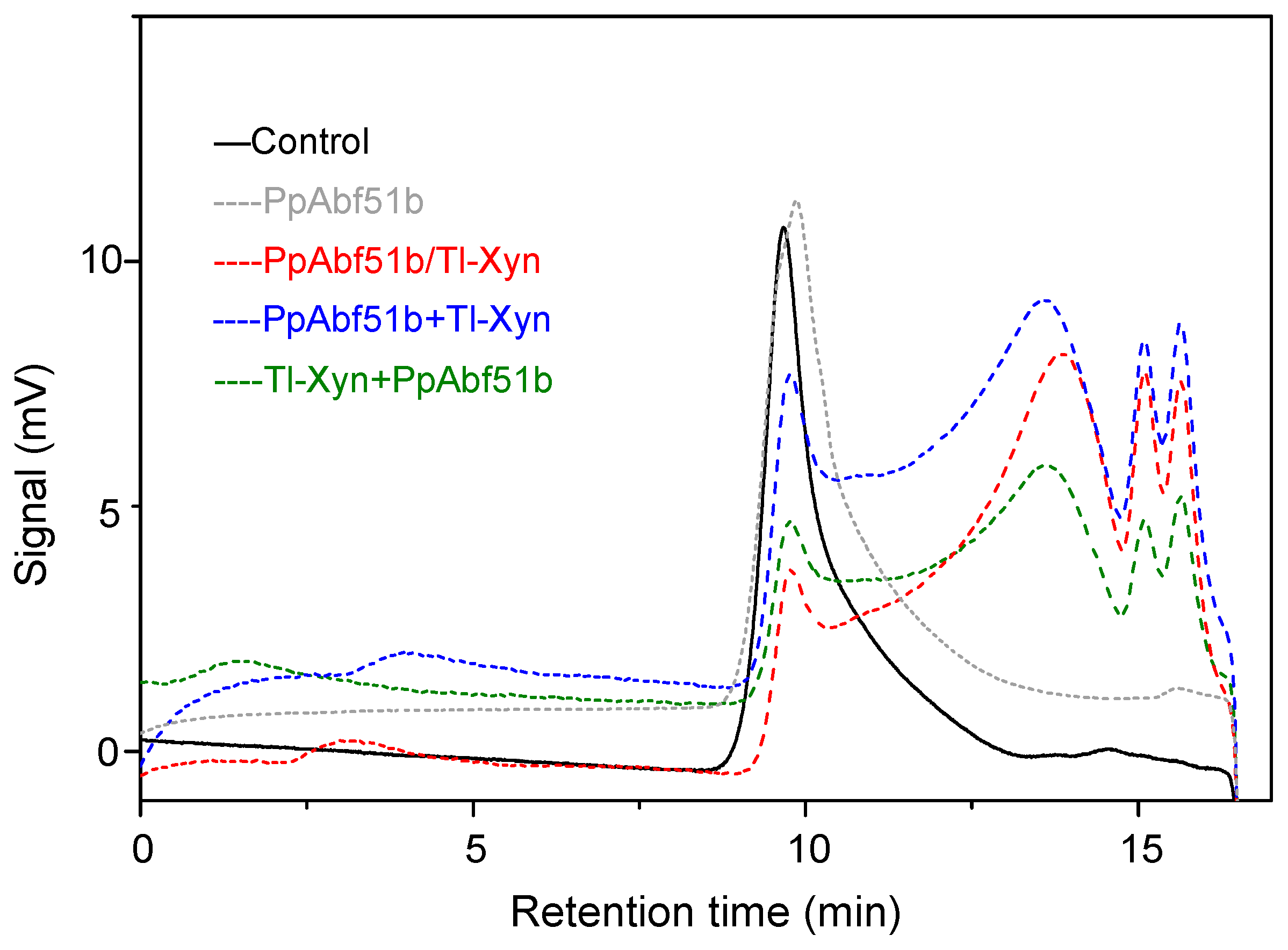
2.7. Synergistic Hydrolysis of Wheat Arabinoxylan by PpAbf51b and Tl-Xyn
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Strains and Culture Conditions
4.3. Hemicellulytic Activities of the P. polymyxa KF-1 Enzyme Extract
4.4. LC-MS/MS Analysis
4.5. Gene Cloning, Protein Expression, and Purification
4.6. Sequence Analysis and Protein Structure Prediction
4.7. Characterization of Recombinant PpAbf51b
4.8. Substrate Specificity of PpAbf51b
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Paulova, L.; Patakova, P.; Branska, B.; Rychtera, M.; Melzoch, K. Lignocellulosic ethanol: Technology design and its impact on process efficiency. Biotechnol. Adv. 2015, 33, 1091–1107. [Google Scholar] [CrossRef] [PubMed]
- Fen, L.; Xuwei, Z.; Nanyi, L.; Puyu, Z.; Shuang, Z.; Xue, Z.; Pengju, L.; Qichao, Z.; Haiping, L. Screening of lignocellulose-degrading superior mushroom strains and determination of their CMCase and laccase activity. Sci. World J. 2014, 2014, 763108. [Google Scholar] [CrossRef] [PubMed]
- Lopes, A.M.; Ferreira Filho, E.X.; Moreira, L.R.S. An update on enzymatic cocktails for lignocellulose breakdown. J. Appl. Microbiol. 2018, 125, 632–645. [Google Scholar] [CrossRef] [PubMed]
- De Gonzalo, G.; Colpa, D.I.; Habib, M.H.M.; Fraaije, M.W. Bacterial enzymes involved in lignin degradation. J. Biotechnol. 2016, 236, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.K.; Arora, D.S. Fungal degradation of lignocellulosic residues: An aspect of improved nutritive quality. Crit. Rev. Microbiol. 2015, 41, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Saha, B.C. Hemicellulose bioconversion. J. Ind. Microbiol. Biotechnol. 2003, 30, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Goldbeck, R.; Damasio, A.R.L.; Goncalves, T.A.; Machado, C.B.; Paixao, D.A.A.; Wolf, L.D.; Mandelli, F.; Rocha, G.J.M.; Ruller, R.; Squina, F.M. Development of hemicellulolytic enzyme mixtures for plant biomass deconstruction on target biotechnological applications. Appl. Microbiol. Biotechnol. 2014, 98, 8513–8525. [Google Scholar] [CrossRef] [PubMed]
- Wilkens, C.; Andersen, S.; Dumon, C.; Berrin, J.-G.; Svensson, B. GH62 arabinofuranosidases: Structure, function and applications. Biotechnol. Adv. 2017, 35, 792–804. [Google Scholar] [CrossRef] [PubMed]
- Seri, K.; Sanai, K.; Matsuo, N.; Kawakubo, K.; Xue, C.; Inoue, S. L-arabinose selectively inhibits intestinal sucrase in an uncompetitive manner and suppresses glycemic response after sucrose ingestion in animals. Metabolism 1996, 45, 1368–1374. [Google Scholar] [CrossRef]
- Lagaert, S.; Pollet, A.; Courtin, C.M.; Volckaert, G. beta-xylosidases and alpha-L-arabinofuranosidases: Accessory enzymes for arabinoxylan degradation. Biotechnol. Adv. 2014, 32, 316–332. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, Q.; Li, Y.; Gao, J.; Fan, X. Draft Genome Sequence of Paenibacillus polymyxa KF-1, an Excellent Producer of Microbicides. Genome Announc. 2016, 4. [Google Scholar] [CrossRef] [PubMed]
- Naghmouchi, K.; Baah, J.; Cudennec, B.; Drider, D. Required characteristics of Paenibacillus polymyxa JB-0501 as potential probiotic. Arch. Microbiol. 2013, 195, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Zhao, Y.; Zhang, G.; Li, Y.; Li, Q. Production optimization, purification, expression, and characterization of a novel α-l-arabinofuranosidase from Paenibacillus polymyxa. Electron. J. Biotechnol. 2018, 36, 24–33. [Google Scholar] [CrossRef]
- Rybakova, D.; Wetzlinger, U.; Muller, H.; Berg, G. Complete Genome Sequence of Paenibacillus polymyxa Strain Sb3-1, a Soilborne Bacterium with Antagonistic Activity toward Plant Pathogens. Genome Announc. 2015, 3. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Bendtsen, J.D.; Kiemer, L.; Fausboll, A.; Brunak, S. Non-classical protein secretion in bacteria. BMC Microbiol. 2005, 5, 58. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, M.R.; Gasteiger, E.; Bairoch, A.; Sanchez, J.C.; Williams, K.L.; Appel, R.D.; Hochstrasser, D.F. Protein identification and analysis tools in the ExPASy server. Methods Mol. Biol. 1999, 112, 531–552. [Google Scholar] [PubMed]
- Tyanova, S.; Temu, T.; Cox, J. The MaxQuant computational platform for mass spectrometry-based shotgun proteomics. Nat. Protoc. 2016, 11, 2301–2319. [Google Scholar] [CrossRef] [PubMed]
- McManus, F.P.; Lamoliatte, F.; Thibault, P. Identification of cross talk between SUMOylation and ubiquitylation using a sequential peptide immunopurification approach. Nat. Protoc. 2017, 12, 2342–2358. [Google Scholar] [CrossRef] [PubMed]
- Paes, G.; Skov, L.K.; O’Donohue, M.J.; Remond, C.; Kastrup, J.S.; Gajhede, M.; Mirza, O. The structure of the complex between a branched pentasaccharide and Thermobacillus xylanilyticus GH-51 arabinofuranosidase reveals xylan-binding determinants and induced fit. Biochemistry 2008, 47, 7441–7451. [Google Scholar] [CrossRef] [PubMed]
- Dumbrepatil, A.; Park, J.-M.; Jung, T.Y.; Song, H.-N.; Jang, M.-U.; Han, N.S.; Kim, T.-J.; Woo, E.J. Structural analysis of alpha-L-arabinofuranosidase from Thermotoga maritima reveals characteristics for thermostability and substrate specificity. J. Microbiol. Biotechnol. 2012, 22, 1724–1730. [Google Scholar] [CrossRef] [PubMed]
- Souza, T.A.C.B.; Santos, C.R.; Souza, A.R.; Oldiges, D.P.; Ruller, R.; Prade, R.A.; Squina, F.M.; Murakami, M.T. Structure of a novel thermostable GH51 alpha-L-arabinofuranosidase from Thermotoga petrophila RKU-1. Protein Sci. 2011, 20, 1632–1637. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Bai, Y.; Yang, P.; Luo, H.; Huang, H.; Meng, K.; Shi, P.; Wang, Y.; Yao, B. A novel bifunctional GH51 exo-alpha-l-arabinofuranosidase/endo-xylanase from Alicyclobacillus sp. A4 with significant biomass-degrading capacity. Biotechnol. Biofuels 2015, 8, 197. [Google Scholar] [CrossRef] [PubMed]
- Bouraoui, H.; Desrousseaux, M.-L.; Ioannou, E.; Alvira, P.; Manai, M.; Remond, C.; Dumon, C.; Fernandez-Fuentes, N.; O’Donohue, M.J. The GH51 alpha-l-arabinofuranosidase from Paenibacillus sp. THS1 is multifunctional, hydrolyzing main-chain and side-chain glycosidic bonds in heteroxylans. Biotechnol. Biofuels 2016, 9, 140. [Google Scholar] [CrossRef] [PubMed]
- Shallom, D.; Belakhov, V.; Solomon, D.; Shoham, G.; Baasov, T.; Shoham, Y. Detailed kinetic analysis and identification of the nucleophile in alpha-L-arabinofuranosidase from Geobacillus stearothermophilus T-6, a family 51 glycoside hydrolase. J. Biol. Chem. 2002, 277, 43667–43673. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Higgins, D.G. Clustal omega. Curr. Protoc. Bioinform. 2014, 48, 3–13. [Google Scholar] [CrossRef]
- Morales, P.; Sendra, J.M.; Perez-Gonzalez, J.A. Purification and characterization of an arabinofuranosidase from Bacillus polymyxa expressed in Bacillus subtilis. Appl. Microbiol. Biotechnol. 1995, 44, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Im, D.-H.; Kimura, K.; Hayasaka, F.; Tanaka, T.; Noguchi, M.; Kobayashi, A.; Shoda, S.; Miyazaki, K.; Wakagi, T.; Fushinobu, S. Crystal structures of glycoside hydrolase family 51 alpha-L-arabinofuranosidase from Thermotoga maritima. Biosci. Biotechnol. Biochem. 2012, 76, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Canakci, S.; Belduz, A.O.; Saha, B.C.; Yasar, A.; Ayaz, F.A.; Yayli, N. Purification and characterization of a highly thermostable alpha-L-Arabinofuranosidase from Geobacillus caldoxylolyticus TK4. Appl. Microbiol. Biotechnol. 2007, 75, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Canakci, S.; Kacagan, M.; Inan, K.; Belduz, A.O.; Saha, B.C. Cloning, purification, and characterization of a thermostable alpha-L-arabinofuranosidase from Anoxybacillus kestanbolensis AC26Sari. Appl. Microbiol. Biotechnol. 2008, 81, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Margolles, A.; de los Reyes-Gavilan, C.G. Purification and functional characterization of a novel alpha-L-arabinofuranosidase from Bifidobacterium longum B667. Appl. Environ. Microbiol. 2003, 69, 5096–5103. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Lee, Y.-E. Cloning, expression, and characterization of a thermostable GH51 alpha-L-arabinofuranosidase from Paenibacillus sp. DG-22. J. Microbiol. Biotechnol. 2014, 24, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Debeche, T.; Cummings, N.; Connerton, I.; Debeire, P.; O’Donohue, M.J. Genetic and biochemical characterization of a highly thermostable alpha-L-arabinofuranosidase from Thermobacillus xylanilyticus. Appl. Environ. Microbiol. 2000, 66, 1734–1736. [Google Scholar] [CrossRef] [PubMed]
- De Giuseppe, P.O.; Souza, T.D.A.; Souza, F.H.M.; Zanphorlin, L.M.; Machado, C.B.; Ward, R.J.; Jorge, J.A.; Furriel, R.P.M.; Murakami, M.T. Structural basis for glucose tolerance in GH1 beta-glucosidases. Acta Crystallogr. D Biol. Crystallogr. 2014, 70, 1631–1639. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, W.H.; Bronnenmeier, K.; Krause, B.; Lottspeich, F.; Staudenbauer, W.L. Debranching of arabinoxylan: Properties of the thermoactive recombinant alpha-L-arabinofuranosidase from Clostridium stercorarium (ArfB). Appl. Microbiol. Biotechnol. 1995, 43, 856–860. [Google Scholar] [CrossRef] [PubMed]
- Guilloux, K.; Gaillard, I.; Courtois, J.; Courtois, B.; Petit, E. Production of arabinoxylan-oligosaccharides from flaxseed (Linum usitatissimum). J. Agric. Food Chem. 2009, 57, 11308–11313. [Google Scholar] [CrossRef] [PubMed]
- Shi, P.; Chen, X.; Meng, K.; Huang, H.; Bai, Y.; Luo, H.; Yang, P.; Yao, B. Distinct actions by Paenibacillus sp. strain E18 alpha-L-arabinofuranosidases and xylanase in xylan degradation. Appl. Environ. Microbiol. 2013, 79, 1990–1995. [Google Scholar] [CrossRef] [PubMed]
- McCleary, B.V.; McGeough, P. A Comparison of Polysaccharide Substrates and Reducing Sugar Methods for the Measurement of endo-1,4-beta-Xylanase. Appl. Biochem. Biotechnol. 2015, 177, 1152–1163. [Google Scholar] [CrossRef] [PubMed]
- Talamantes, T.; Ughy, B.; Domonkos, I.; Kis, M.; Gombos, Z.; Prokai, L. Label-free LC-MS/MS identification of phosphatidylglycerol-regulated proteins in Synechocystis sp. PCC6803. Proteomics 2014, 14, 1053–1057. [Google Scholar] [CrossRef] [PubMed]
- Tyanova, S.; Temu, T.; Carlson, A.; Sinitcyn, P.; Mann, M.; Cox, J. Visualization of LC-MS/MS proteomics data in MaxQuant. Proteomics 2015, 15, 1453–1456. [Google Scholar] [CrossRef] [PubMed]
- Sonnett, M.; Gupta, M.; Nguyen, T.; Wuhr, M. Quantitative Proteomics for Xenopus Embryos II, Data Analysis. Methods Mol. Biol. 2018, 1865, 195–215. [Google Scholar] [CrossRef] [PubMed]
- Lessard, J.C. Molecular cloning. Methods Enzymol. 2013, 529, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.M. The bicinchoninic acid (BCA) assay for protein quantitation. Methods Mol. Biol. 1994, 32, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Brunelle, J.L.; Green, R. One-dimensional SDS-polyacrylamide gel electrophoresis (1D SDS-PAGE). Methods Enzymol. 2014, 541, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Biasini, M.; Bienert, S.; Waterhouse, A.; Arnold, K.; Studer, G.; Schmidt, T.; Kiefer, F.; Gallo Cassarino, T.; Bertoni, M.; Bordoli, L.; et al. SWISS-MODEL: Modelling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Res. 2014, 42, W252–W258. [Google Scholar] [CrossRef] [PubMed]
- Ghim, Y.S.; Chang, H.N. Diffusional falsification of kinetic constants on Lineweaver-Burk plots. J. Theor. Biol. 1983, 105, 91–102. [Google Scholar] [CrossRef]
- Wu, D.; Cui, L.; Yang, G.; Ning, X.; Sun, L.; Zhou, Y. Preparing rhamnogalacturonan II domains from seven plant pectins using Penicillium oxalicum degradation and their structural comparison. Carbohydr. Polym. 2018, 180, 209–215. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measured Activity | Substrate | Enzyme Activity (U/mg) 1 |
|---|---|---|
| β-Xylosidase | pNPβXyl | 1.45 ± 0.13 |
| α-L-arabinofuranosidase | pNPαAraf | 12.66 ± 0.34 |
| β-Mannosidase | pNPβMan | 1.22 ± 0.11 |
| α-Galactosidase | pNPαGal | 1.98 ± 0.16 |
| α-Glucuronidase | pNPαGlcA | - 2 |
| β-1, 4-Xylanase | oat spelt xylan | 2.68 ± 0.28 |
| β-1, 4-Mannanase | locust bean gum | 0.21 ± 0.02 |
| Entry Name (Uniprot) | Accession No. | Protein Description 1 | Signal Peptide 2 | Non-Classical Signal 3 | Family | Predicted Mw (kDa)/pI 4 | Protein Group Score 5 | Unique Peptides 6 |
|---|---|---|---|---|---|---|---|---|
| A0A378Y5A3 | WP_025676671.1 | 1,4-β-xylanase | 1–32 | GH10 | 78.516/5.24 | 209 | 1 | |
| E3EBI0 | WP_013373220.1 | endo-1,4-β-xylanase | 1–28 | GH11 | 23.203/9.40 | 180 | 2 | |
| E3EB21 | WP_013373834.1 | β-xylanase | 1–30 | GH10 | 35.288/8.98 | 178 | 1 | |
| B1A0K7 | ABZ80847.1 | endo-(1,4)-β-xylanase | 1–26 | GH11 | 68.175/6.09 | 46 | 1 | |
| E3E7K8 | WP_013371416.1 | xyloglucanase | 1–31 | - | 108.95/5.49 | 34 | 1 | |
| E3EGZ1 | WP_013370178.1 | β-xylosidase | 1–32 | CBM13-GH43 | 53.813/5.67 | 150 | 6 | |
| E3E7G9 | WP_016324892.1 | β-xylosidase | 1–32 | GH43 | 56.311/6.32 | 132 | 3 | |
| E3EJD5 | WP_013371027.1 | α-galactosidase | NO | NO | GH27 | 48.518/5.17 | 79 | 1 |
| E3EAP2 | WP_043886149.1 | β-mannosidase | 1–39 | CBM3-CBM35-GH26-GH44 | 149/5.49 | 119 | 3 | |
| E0RKL8 | WP_013310261.1 | arabinoxylan arabinofuranohydrolase | 1–26 | CBM36-CBM6-GH43 | 67.508/5.95 | 148 | 1 | |
| E3ECR1 | WP_013368995.1 | α-N-arabinofuranosidase | NO | NO | GH51 | 56.868/5.57 | 105.9 | 3 |
| E3E519 | WP_014599997.1 | α-N-arabinofuranosidase | NO | NO | GH51 | 56.337/5.62 | 54.3 | 1 |
| V5LER1 | WP_058828950.1 | arabinan endo-1,5-α-L-arabinosidase | 1–35 | GH43 | 35.569/9.17 | 34 | 1 |
| Metal ions or Chemicals (10 mM) | Relative Activity (%) 1 |
|---|---|
| NaCl | 103.2 ± 3.4 |
| KCl | 102.4 ± 1.2 |
| CaCl2 | 91.2 ± 2.8 |
| MgCl2 | 92.5 ± 2.6 |
| FeCl2 | 130.5 ± 2.4 |
| MnCl2 | 82.8 ± 1.5 |
| CuCl2 | 110.5 ± 0.9 |
| EDTA | 91.9 ± 0.9 |
| ZnCl2 | 80.3 ± 2.6 |
| AlCl3 | 72.4 ± 1.8 |
| CoCl2 | 90.8 ± 0.3 |
| FeCl3 | 77.4 ± 3.2 |
| DTT | 95.2 ± 0.4 |
| SDS | 100.2 ± 1.7 |
| Tween-40 | 80.3 ± 1.2 |
| Tween-60 | 92.4 ± 3.0 |
| Tween-80 | 90.0 ± 0.8 |
| TritonX-100 | 111.0 ± 3.2 |
| Substrate | Relative Activity (%) 1 |
|---|---|
| pNPαAraf | 100 ± 0.5 |
| pNPαArap | - 2 |
| pNPβXyl | - |
| pNPαGal | 1.48 ± 0.78 |
| pNPβGal | - |
| pNPαGlc | - |
| pNPβGlc | - |
| pNPαRha | - |
| pNPαMan | - |
| pNPβMan | - |
| Enzyme Added | L-Ara (mg/g) 1 | D-Xyl (mg/g) | Xylobiose (mg/g) | Degree of Synergy 2 | |
|---|---|---|---|---|---|
| First Reaction | Second Reaction | ||||
| PpAbf51b | - 3 | 29.4 ± 2.7 | - | - | - |
| Tl-Xyn | - | - | 45.5 ± 3.0 | 117.9 ± 3.0 | - |
| PpAbf51b | Tl-Xyn | 49.5 ± 1.1 | 55.7 ± 0.1 | 169.8 ± 1.3 | 1.43 |
| Tl-Xyn | PpAbf51b | 46.0 ± 2.15 | 69.9 ± 1.2 | 206.0 ± 2.4 | 1.67 |
| PpAbf51b+Tl-Xyn | - | 45.6 ± 0.3 | 50.0 ± 5.1 | 169.9 ± 1.6 | 1.38 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Y.; Zhao, Y.; Tian, S.; Zhang, G.; Li, Y.; Li, Q.; Gao, J. Screening of a Novel Glycoside Hydrolase Family 51 α-L-Arabinofuranosidase from Paenibacillus polymyxa KF-1: Cloning, Expression, and Characterization. Catalysts 2018, 8, 589. https://doi.org/10.3390/catal8120589
Hu Y, Zhao Y, Tian S, Zhang G, Li Y, Li Q, Gao J. Screening of a Novel Glycoside Hydrolase Family 51 α-L-Arabinofuranosidase from Paenibacillus polymyxa KF-1: Cloning, Expression, and Characterization. Catalysts. 2018; 8(12):589. https://doi.org/10.3390/catal8120589
Chicago/Turabian StyleHu, Yanbo, Yan Zhao, Shuang Tian, Guocai Zhang, Yumei Li, Qiang Li, and Juan Gao. 2018. "Screening of a Novel Glycoside Hydrolase Family 51 α-L-Arabinofuranosidase from Paenibacillus polymyxa KF-1: Cloning, Expression, and Characterization" Catalysts 8, no. 12: 589. https://doi.org/10.3390/catal8120589
APA StyleHu, Y., Zhao, Y., Tian, S., Zhang, G., Li, Y., Li, Q., & Gao, J. (2018). Screening of a Novel Glycoside Hydrolase Family 51 α-L-Arabinofuranosidase from Paenibacillus polymyxa KF-1: Cloning, Expression, and Characterization. Catalysts, 8(12), 589. https://doi.org/10.3390/catal8120589